Alterations of Arbuscular Mycorrhizal Fungal Diversity in Soil with Elevation in Tropical Forests of China


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Collection
2.2. Analyses of Soil Parameters
2.3. DNA Extraction and PCR Amplification
2.4. Processing of Sequencing Data
2.5. Data Analysis
3. Results
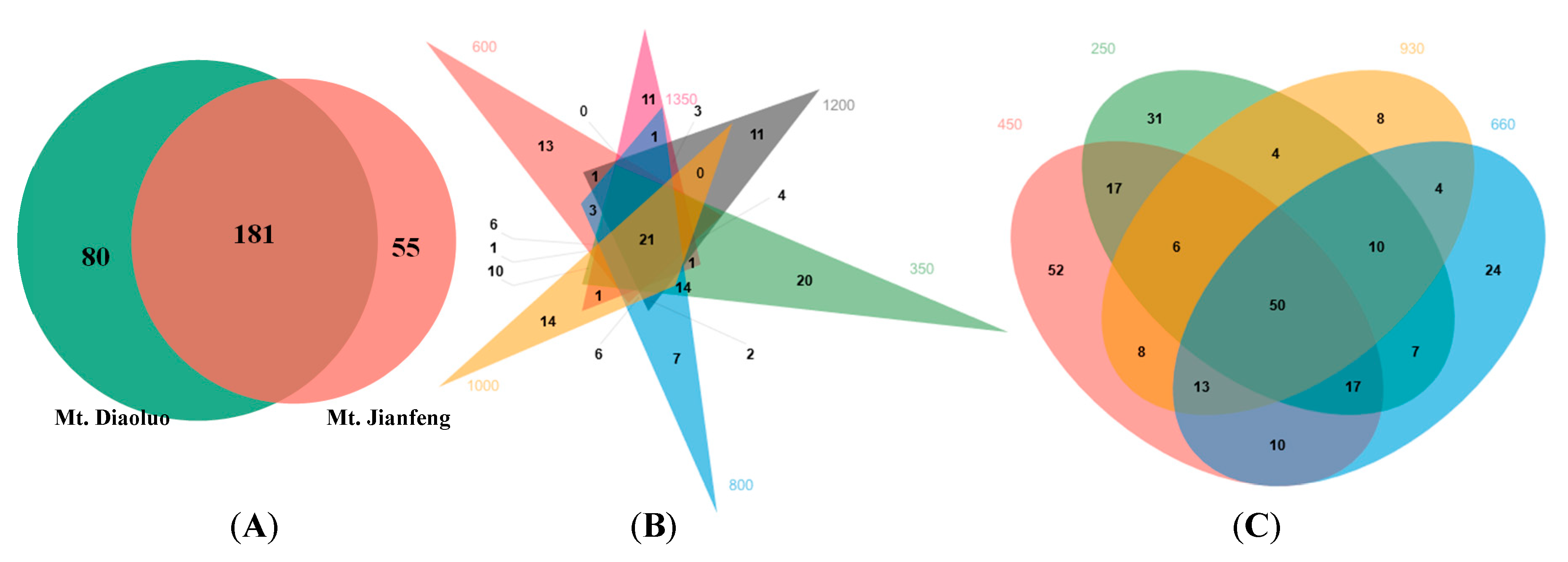
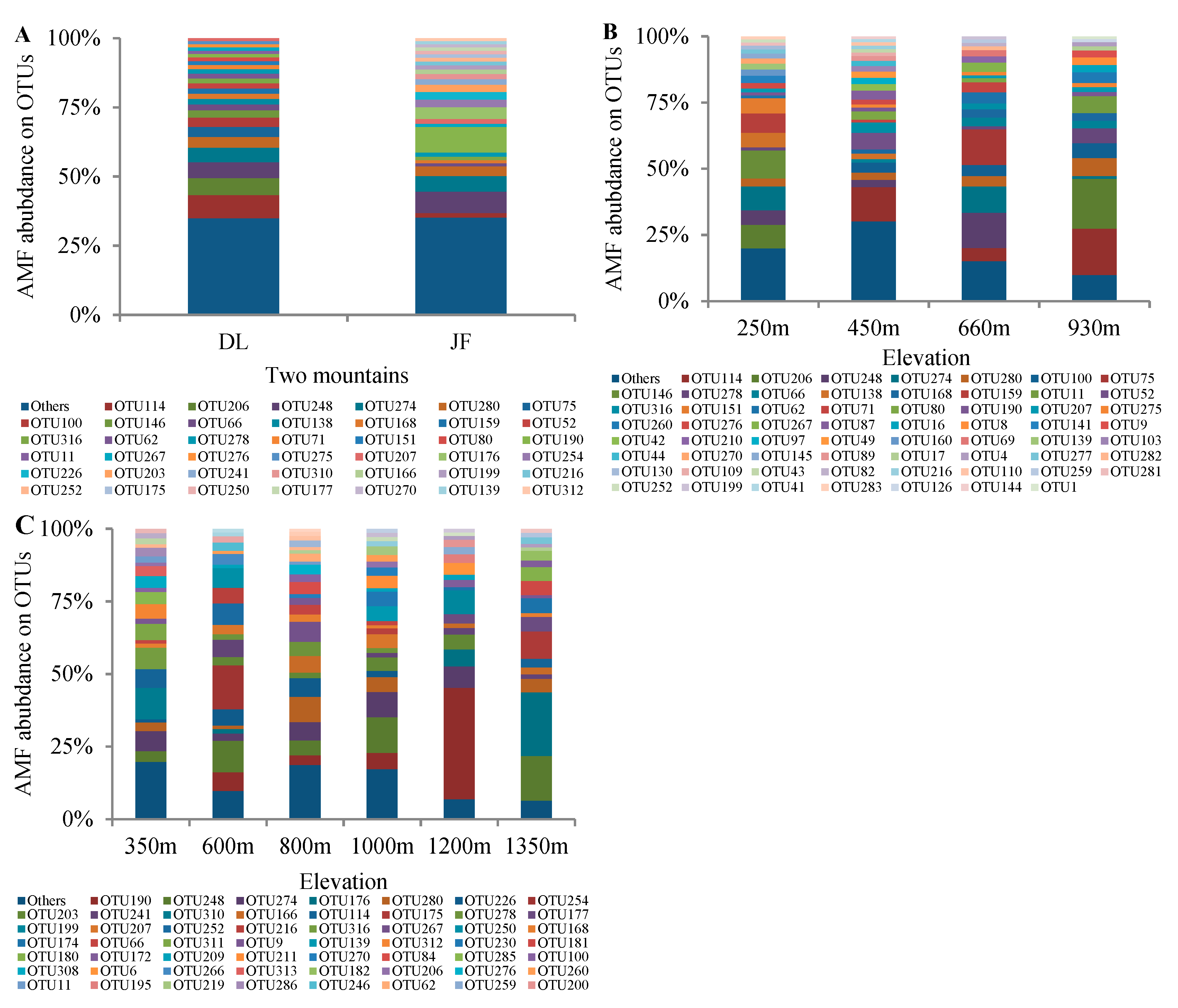
3.1. AMF Community Composition and Distribution in Tropical Forest of China
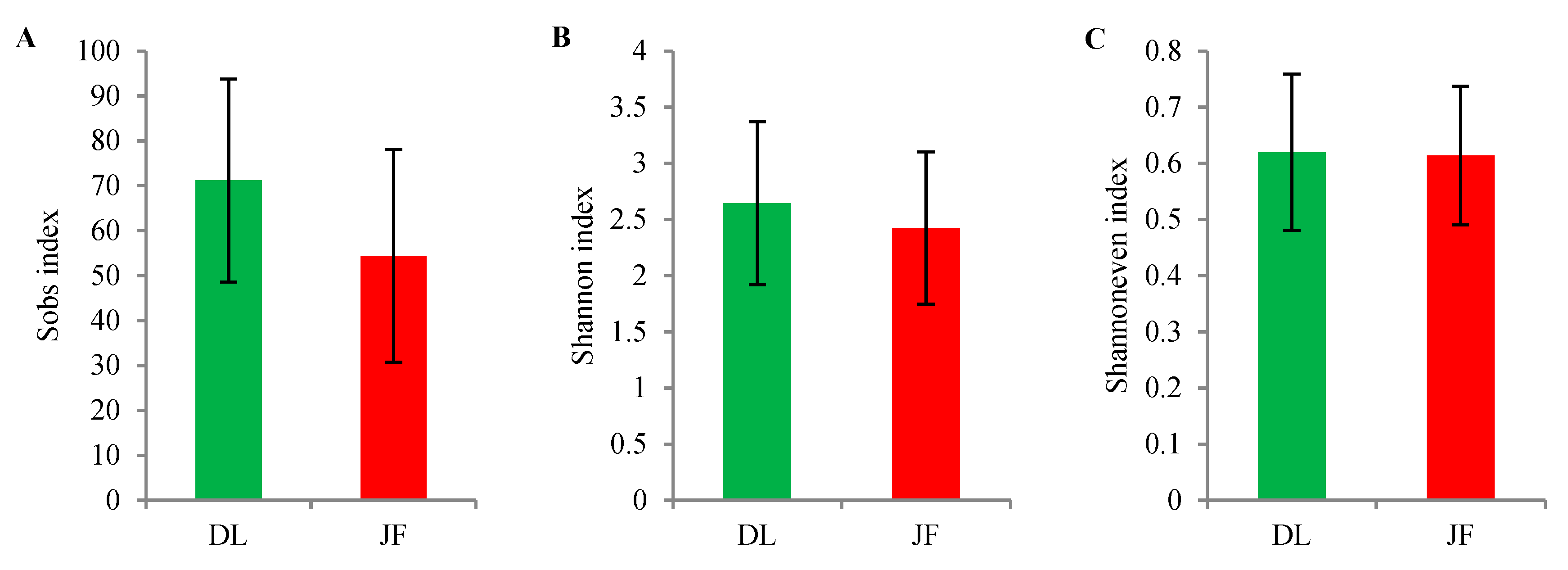
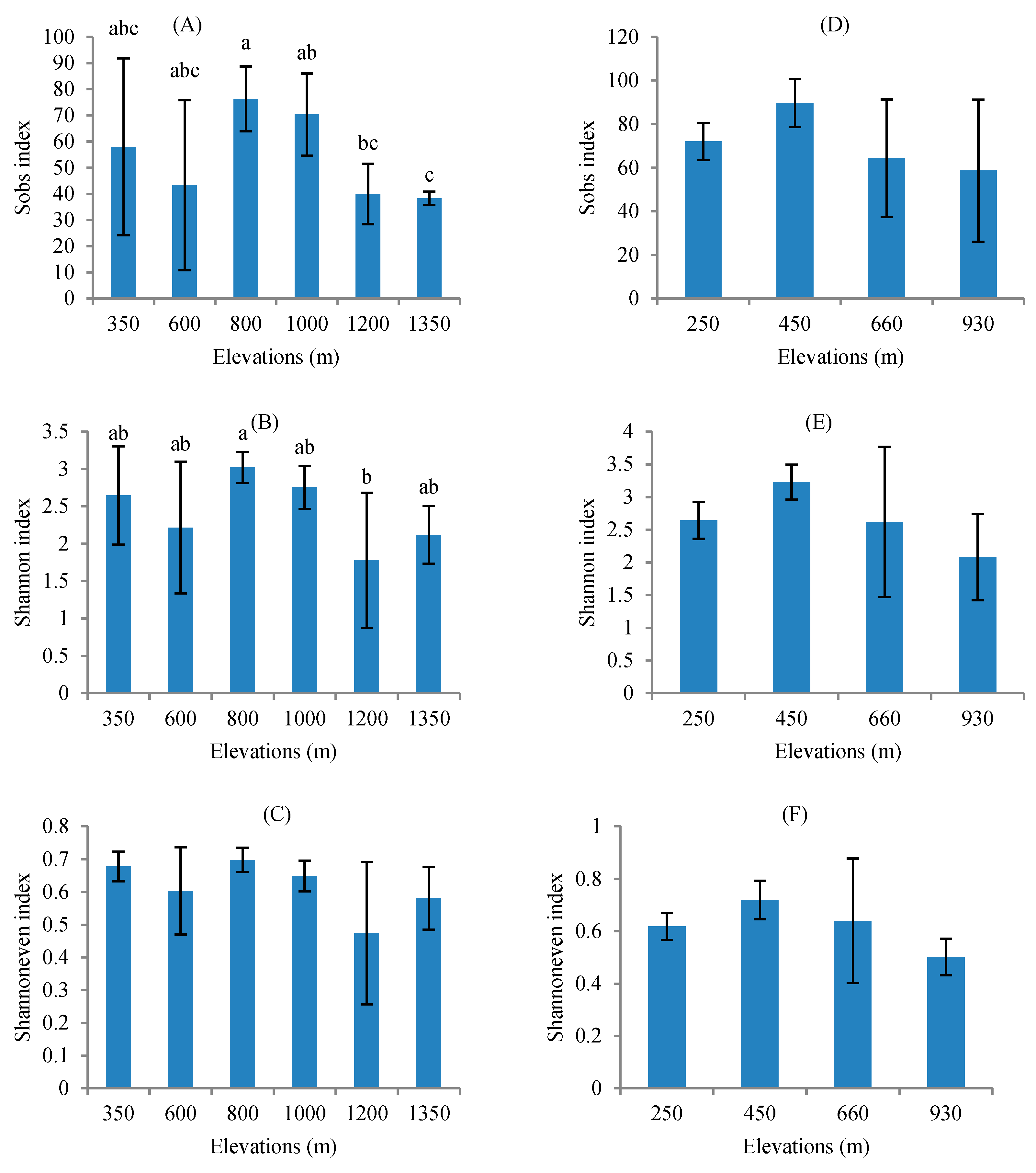
3.2. AMF Diversity between Different Mountains and among Different Elevations in Tropical Forest of China
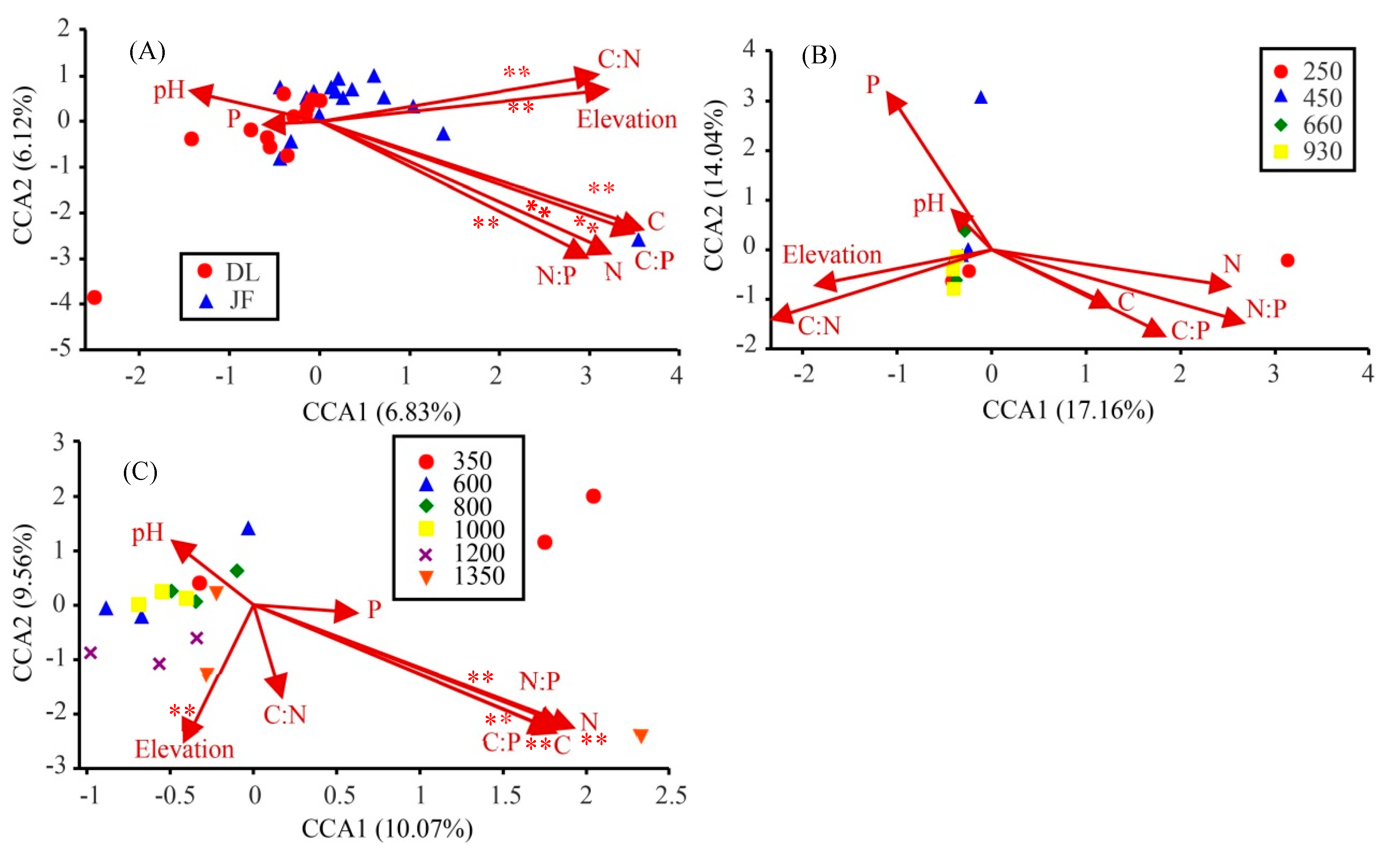
3.3. Contribution of Environmental Factors to AMF Diversity in Tropical Forest of China
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, UK, 2008; pp. 13–89. [Google Scholar]
- Shi, Z.; Zhang, X.; Xu, S.; Lan, Z.; Li, K. Mycorrhizal relationship in lupines: A review. Legume Res. 2017, 40, 965–973. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Jing, X.; Sanders, N.J.; Shi, Y.; Chu, H.; Classen, A.T.; Zhao, K.; Chen, L.; Shi, Y.; Jiang, Y.; He, J.S. The links between ecosystem multifunctionality and above-and belowground biodiversity are mediated by climate. Nat. Commun. 2015, 6, 8159. [Google Scholar] [CrossRef]
- Bachelot, B.; Uriarte, M.; McGuire, K.L.; Thompson, J.; Zimmerman, J. Arbuscular mycorrhizal fungal diversity and natural enemies promote coexistence of tropical tree species. Ecology 2017, 98, 712–720. [Google Scholar] [CrossRef] [Green Version]
- Davison, J.; Moora, M.; Öpik, M.; Adholeya, A.; Ainsaar, L.; Bâ, A.; Burla, S.; Diedhiou, A.G.; Hiiesalu, I.; Jairus, T.; et al. Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism. Science 2015, 349, 970–973. [Google Scholar] [CrossRef] [Green Version]
- Nasto, M.K.; Osborne, B.B.; Lekberg, Y.; Asner, G.P.; Balzotti, C.S.; Porder, S.; Taylor, P.G.; Townsend, A.R.; Cleveland, C.C. Nutrient acquisition, soil phosphorus partitioning and competition among trees in a lowland tropical rain forest. New Phytol. 2017, 214, 1506–1517. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.B.; Lee, D.J.; Dweikat, I.M.; Wedin, D.A.; Yuen, G.Y.; Drijber, R.A. Molecular diversity of arbuscular mycorrhizae in roots of Juniperus virginiana invasive to grasslands. Soil Sci. Soc. Am. J. 2017, 81, 526–536. [Google Scholar] [CrossRef]
- Shi, Z.; Chen, Y.; Hou, X.; Gao, S.; Wang, F. Arbuscular mycorrhizal fungi associated with tree peony in 3 geographic locations in China. Turk. J. Agric. For. 2013, 37, 726–733. [Google Scholar] [CrossRef]
- Steidinger, B.S.; Crowther, T.W.; Liang, J.; van Nuland, M.E.; Werner, G.D.A.; Reich, P.B.; Nabuurs, G.J.; de-Miguel, S.; Zhou, M.; Picard, N.; et al. Climatic controls of decomposition drive the global biogeography of forest-tree symbioses. Nature 2019, 569, 404. [Google Scholar] [CrossRef]
- Santiago, L.S. Nutrient limitation of eco-physiological processes in tropical trees. Trees 2015, 29, 1291–1300. [Google Scholar] [CrossRef] [Green Version]
- Averill, C.; Turner, B.L.; Finzi, A.C. Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature 2014, 505, 543–545. [Google Scholar] [CrossRef]
- Camenzind, T.; Homeier, J.; Dietrich, K.; Hempel, S.; Hertel, D.; Krohn, A.; Leuschner, C.; Oelmann, Y.; AxelOlsson, P.; Suárez, J.P.; et al. Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradient in tropical montane forests. Soil Biol. Biochem. 2016, 94, 37–47. [Google Scholar] [CrossRef]
- Alexander, I.; Selosse, M.A. Mycorrhizas in tropical forests: A neglected research imperative. New Phytol. 2009, 182, 14–16. [Google Scholar] [CrossRef]
- Vieira, L.C.; da Silva, D.K.A.; da Silva, I.R.; Gonçalves, C.M.; de Assis, D.M.A.; Oehl, F.; da Silva, G.A. Ecological aspects of arbuscular mycorrhizal fungal communities in different habitat types of a Brazilian mountainous area. Ecol. Res. 2019, 34, 182–192. [Google Scholar] [CrossRef]
- Vieira, L.C.; da Silva, D.K.A.; de Melo, M.A.C.; Escobar, I.E.C.; Oehl, F.; da Silva, G.A. Edaphic factors influence the distribution of arbuscular mycorrhizal fungi along an altitudinal gradient of a tropical mountain. Microb. Ecol. 2019, 1–10. [Google Scholar] [CrossRef]
- Wagg, C.; Husband, B.C.; Green, D.S.; Massicotte, H.B.; Peterson, R.L. Soil microbial communities from an elevational cline differ in their effect on conifer seedling growth. Plant Soil 2011, 340, 491–504. [Google Scholar] [CrossRef]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Lal, R.; Wielopolski, L.; Martin, M.Z.; Ebinger, M.H. Evaluation of Different Soil Carbon Determination Methods. Crit. Rev. Plant Sci. 2009, 28, 164–178. [Google Scholar] [CrossRef]
- Zobeck, T.M.; Baddock, M.; Van Pelt, R.S.; Tatarko, J.; Acosta-Martinez, V. Soil property effects on wind erosion of organic soils. Aeolian Res. 2013, 10, 43–51. [Google Scholar] [CrossRef]
- Wang, H.M.; Wang, W.J.; Chen, H.; Zhang, Z.; Mao, Z.; Zu, Y.G. Temporal changes of soil physic-chemical properties at different soil depths during larch afforestation by multivariate analysis of covariance. Ecol. Evol. 2014, 4, 1039–1048. [Google Scholar] [CrossRef]
- Uparse OUT Clustering. Available online: http://drive5.com/uparse/ (accessed on 8 May 2019).
- RDP Classifier Algorithm. Available online: http://rdp.cme.msu.edu/ (accessed on 18 April 2019).
- Jin, Y.; Liu, Y.; Zhao, L.; Zhao, F.; Feng, J.; Li, S.; Chen, H.; Sun, J.; Zhu, B.; Geng, R.; et al. Gut microbiota in patients after surgical treatment for colorectal cancer. Environ. Microbiol. 2019, 21, 772–783. [Google Scholar] [CrossRef]
- Xiong, W.; Zhao, Q.Y.; Xue, C.; Xun, W.B.; Zhao, J.; Wu, H.S.; Li, R.; Shen, Q.R. Comparison of Fungal Community in Black Pepper-Vanilla and Vanilla Monoculture Systems Associated with Vanilla Fusarium Wilt Disease. Front. Microbiol. 2016, 7, 117. [Google Scholar] [CrossRef]
- Majorbio. Available online: www.i-sanger.com/ (accessed on 16 June 2019).
- Öpik, M.; Zobel, M.; Cantero, J.J.; Davison, J.; Facelli, J.M.; Hiiesalu, I.; Jairus, T.; Kalwij, J.M.; Koorem, K.; Leal, M.E.; et al. Global sampling of plant roots expands the described molecular diversity of arbuscular mycorrhizal fungi. Mycorrhiza 2013, 2, 411–430. [Google Scholar] [CrossRef]
- Qiu, Z.J.; Qiu, J.R.; Zhou, G.Y.; Liu, H.W.; Li, G.M. Comparative analysis of meteorological elements between Hainan’s Diaoluoshan and Jiangfengling tropical forest region. Ecol. Sci. 2004, 23, 338–341. [Google Scholar]
- Li, X.; Gai, J.; Cai, X.; Li, X.; Christie, P.; Zhang, F.; Zhang, J. Molecular diversity of arbuscular mycorrhizal fungi associated with two co-occurring perennial plant species on a Tibetan altitudinal gradient. Mycorrhiza 2014, 24, 95–107. [Google Scholar] [CrossRef]
- Liu, M.; Zheng, R.; Bai, S.; Bai, Y.; Wang, J. Slope aspect influences arbuscular mycorrhizal fungus communities in arid ecosystems of the Daqingshan Mountains, Inner Mongolia, North China. Mycorrhiza 2017, 27, 189–200. [Google Scholar] [CrossRef]
- Gai, J.P.; Tian, H.; Yang, F.Y.; Christie, P.; Li, X.L.; Klironomosa, J.N. Arbuscular mycorrhizal fungal diversity along a Tibetan elevation gradient. Pedobiologia 2012, 55, 145–151. [Google Scholar] [CrossRef]
- Schappe, T.; Albornoz, F.E.; Turner, B.L.; Neat, A.; Condit, R. The role of soil chemistry and plant neighbourhoods in structuring fungal communities in three Panamanian rainforests. J. Ecol. 2017, 105, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Mickan, B.S.; Hart, M.M.; Solaiman, Z.M.; Jenkins, S.; Siddique, K.H.; Abbott, L.K. Molecular divergence of fungal communities in soil, roots and hyphae highlight the importance of sampling strategies. Rhizosphere 2017, 4, 104–111. [Google Scholar] [CrossRef]
- Lugo, M.A.; Ferrero, M.; Menoyo, E.; Estévez, M.C.; Siñeriz, F.; Anton, A. Arbuscular mycorrhizal fungi and rhizospheric bacteria diversity along an altitudinal gradient in South American Puna Grassland. Microb. Ecol. 2008, 55, 705–713. [Google Scholar] [CrossRef]
- Egan, C.P.; Callaway, R.M.; Hart, M.M.; Pither, J.; Klironomos, J. Phylogenetic structure of arbuscular mycorrhizal fungal communities along an elevation gradient. Mycorrhiza 2017, 27, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Bever, J.D. Plant preferential allocation and fungal reward decline with soil phosphorus enrichment: Implications for evolution of the arbuscular mycorrhizal mutualism. Ecosphere 2016, 7, e01256. [Google Scholar] [CrossRef]
- Moora, M.; Davison, J.; Öpik, M.; Metsis, M.; Saks, Ü.; Jairus, T.; Vasar, M.; Zobel, M. Anthropogenic land use shapes the composition and phylogenetic structure of soil arbuscular mycorrhizal fungal communities. FEMS Microbiol. Ecol. 2014, 90, 609–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, I.R.; de Mello, C.M.A.; Neto, R.A.F.; da Silva, D.K.A.; de Melo, A.L.; Oehl, F.; Costa Maia, L. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl. Soil Ecol. 2014, 84, 166–175. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Genus | Number of OTUs | Total OTUs | |
|---|---|---|---|---|---|
| Mt. JF | Mt. DL | ||||
| Archaeosporales | Ambisporaceae | Ambispora | 1 | 1 | 1 |
| Archaeospora | 3 | 5 | 6 | ||
| Unclassified | 2 | 2 | 2 | ||
| Unclassified | 1 | 1 | |||
| Diversisporales | Acaulosporaceae | Acaulospora | 13 | 18 | 19 |
| Diversisporaceae | Diversispora | 6 | 1 | 7 | |
| Unclassified | 1 | 2 | 2 | ||
| Gigasporaceae | Scutellospora | 15 | 13 | 16 | |
| Unclassified | 5 | 6 | 7 | ||
| Glomerales | Glomeraceae | Glomus | 161 | 196 | 218 |
| Paraglomerales | Paraglomeraceae | Paraglomus | 2 | 1 | 3 |
| unclassified | 26 | 16 | 34 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Z.; Yin, K.; Wang, F.; Mickan, B.S.; Wang, X.; Zhou, W.; Li, Y. Alterations of Arbuscular Mycorrhizal Fungal Diversity in Soil with Elevation in Tropical Forests of China. Diversity 2019, 11, 181. https://doi.org/10.3390/d11100181
Shi Z, Yin K, Wang F, Mickan BS, Wang X, Zhou W, Li Y. Alterations of Arbuscular Mycorrhizal Fungal Diversity in Soil with Elevation in Tropical Forests of China. Diversity. 2019; 11(10):181. https://doi.org/10.3390/d11100181
Chicago/Turabian StyleShi, Zhaoyong, Kejing Yin, Fayuan Wang, Bede S. Mickan, Xugang Wang, Wenli Zhou, and Yajuan Li. 2019. "Alterations of Arbuscular Mycorrhizal Fungal Diversity in Soil with Elevation in Tropical Forests of China" Diversity 11, no. 10: 181. https://doi.org/10.3390/d11100181
APA StyleShi, Z., Yin, K., Wang, F., Mickan, B. S., Wang, X., Zhou, W., & Li, Y. (2019). Alterations of Arbuscular Mycorrhizal Fungal Diversity in Soil with Elevation in Tropical Forests of China. Diversity, 11(10), 181. https://doi.org/10.3390/d11100181