Water Availability Coincides with Population Declines for an Endangered Butterfly


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
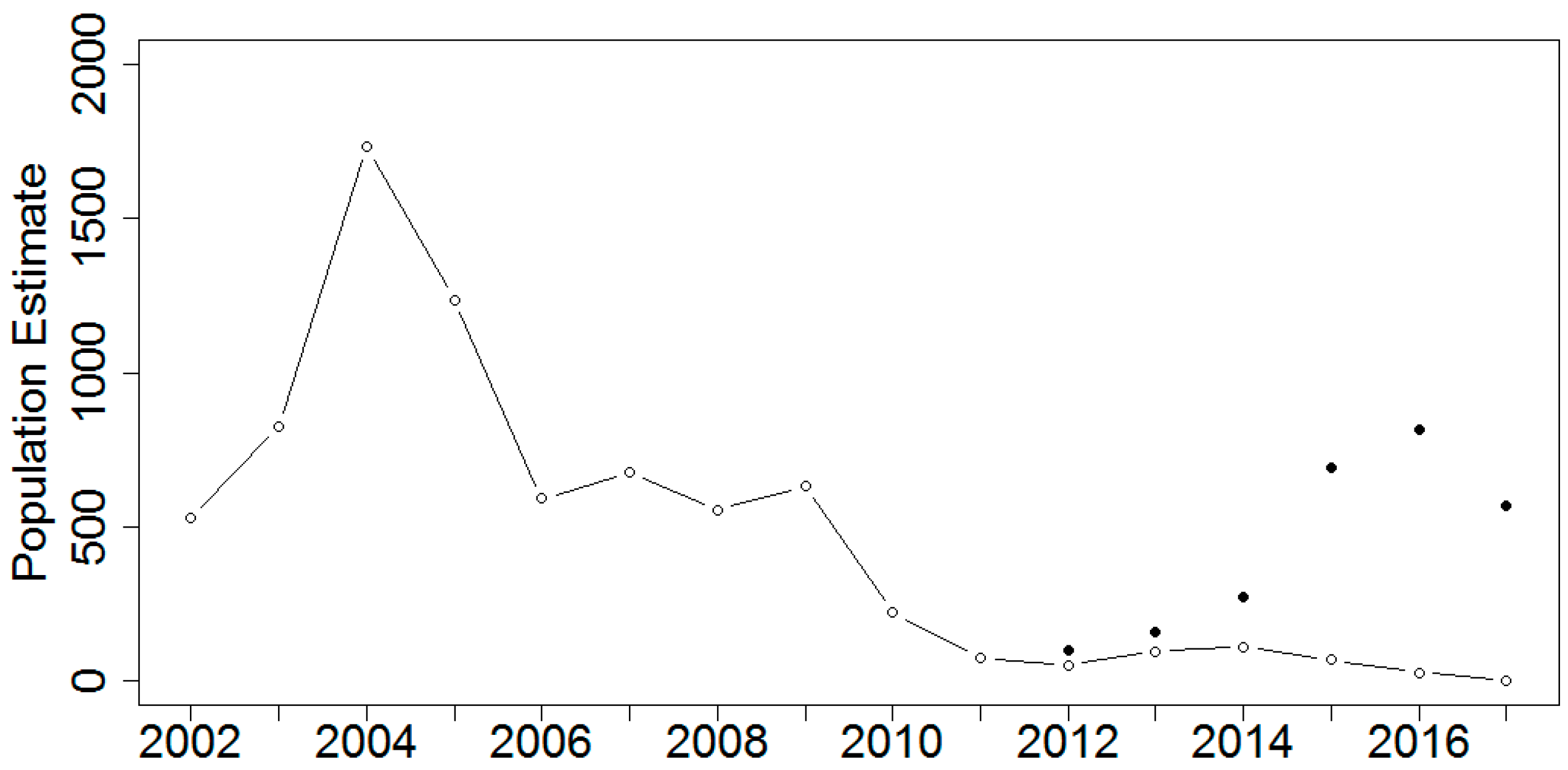
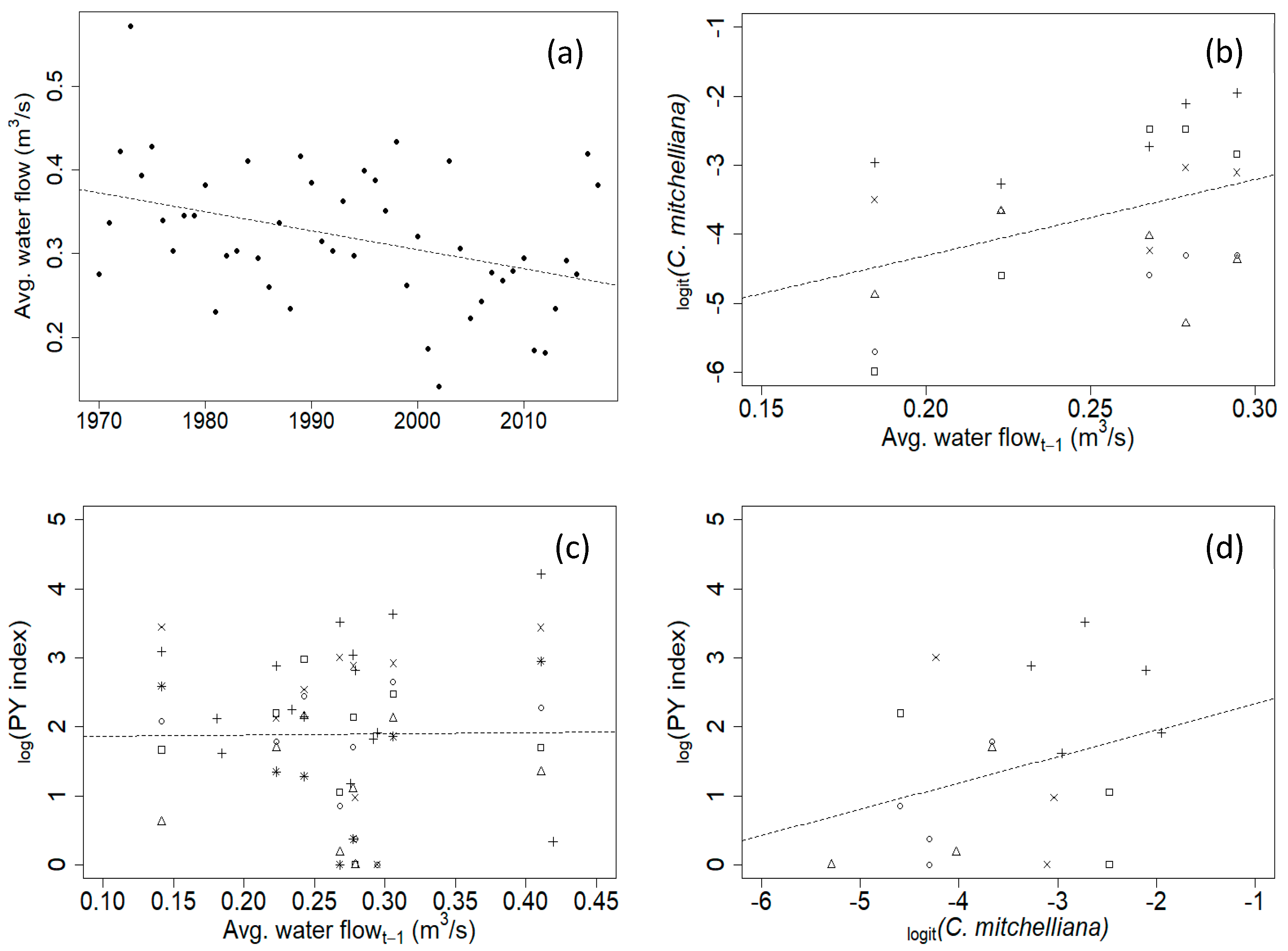
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.-B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; Meester, L.D.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef] [PubMed]
- Scheffers, B.R.; Meester, L.D.; Bridge, T.C.L.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.M.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The broad footprint of climate change from genes to biomes to people. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Schuetz, J.G.; Langham, G.M.; Soykan, C.U.; Wilsey, C.B.; Auer, T.; Sanchez, C.C. Making spatial prioritizations robust to climate change uncertainties: A case study with North American birds. Ecol. Appl. 2015, 25, 1819–1831. [Google Scholar] [CrossRef] [PubMed]
- Ockendon, N.; Baker, D.J.; Carr, J.A.; White, E.C.; Almond, R.E.A.; Amano, T.; Bertram, E.; Bradbury, R.B.; Bradley, C.; Butchart, S.H.M.; et al. Mechanisms underpinning climatic impacts on natural populations: Altered species interactions are more important than direct effects. Glob. Chang. Biol. 2014, 20, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Traill, L.W.; Lim, M.L.M.; Sodhi, N.S.; Bradshaw, C.J.A. Mechanisms driving change: Altered species interactions and ecosystem function through global warming. J. Anim. Ecol. 2010, 79, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Cahill, A.E.; Aiello-Lammens, M.E.; Fisher-Reid, M.C.; Hua, X.; Karanewsky, C.J.; Ryu, H.Y.; Sbeglia, G.C.; Spagnolo, F.; Waldron, J.B.; Warsi, O.; et al. How does climate change cause extinction? Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef] [PubMed]
- Barnett, T.P.; Pierce, D.W.; Hidalgo, H.G.; Bonfils, C.; Santer, B.D.; Das, T.; Bala, G.; Wood, A.W.; Nozawa, T.; Mirin, A.A.; et al. Human-Induced Changes in the Hydrology of the Western United States. Science 2008, 319, 1080–1083. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W.; Donahue, W.F. An impending water crisis in Canada’s western prairie provinces. Proc. Natl. Acad. Sci. USA 2006, 103, 7210–7216. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.B.; Murphy, D.D.; White, R.R. Sun, Slope, and Butterflies: Topographic Determinants of Habitat Quality for Euphydryas Editha. Ecology 1988, 69, 1486–1496. [Google Scholar] [CrossRef]
- Henry, E.H.; Haddad, N.M.; Wilson, J.; Hughes, P.; Gardner, B. Point-count methods to monitor butterfly populations when traditional methods fail: A case study with Miami blue butterfly. J. Insect Conserv. 2015, 19, 519–529. [Google Scholar] [CrossRef]
- Haddeland, I.; Heinke, J.; Biemans, H.; Eisner, S.; Flörke, M.; Hanasaki, N.; Konzmann, M.; Ludwig, F.; Masaki, Y.; Schewe, J.; et al. Global water resources affected by human interventions and climate change. Proc. Natl. Acad. Sci. USA 2014, 111, 3251–3256. [Google Scholar] [CrossRef] [PubMed]
- Nagy, R.C.; Porder, S.; Neill, C.; Brando, P.; Quintino, R.M.; do Nascimento, S.A. Structure and composition of altered riparian forests in an agricultural Amazonian landscape. Ecol. Appl. 2015, 25, 1725–1738. [Google Scholar] [CrossRef] [PubMed]
- Greet, J.; Cousens, R.D.; Webb, J.A. Flow regulation affects temporal patterns of riverine plant seed dispersal: Potential implications for plant recruitment. Freshw. Biol. 2012, 57, 2568–2579. [Google Scholar] [CrossRef]
- Johnson, S.N.; Staley, J.T.; McLeod, F.A.L.; Hartley, S.E. Plant-mediated effects of soil invertebrates and summer drought on above-ground multitrophic interactions. J. Ecol. 2011, 99, 57–65. [Google Scholar] [CrossRef]
- Jamieson, M.A.; Trowbridge, A.M.; Raffa, K.F.; Lindroth, R.L. Consequences of Climate Warming and Altered Precipitation Patterns for Plant-Insect and Multitrophic Interactions. Plant. Physiol. 2012, 160, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Diamond, S.E.; Cayton, H.; Wepprich, T.; Jenkins, C.N.; Dunn, R.R.; Haddad, N.M.; Ries, L. Unexpected phenological responses of butterflies to the interaction of urbanization and geographic temperature. Ecology 2014, 95, 2613–2621. [Google Scholar] [CrossRef]
- Oliver, T.H.; Marshall, H.H.; Morecroft, M.D.; Brereton, T.; Prudhomme, C.; Huntingford, C. Interacting effects of climate change and habitat fragmentation on drought-sensitive butterflies. Nat. Clim. Chang. 2015, 5, 941–945. [Google Scholar] [CrossRef]
- Fox, R. The decline of moths in Great Britain: A review of possible causes. Insect Conserv. Divers. 2013, 6, 5–19. [Google Scholar] [CrossRef]
- McLaughlin, J.F.; Hellmann, J.J.; Boggs, C.L.; Ehrlich, P.R. Climate change hastens population extinctions. Proc. Natl. Acad. Sci. USA 2002, 99, 6070–6074. [Google Scholar] [CrossRef] [PubMed]
- Wallisdevries, M.F.; Van Swaay, C.A.M. Global warming and excess nitrogen may induce butterfly decline by microclimatic cooling. Glob. Chang. Biol. 2006, 12, 1620–1626. [Google Scholar] [CrossRef]
- Ehrlich, P.R.; Murphy, D.D.; Singer, M.C.; Sherwood, C.B.; White, R.R.; Brown, I.L. Extinction, reduction, stability and increase: The responses of checkerspot butterfly (Euphydryas) populations to the California drought. Oecologia 1980, 46, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; O’Connor, R.J. Predicting Extinctions as a Result of Climate Change. Ecology 2006, 87, 1611–1615. [Google Scholar] [CrossRef]
- Severns, P.M.; Boldt, L.; Villegas, S. Conserving a wetland butterfly: Quantifying early lifestage survival through seasonal flooding, adult nectar, and habitat preference. J. Insect Conserv. 2006, 10, 361. [Google Scholar] [CrossRef]
- Joy, J.; Pullin, A.S. The effects of flooding on the survival and behaviour of overwintering large heath butterfly Coenonympha tullia larvae. Biol. Conserv. 1997, 82, 61–66. [Google Scholar] [CrossRef]
- Webb, M.R.; Pullin, A.S. Effects of submergence by winter floods on diapausing caterpillars of a wetland butterfly, Lycaena dispar batavus. Ecol. Entomol. 1998, 23, 96–99. [Google Scholar] [CrossRef]
- Fies, R.; Rabl, D.; Schulze, C.H.; Fiedler, K. Summer floods shape meadow butterfly communities in a floodplain nature reserve in Central Europe. J. Insect Conserv. 2016, 20, 433–445. [Google Scholar] [CrossRef]
- Drahovzal, S.A.; Loftin, C.S.; Rhymer, J. Environmental predictors of shrubby cinquefoil (Dasiphora fruticose) habitat and quality as host for Maine’s endangered Clayton’s copper butterfly (Lycaena dorcas claytoni). Wetl. Ecol. Manag. 2015, 23, 891–908. [Google Scholar] [CrossRef]
- Kuefler, D.; Haddad, N.M.; Hall, S.; Hudgens, B.; Bartel, B.; Hoffman, E. Distribution, population structure and habitat use of the endangered Saint Francis Satyr butterfly, Neonympha mitchellii francisci. Am. Midl. Nat. 2008, 159, 298–320. [Google Scholar] [CrossRef]
- Cayton, H.; Henry, E.; Kiekebusch, E.; Haddad, N. Research for the Conservation and Restoration of an Endangered Butterfly, the St. Francis’ Satyr; Report Submitted to Fort Bragg Army Installation; Fort Bragg Army Installation: Ft. Bragg, NC, USA, 2016. [Google Scholar]
- Cayton, H.; Haddad, N.M.; Ball, B.; Henry, E.; Aschehoug, E. Habitat Restoration as a Recovery Tool for a Disturbance-Dependent Butterfly, The Endangered St. Francis’ Satyr. In Butterfly Conservation in North America; Daniels, J.C., Ed.; Springer: Heidelberg, Germany, 2015; pp. 147–159. ISBN 978-94-017-9851-8. [Google Scholar]
- Cayton, H.L.; Haddad, N.M.; Gross, K.; Diamond, S.E.; Ries, L. Do growing degree days predict phenology across butterfly species? Ecology 2015, 96, 1473–1479. [Google Scholar] [CrossRef]
- Pollard, E. A method for assessing changes in the abundance of butterflies. Biol. Conserv. 1977, 12, 115–134. [Google Scholar] [CrossRef]
- Haddad, N.M.; Hudgens, B.; Damiani, C.; Gross, K.; Kuefler, D.; Pollock, K. Determining Optimal Population Monitoring for Rare Butterflies. Conserv. Biol. 2008, 22, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Pollard, E.; Yates, T.J. Monitoring Butterflies for Ecology and Conservation; Springer: Heidelberg, Germany, 1994; ISBN 978-0-412-63460-4. [Google Scholar]
- Bartel, R.A.; Haddad, N.M.; Wright, J.P. Ecosystem engineers maintain a rare species of butterfly and increase plant diversity. Oikos 2010, 119, 883–890. [Google Scholar] [CrossRef]
- Warton, D.I.; Hui, F.K.C. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhao, X.; Liang, S.; Zhou, T.; Huang, K.; Tang, B.; Zhao, W. Time-lag effects of global vegetation responses to climate change. Glob. Chang. Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef] [PubMed]
- Aschehoug, E.T.; Sivakoff, F.S.; Cayton, H.L.; Morris, W.F.; Haddad, N.M. Habitat restoration affects immature stages of a wetland butterfly through indirect effects on predation. Ecology 2015, 96, 1761–1767. [Google Scholar] [CrossRef] [PubMed]
- Kiesecker, J.M.; Blaustein, A.R.; Belden, L.K. Complex causes of amphibian population declines. Nature 2001, 410, 681–684. [Google Scholar] [CrossRef] [PubMed]
- McMenamin, S.K.; Hadly, E.A.; Wright, C.K. Climatic change and wetland desiccation cause amphibian decline in Yellowstone National Park. Proc. Natl. Acad. Sci. USA 2008, 105, 16988–16993. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, F.J.; Donald, P.F.; Pain, D.J.; Burfield, I.J.; van Bommel, F.P.J. Long-term population declines in Afro-Palearctic migrant birds. Biol. Conserv. 2006, 131, 93–105. [Google Scholar] [CrossRef]
- Quesnelle, P.E.; Fahrig, L.; Lindsay, K.E. Effects of habitat loss, habitat configuration and matrix composition on declining wetland species. Biol. Conserv. 2013, 160, 200–208. [Google Scholar] [CrossRef]
- Ogutu, J.O.; Owen-Smith, N. ENSO, rainfall and temperature influences on extreme population declines among African savanna ungulates. Ecol. Lett. 2003, 6, 412–419. [Google Scholar] [CrossRef]
- Adams, R.A.; Hayes, M.A. Water availability and successful lactation by bats as related to climate change in arid regions of western North America. J. Anim. Ecol. 2008, 77, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Brook, B.W.; Sodhi, N.S.; Bradshaw, C.J.A. Synergies among extinction drivers under global change. Trends Ecol. Evol. 2008, 23, 453–460. [Google Scholar] [CrossRef] [PubMed]
- United States Census Bureau. 2017. Available online: https://www.census.gov/en.html (accessed on 19 July 2016).
- USGS National Water Information System. 2017. Available online: https://waterdata.usgs.gov/nwis (accessed on 22 July 2016).
- NOAA National Centers for Environmental Information (NCEI). 2017. Available online: https://www.ncdc.noaa.gov/ (accessed on 30 April 2018).
- Stefanescu, C.; Carnicer, J.; Peñuelas, J. Determinants of species richness in generalist and specialist Mediterranean butterflies: The negative synergistic forces of climate and habitat change. Ecography 2011, 34, 353–363. [Google Scholar] [CrossRef]
- IPCC. IPCC Working Group I Contribtuion to the the IPCC Fifth Assessment Report—Climate Change 2013: The Physical Science Basis; IPCC: Geneva, Switzerland, 2013; p. 207. [Google Scholar]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cayton, H.L.; Haddad, N.M. Water Availability Coincides with Population Declines for an Endangered Butterfly. Diversity 2018, 10, 94. https://doi.org/10.3390/d10030094
Cayton HL, Haddad NM. Water Availability Coincides with Population Declines for an Endangered Butterfly. Diversity. 2018; 10(3):94. https://doi.org/10.3390/d10030094
Chicago/Turabian StyleCayton, Heather L., and Nick M. Haddad. 2018. "Water Availability Coincides with Population Declines for an Endangered Butterfly" Diversity 10, no. 3: 94. https://doi.org/10.3390/d10030094
APA StyleCayton, H. L., & Haddad, N. M. (2018). Water Availability Coincides with Population Declines for an Endangered Butterfly. Diversity, 10(3), 94. https://doi.org/10.3390/d10030094