First Record of Amphioxus Branchiostoma californiense (Amphioxiformes: Branchiostomatidae) Adjacent to a Shallow Submarine Hydrothermal System at Banderas Bay (Mexico)


Abstract
1. Introduction
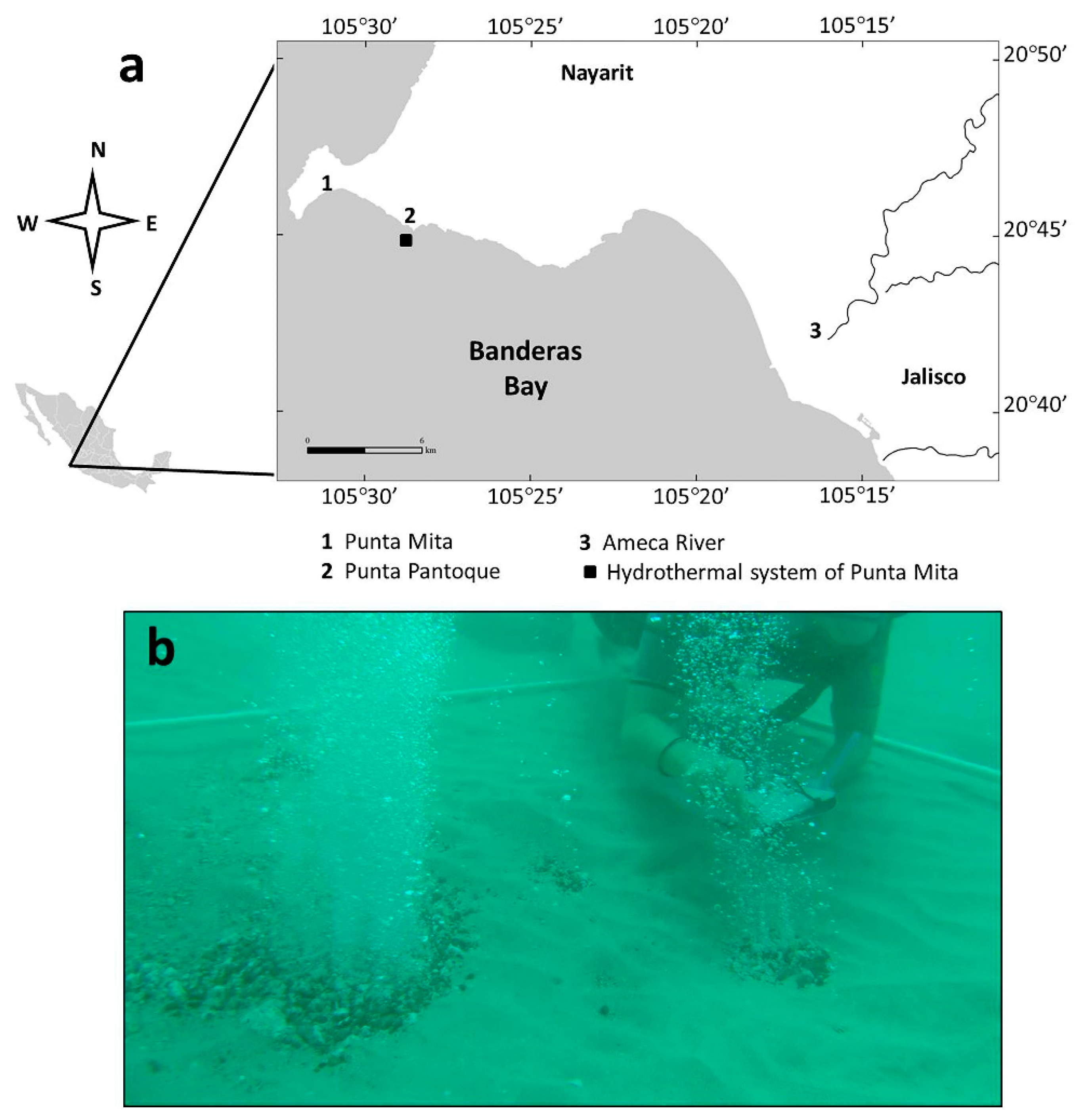
2. Materials and Methods
3. Results
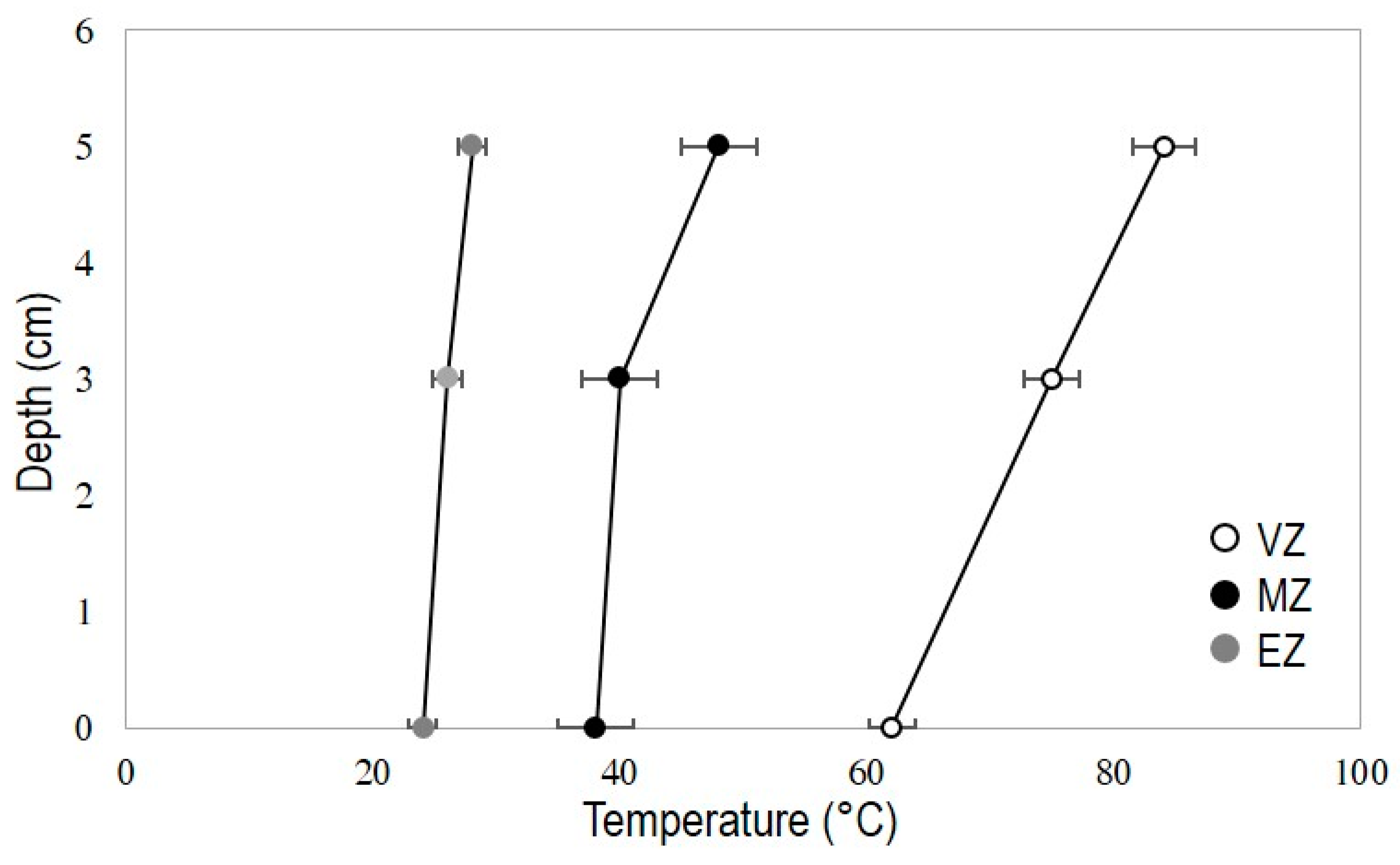
3.1. Pore-Water and Marine Sediments Measurements
3.2. Diagnosis
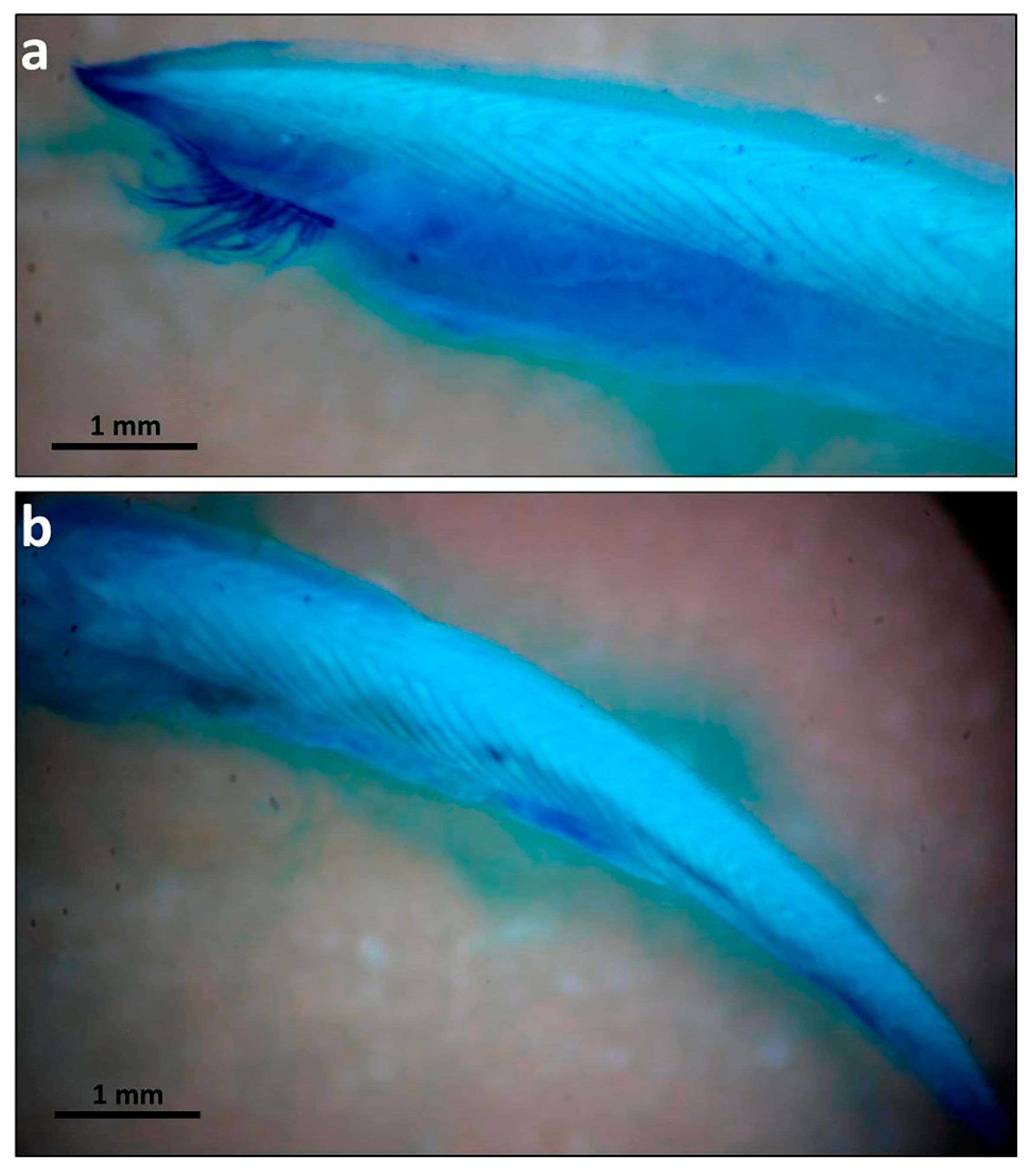
3.3. External Morphology
3.4. Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Del Moral-Flores, L.F.; Guadarrama-Martínez, M.Á.; Flores-Coto, C. Composición taxonómica y distribución de los cefalocordados (Cephalochordata: Amphioxiformes) en México. Lat. Am. J. Aquat. Res. 2016, 44, 497–503. [Google Scholar]
- Kirkaldy, J.W. A revision of the genera and species of the Branchiostomidae. Q. J. Microsc. Sci. 1895, 37, 303–323. [Google Scholar]
- Poss, S.G.; Boschung, H.T. Lancelets (Cephalochordata: Branchiostomatidae): how many species are valid? Isr. J. Zool. 1996, 42, 13–66. [Google Scholar]
- Nishikawa, T. A New Deep-water Lancelet (Cephalochordata) from off Cape Nomamisaki, SW Japan, with a Proposal of the Revised System Recovering the Genus Asymmetron. Zoolog. Sci. 2004, 21, 1131–1136. [Google Scholar] [CrossRef][Green Version]
- Nelson, J.S. Fishes of the World, 4th ed.; Wiley: New York, NY, USA, 2006; ISBN 9780471250319. [Google Scholar]
- Yu, J.K.S.; Holland, L.Z. Cephalochordates (amphioxus or lancelets): a model for understanding the evolution of chordate characters. Cold Spring Harb. Protoc. 2009, 4, 1084–1092. [Google Scholar] [CrossRef]
- Galván-Villa, C.M.; Ríos-Jara, E.; Ayón-Parente, M. New records of the Californian lancelet Branchiostoma californiense (Cephalochordata: Branchiostomidae) from the Pacific coast of Mexico. Rev. Mex. Biodivers. 2017, 88, 995–998. [Google Scholar] [CrossRef]
- Kon, T.; Nohara, M.; Yamanoue, Y.; Fujiwara, Y.; Nishida, M.; Nishikawa, T. Phylogenetic position of a whale-fall lancelet (Cephalochordata) inferred from whole mitochondrial genome sequences. BMC Evol. Biol. 2007, 7, 1–12. [Google Scholar] [CrossRef]
- Richardson, B.; McKenzie, A. Taxonomy and distribution of Australian Cephalochordates (Chordata: Cephalochordata). Invertebr. Taxon. 1994, 8, 1443–1459. [Google Scholar] [CrossRef]
- Gibbs, P.E.; Wickstead, J.H. The myotome formula of the lancelet Epigonichthys lucayanum (Acrania): can variations be related to larval dispersion patterns? J. Nat. Hist. 1996, 30, 615–627. [Google Scholar] [CrossRef]
- Da Silva, L.F.B.; Tavares, M.; Soares-Gomes, A. Population structure of the lancelet Branchiostoma caribaeum (Cephalochordata: Branchiostomidae) in the Baía de Guanabara, Rio de Janeiro, southeastern Brazil. Rev. Bras. Zool. 2008, 25, 617–623. [Google Scholar] [CrossRef]
- Florkin, M.; Scheer, B.T. Deuterostomians, Cyclostomes, and Fishes, 1st ed.; Academic Press: New York, NY, USA, 1974. [Google Scholar]
- Garcia-Fernàndez, J.; Benito-Gutiérrez, È. It’s a long way from amphioxus: descendants of the earliest chordate. BioEssays 2009, 31, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Canet, C.; Prol-Ledesma, R.M. Mineralizing processes at shallow submarine hydrothermal vents: Examples from Mexico. Spec. Pap. Geol. Soc. Am. 2007, 422, 359–376. [Google Scholar]
- Prol Ledesma, R.M.; Juarez M, G. Geothermal map of Mexico. J. Volcanol. Geotherm. Res. 1986, 28, 351–362. [Google Scholar]
- Canet, C.; Prol-Ledesma, R.M. Procesos de mineralización en manantiales hidrotermales submarinos someros. Ejemplos en México. Boletín la Soc. Geológica Mex. 2006, 58, 83–102. [Google Scholar] [CrossRef]
- Núñez-Cornú, F.J.; Prol-Ledesma, R.M.; Cupul-Magaña, A.; Suárez-Plascencia, C. Near shore submarine hydrothermal activity in Bahia Banderas, western Mexico. Geofis. Int. 2000, 39, 171–178. [Google Scholar]
- Prol-Ledesma, R.M.; Canet, C.; Melgarejo, J.C.; Tolson, G.; Rubio-Ramos, M.A.; Cruz-Ocampo, J.C.; Ortega-Osorio, A.; Torres-Vera, M.A.; Reyes, A. Cinnabar deposition in submarine coastal hydrothermal vents, Pacific margin of Central Mexico. Econ. Geol. 2002, 97, 1331–1340. [Google Scholar] [CrossRef]
- Hubbs, C.L. A list of the lancelets of the world with diagnoses of five new species of Branchiostoma. Occas. Pap. Museum Zool. 1922, 105, 1–16. [Google Scholar]
- Vargas, J.A.; Dean, H.K. On Branchiostoma californiense (Cephalochordata) from the Gulf of Nicoya estuary, Costa Rica. Rev. Biol. Trop. 2010, 58, 1143–1148. [Google Scholar] [CrossRef][Green Version]
- Laudien, J.; Rojo, M.E.; Oliva, M.E.; Arntz, W.E.; Thatje, S. Sublittoral soft bottom communities and diversity of Mejillones Bay in northern Chile (Humboldt Current upwelling system). Helgol. Mar. Res. 2007, 61, 103–116. [Google Scholar] [CrossRef]
- Gilchrist, J.D.F. III.—A new species of Amphioxus from the Bazaruto Islands (Branchiostoma bazarutense). Ann. Mag. Nat. Hist. 1923, 12, 62–65. [Google Scholar] [CrossRef]
- Prol-Ledesma, R.M.; Dando, P.R.; de Ronde, C.E.J. Special issue on “shallow-water hydrothermal venting”. Chem. Geol. 2005, 224, 1–4. [Google Scholar] [CrossRef]
- Tarasov, V.G.; Gebruk, A.V.; Mironov, A.N.; Moskalev, L.I. Deep-sea and shallow-water hydrothermal vent communities: Two different phenomena? Chem. Geol. 2005, 224, 5–39. [Google Scholar] [CrossRef]
- Melwani, A.R.; Kim, S.L. Benthic infaunal distributions in shallow hydrothermal vent sediments. Acta Oecol. 2008, 33, 162–175. [Google Scholar] [CrossRef]
- Carpizo-Ituarte, E.; Vizcaíno-Ochoa, V.; Chi-Barragán, G.; Tapia-Vázquez, O.; Cupul-Magaña, A.L.; Medina-Rosas, P. Evidencia de reproducción sexual en los corales hermatípicos Pocillopora damicornis, Porites panamensis y Pavona gigantea en Bahía de Banderas, Pacífico mexicano. Ciencias Mar. 2011, 37, 97–112. [Google Scholar] [CrossRef][Green Version]




{kind=link}
{kind=link}
{kind=link}
| Zone | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| VZ | MZ | EZ | ||||||||
| Sample | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |
| Sampled cephalochordates | 0 | 0 | 0 | 2 | 0 | 1 | 1 | 0 | 4 | 8 |
| Zone | pH | Salinity (ppt) | Water Temperature (°C) |
|---|---|---|---|
| VZ | 7.67 ± 0.01 | 17.2 ± 0.07 | 89 ± 0.47 |
| MZ | 8.04 ± 0.00 | 34.9 ± 0.04 | 26.8 ± 0.09 |
| EZ | 8.05 ± 0.00 | 35.8 ± 0.02 | 25.6 ± 0.21 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Uribe, M.C.; Chávez-Dagostino, R.M.; Del Moral-Flores, L.F.; Bravo-Olivas, M.L. First Record of Amphioxus Branchiostoma californiense (Amphioxiformes: Branchiostomatidae) Adjacent to a Shallow Submarine Hydrothermal System at Banderas Bay (Mexico). Diversity 2019, 11, 227. https://doi.org/10.3390/d11120227
Rodríguez-Uribe MC, Chávez-Dagostino RM, Del Moral-Flores LF, Bravo-Olivas ML. First Record of Amphioxus Branchiostoma californiense (Amphioxiformes: Branchiostomatidae) Adjacent to a Shallow Submarine Hydrothermal System at Banderas Bay (Mexico). Diversity. 2019; 11(12):227. https://doi.org/10.3390/d11120227
Chicago/Turabian StyleRodríguez-Uribe, María Carolina, Rosa María Chávez-Dagostino, Luis Fernando Del Moral-Flores, and Myrna Leticia Bravo-Olivas. 2019. "First Record of Amphioxus Branchiostoma californiense (Amphioxiformes: Branchiostomatidae) Adjacent to a Shallow Submarine Hydrothermal System at Banderas Bay (Mexico)" Diversity 11, no. 12: 227. https://doi.org/10.3390/d11120227
APA StyleRodríguez-Uribe, M. C., Chávez-Dagostino, R. M., Del Moral-Flores, L. F., & Bravo-Olivas, M. L. (2019). First Record of Amphioxus Branchiostoma californiense (Amphioxiformes: Branchiostomatidae) Adjacent to a Shallow Submarine Hydrothermal System at Banderas Bay (Mexico). Diversity, 11(12), 227. https://doi.org/10.3390/d11120227