Diatoms Dominate and Alter Marine Food-Webs When CO2 Rises

 , , ,
, , , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Site, Carbonate Chemistry, and Nutrients
2.2. Field Survey
2.3. Photophysiology and Production
2.4. Associated Fauna
2.5. Statistical Analysis
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nelson, D.M.; Tréguer, P.; Brzezinski, M.A.; Leynaert, A.; Quéguiner, B. Production and dissolution of biogenic silica in the ocean: Revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycles 1995, 9, 359–372. [Google Scholar] [CrossRef]
- Harvey, B.P.; Gwynn-Jones, D.; Moore, P.J. Meta-analysis reveals complex marine biological responses to the interactive effects of ocean acidification and warming. Ecol. Evol. 2013, 3, 1016–1030. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Beardall, J.; Häder, D.-P.; Hall-Spencer, J.M.; Gao, G.; Hutchins, D.A. Effects of ocean acidification on marine photosynthetic organisms Under the concurrent influences of warming, UV radiation, and deoxygenation. Front. Mar. Sci. 2019, 6, 322. [Google Scholar] [CrossRef]
- Brandenburg, K.M.; Velthuis, M.; Van de Waal, D.B. Meta-analysis reveals enhanced growth of marine harmful algae from temperate regions with warming and elevated CO2 levels. Glob. Chang. Biol. 2019, 25, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Bach, L.T.; Taucher, J. CO2 effects on diatoms: A synthesis of more than a decade of ocean acidification experiments with natural communities. Ocean Sci. 2019, 15, 1159–1175. [Google Scholar] [CrossRef] [Green Version]
- Reinfelder, J.R. Carbon concentrating mechanisms in eukaryotic marine phytoplankton. Annu. Rev. Mar. Sci. 2011, 3, 291–315. [Google Scholar] [CrossRef] [Green Version]
- Bach, L.T.; Hernández-Hernández, N.; Taucher, J.; Spisla, C.; Sforna, C.; Riebesell, U.; Arístegui, J. Effects of elevated CO2 on a natural diatom community in the subtropical NE Atlantic. Front. Mar. Sci. 2019, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Macreadie, P.I.; Anton, A.; Raven, J.A.; Beaumont, N.; Connolly, R.M.; Friess, D.A.; Kelleway, J.J.; Kennedy, H.; Kuwae, T.; Lavery, P.S.; et al. The future of Blue Carbon science. Nat. Commun. 2019, 10, 3998. [Google Scholar] [CrossRef] [Green Version]
- Sarthou, G.; Timmermans, K.R.; Blain, S.; Tréguer, P. Growth physiology and fate of diatoms in the ocean: A review. Iron Resour. Ocean. Nutr. Adv. Glob. Environ. Simul. 2005, 53, 25–42. [Google Scholar] [CrossRef]
- Flynn, K.J.; Blackford, J.C.; Baird, M.E.; Raven, J.A.; Clark, D.R.; Beardall, J.; Brownlee, C.; Fabian, H.; Wheeler, G.L. Changes in pH at the exterior surface of plankton with ocean acidification. Nat. Clim. Chang. 2012, 2, 510–513. [Google Scholar] [CrossRef]
- Tortell, P.D.; Payne, C.D.; Li, Y.; Trimborn, S.; Rost, B.; Smith, W.O.; Riesselman, C.; Dunbar, R.B.; Sedwick, P.; DiTullio, G.R. CO2 sensitivity of Southern Ocean phytoplankton. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Hare, C.E.; Rose, J.M.; Handy, S.M.; DiTullio, G.R.; Lee, P.A.; Smith, W.O.; Peloquin, J.; Tozzi, S.; Sun, J.; et al. Interactive effects of iron, irradiance and CO2 on Ross Sea phytoplankton. Deep Sea Res. Part Oceanogr. Res. Pap. 2010, 57, 368–383. [Google Scholar] [CrossRef]
- Sommer, U.; Stibor, H.; Katechakis, A.; Sommer, F.; Hansen, T. Pelagic food web configurations at different levels of nutrient richness and their implications for the ratio fish production:primary production. Hydrobiologia 2002, 484, 11–20. [Google Scholar] [CrossRef]
- Feng, Y.; Hare, C.E.; Leblanc, K.; Rose, J.M.; Zhang, Y.; DiTullio, G.R.; Lee, P.A.; Wilhelm, S.W.; Rowe, J.M.; Sun, J.; et al. Effects of increased pCO2 and temperature on the North Atlantic spring bloom. I. The phytoplankton community and biogeochemical response. Mar. Ecol. Prog. Ser. 2009, 388, 13–25. [Google Scholar] [CrossRef]
- Eggers, S.L.; Lewandowska, A.M.; Barcelos, E.; Ramos, J.; Blanco-Ameijeiras, S.; Gallo, F.; Matthiessen, B. Community composition has greater impact on the functioning of marine phytoplankton communities than ocean acidification. Glob. Chang. Biol. 2014, 20, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Taucher, J.; Bach, L.T.; Boxhammer, T.; Nauendorf, A.; Achterberg, E.P.; Algueró-Muñiz, M.; Arístegui, J.; Czerny, J.; Esposito, M.; Guan, W.; et al. Influence of ocean acidification and deep water upwelling on oligotrophic plankton communities in the subtropical North Atlantic: Insights from an in situ mesocosm study. Front. Mar. Sci. 2017, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Bach, L.T.; Alvarez-Fernandez, S.; Hornick, T.; Stuhr, A.; Riebesell, U. Simulated ocean acidification reveals winners and losers in coastal phytoplankton. PLoS ONE 2017, 12, e0188198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, K.G.; Bach, L.T.; Bellerby, R.G.J.; Bermúdez, R.; Büdenbender, J.; Boxhammer, T.; Czerny, J.; Engel, A.; Ludwig, A.; Meyerhöfer, M.; et al. Phytoplankton blooms at increasing levels of atmospheric carbon dioxide: Experimental evidence for negative effects on prymnesiophytes and positive on small picoeukaryotes. Front. Mar. Sci. 2017, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Díaz, A.; Díaz-Pérez, L.; Suárez, L.Á.; Morán, X.A.G.; Teira, E.; Marañón, E. Decrease in the autotrophic-to-heterotrophic biomass ratio of picoplankton in oligotrophic marine waters due to bottle enclosure. Appl. Environ. Microbiol. 2011, 77, 5739. [Google Scholar] [CrossRef] [Green Version]
- Riebesell, U.; Aberle-Malzahn, N.; Achterberg, E.P.; Algueró-Muñiz, M.; Alvarez-Fernandez, S.; Arístegui, J.; Bach, L.T.; Boersma, M.; Boxhammer, T.; Guan, W.; et al. Toxic algal bloom induced by ocean acidification disrupts the pelagic food web. Nat. Clim. Chang. 2018, 8, 1082–1086. [Google Scholar] [CrossRef]
- Fabricius, K.E.; Langdon, C.; Uthicke, S.; Humphrey, C.; Noonan, S.; De’ath, G.; Okazaki, R.; Muehllehner, N.; Glas, M.S.; Lough, J.M. Losers and winners in coral reefs acclimatized to elevated carbon dioxide concentrations. Nat. Clim. Chang. 2011, 1, 165–169. [Google Scholar] [CrossRef]
- Agostini, S.; Harvey, B.P.; Wada, S.; Kon, K.; Milazzo, M.; Inaba, K.; Hall-Spencer, J.M. Ocean acidification drives community shifts towards simplified non-calcified habitats in a subtropical-temperate transition zone. Sci. Rep. 2018, 8, 11354. [Google Scholar] [CrossRef] [Green Version]
- Hall-Spencer, J.M.; Rodolfo-Metalpa, R.; Martin, S.; Ransome, E.; Fine, M.; Turner, S.M.; Rowley, S.J.; Tedesco, D.; Buia, M.-C. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 2008, 454, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Johnson, V.R.; Brownlee, C.; Rickaby, R.E.M.; Graziano, M.; Milazzo, M.; Hall-Spencer, J.M. Responses of marine benthic microalgae to elevated CO2. Mar. Biol. 2011, 160, 1813–1824. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.V.; Brownlee, C.; Milazzo, M.; Hall-Spencer, M.J. Marine microphytobenthic assemblage shift along a natural shallow-water CO2 gradient subjected to multiple environmental stressors. J. Mar. Sci. Eng. 2015, 3, 1425–1447. [Google Scholar] [CrossRef] [Green Version]
- Harvey, B.P.; Agostini, S.; Wada, S.; Inaba, K.; Hall-Spencer, J.M. Dissolution: the Achilles’ heel of the triton shell in an acidifying ocean. Front. Mar. Sci. 2018, 5, 371. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013—The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2013; p. 1535. [Google Scholar]
- Agostini, S.; Wada, S.; Kon, K.; Omori, A.; Kohtsuka, H.; Fujimura, H.; Tsuchiya, Y.; Sato, T.; Shinagawa, H.; Yamada, Y.; et al. Geochemistry of two shallow CO2 seeps in Shikine Island (Japan) and their potential for ocean acidification research. Reg. Stud. Mar. Sci. 2015, 2, 45–53. [Google Scholar] [CrossRef]
- Pierrot, D.; Lewis, E.; Wallace, D.W.R. MS Excel Program Developed for CO2 System Calculations, ORNL/CDIAC-105; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, US Department of Energy: Oak Ridge, TN, USA, 2006. [Google Scholar]
- Mehrbach, C.; Culberson, C.H.; Hawley, J.E.; Pytkowicz, R.M. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 1973, 18, 897–907. [Google Scholar] [CrossRef]
- Dickson, A.G.; Millero, F.J. A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep Sea Res. Part Oceanogr. Res. Pap. 1987, 34, 1733–1743. [Google Scholar] [CrossRef]
- Dickson, A.G. Thermodynamics of the dissociation of boric acid in potassium chloride solutions from 273.15 to 318.15 K. J. Chem. Eng. Data 1990, 35, 253–257. [Google Scholar] [CrossRef]
- Uppström, L.R. The boron/chlorinity ratio of deep-sea water from the Pacific Ocean. Deep Sea Res. Oceanogr. Abstr. 1974, 21, 161–162. [Google Scholar] [CrossRef]
- Hansen, H.; Koroleff, F. Determination of Nutrients. In Methods of Seawater Analysis; Grasshoff, K., Kremling, K., Ehrhardt, M., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2007; pp. 159–228. [Google Scholar]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Porra, R.; Thompson, W.; Kriedemann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta BBA Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Harianto, J.; Carey, N.; Byrne, M. RespR—An R package for the manipulation and analysis of respirometry data. Methods Ecol. Evol. 2019, 10, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing. Dev. Core 2019, 201. Available online: https://www.r-project.org/ (accessed on 10 December 2019).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package (R Package Version 2.5–5). 2019. Available online: https://rdrr.io/cran/vegan/ (accessed on 10 October 2019).
- Kociolek, J.P.; Coste, M.; Ector, L.; Liu, Y.; Kulikovskiy, M.; Lundholm, N.; Ludwig, T.; Potapova, M.; Rimet, F.; Sabbe, K.; et al. DiatomBase Biddulphia Biddulphiana (J.E.Smith) Boyer. 1900. Available online: http://www.diatombase.org/aphia.php?p=taxdetails&id=162952 (accessed on 1 June 2018).
- Round, F.E.; Crawford, R.M.; Mann, D.G. Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; ISBN 978-0-521-36318-1. [Google Scholar]
- Hoagland, K.D.; Rosowski, J.R.; Gretz, M.R.; Roemer, S.C. Diatom extracellular polymeric substances: Function, fine structure, chemistry, and physiology. J. Phycol. 1993, 29, 537–566. [Google Scholar] [CrossRef]
- Galland, G.R.; Pennebaker, S.L. A benthic diatom bloom in the Gulf of California, Mexico. BioInvasions Rec. 2012, 1, 65–69. [Google Scholar] [CrossRef]
- Allen, R.; Summerfield, T.C.; Harvey, B.P.; Agostini, S.; Rastrick, S.P.S.; Hall-Spencer, J.M.; Hoffmann, L.J. The effect of ocean acidification on biofilm community composition and early successional dynamics: A high-throughput sequencing study. Unpublished work, 2019. [Google Scholar]
- Milligan, A.J.; Morel, F.M.M. A proton buffering role for silica in diatoms. Science 2002, 297, 1848–1850. [Google Scholar] [CrossRef]
- Brzezinski, M.A. The Si:C:N ratio of marine diatoms: Interspecific variability and the effects of some environmental variables. J. Phycol. 1985, 21, 347–357. [Google Scholar] [CrossRef]
- Flynn, K.J.; Martin-Jézéquel, V. Modelling Si–N-limited growth of diatoms. J. Plankton Res. 2000, 22, 447–472. [Google Scholar] [CrossRef] [Green Version]
- Underhill, P.A. Nitrate uptake kinetics and clonal variability in the neritic diatom Biddulphia aurita. J. Phycol. 1977, 13, 170–176. [Google Scholar] [CrossRef]
- Qasim, S.Z.; Bhattathiri, P.M.A.; Devassy, V.P. Growth kinetics and nutrient requirements of two tropical marine phytoplankters. Mar. Biol. 1973, 21, 299–304. [Google Scholar] [CrossRef]
- Egge, J.; Aksnes, D. Silicate as regulating nutrient in phytoplankton competition. Mar. Ecol. Prog. Ser. 1992, 83, 281–289. [Google Scholar] [CrossRef]
- Cattano, C.; Agostini, S.; Harvey, B.P.; Wada, S.; Quattrocchi, F.; Turco, G.; Inaba, K.; Hall-Spencer, J.M.; Milazzo, M. Consistent spatio-temporal changes in fish communities as a result of benthic habitat shifts under ocean acidification conditions. Unpublished work, 2019. [Google Scholar]
- Hopkinson, B.M.; Dupont, C.L.; Allen, A.E.; Morel, F.M. Efficiency of the CO2-concentrating mechanism of diatoms. Proc. Natl. Acad. Sci. USA 2011, 108, 3830–3837. [Google Scholar] [CrossRef] [Green Version]
- Young, J.N.; Morel, F.M. Biological oceanography: The CO2 switch in diatoms. Nat. Clim. Chang. 2015, 5, 722. [Google Scholar] [CrossRef]
- Trimborn, S.; Wolf-Gladrow, D.; Richter, K.-U.; Rost, B. The effect of pCO2 on carbon acquisition and intracellular assimilation in four marine diatoms. J. Exp. Mar. Bio. Ecol. 2009, 376, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Gao, K.; Riebesell, U. CO2-induced seawater acidification affects physiological performance of the marine diatom Phaeodactylum tricornutum. Biogeosciences 2010, 7, 2915–2923. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Gao, K. Physiological responses of the marine diatom Thalassiosira pseudonana to increased pCO2 and seawater acidity. Mar. Environ. Res. 2012, 79, 142–151. [Google Scholar] [CrossRef]
- Shi, D.; Hong, H.; Su, X.; Liao, L.; Chang, S.; Lin, W. The physiological response of marine diatoms to ocean acidification: Differential roles of seawater pCO2 and pH. J. Phycol. 2019, 55, 521–533. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Micheli, F.; Gambi, M.C.; Martz, T.R. Divergent ecosystem responses within a benthic marine community to ocean acidification. Proc. Natl. Acad. Sci. USA 2011, 108, 14515–14520. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, S.; Martínez-Crego, B.; Andolina, C.; Massa-Gallucci, A.; Connell, S.D.; Gambi, M.C. Ocean acidification as a driver of community simplification via the collapse of higher-order and rise of lower-order consumers. Sci. Rep. 2017, 7, 4018. [Google Scholar] [CrossRef] [Green Version]
- Harvey, B.P.; Al-Janabi, B.; Broszeit, S.; Cioffi, R.; Kumar, A.; Aranguren-Gassis, M.; Bailey, A.; Green, L.; Gsottbauer, C.M.; Hall, E.F.; et al. Evolution of marine organisms under climate change at different levels of biological organisation. Water 2014, 6, 3545–3574. [Google Scholar] [CrossRef] [Green Version]
- Connell, S.D.; Doubleday, Z.A.; Foster, N.R.; Hamlyn, S.B.; Harley, C.D.G.; Helmuth, B.; Kelaher, B.P.; Nagelkerken, I.; Rodgers, K.L.; Sarà, G.; et al. The duality of ocean acidification as a resource and a stressor. Ecology 2018, 99, 1005–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micheli, F.; Cottingham, K.L.; Bascompte, J.; Bjørnstad, O.N.; Eckert, G.L.; Fischer, J.M.; Keitt, T.H.; Kendall, B.E.; Klug, J.L.; Rusak, J.A. The dual nature of community variability. Oikos 1999, 85, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Kohda, M. Differences in gill raker morphology between two local populations of a benthophagous filter-feeding fish, Goniistius zonatus (Cheilodactylidae). Ichthyol. Res. 2001, 48, 269–273. [Google Scholar] [CrossRef]
- Sazima, I. Field evidence for suspension feeding in Pseudocaranx dentex, with comments on ram filtering in other jacks (Carangidae). Environ. Biol. Fishes 1998, 53, 225–229. [Google Scholar] [CrossRef]
- Sunday, J.M.; Calosi, P.; Dupont, S.; Munday, P.L.; Stillman, J.H.; Reusch, T.B.H. Evolution in an acidifying ocean. Trends Ecol. Evol. 2014, 29, 117–125. [Google Scholar] [CrossRef]
- Hall-Spencer, J.M.; Harvey, B.P. Ocean acidification impacts on coastal ecosystem services due to habitat degradation. Emerg. Top. Life Sci. 2019, 3, 197–206. [Google Scholar]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; O’Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]





{kind=link}
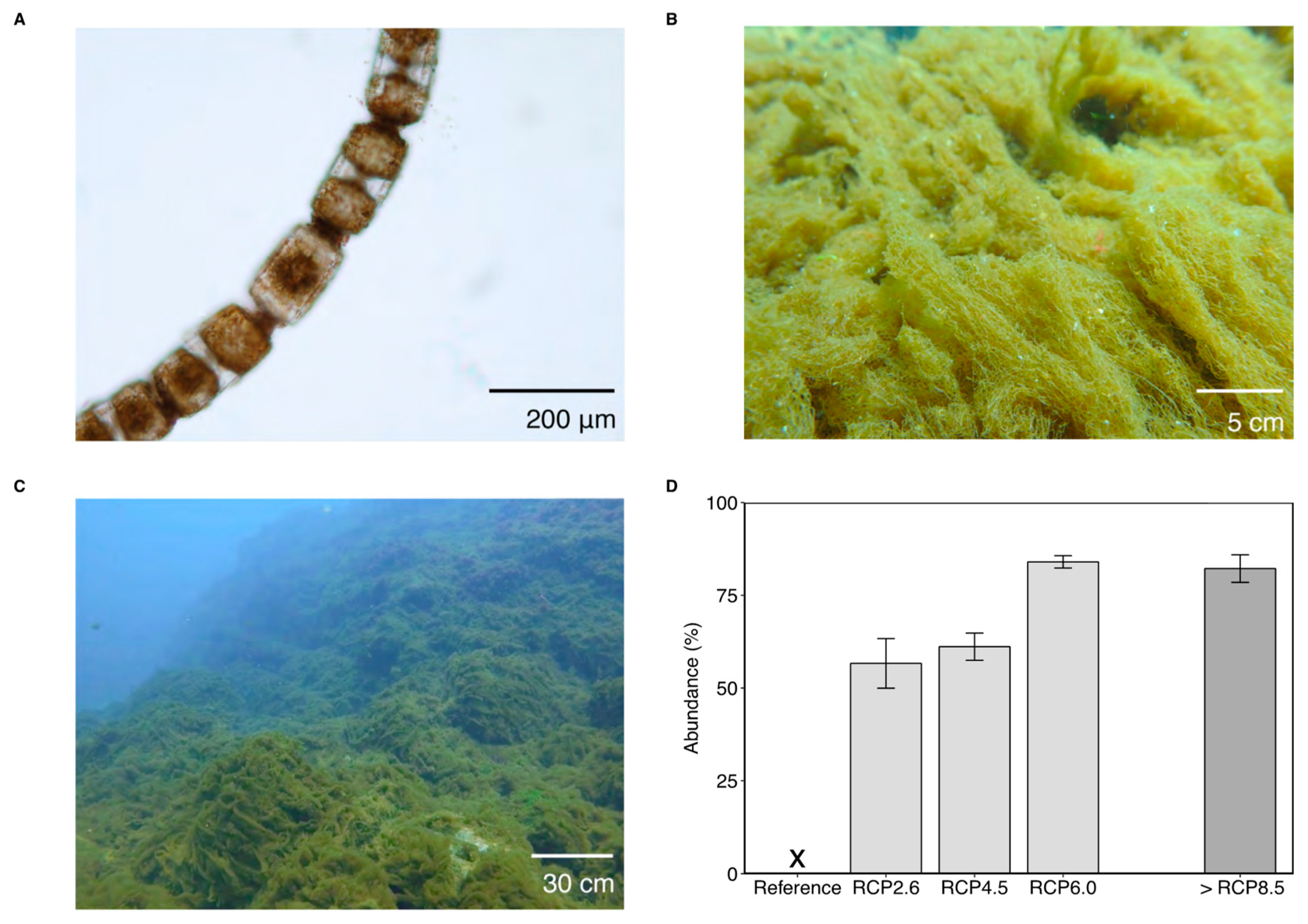{kind=link}
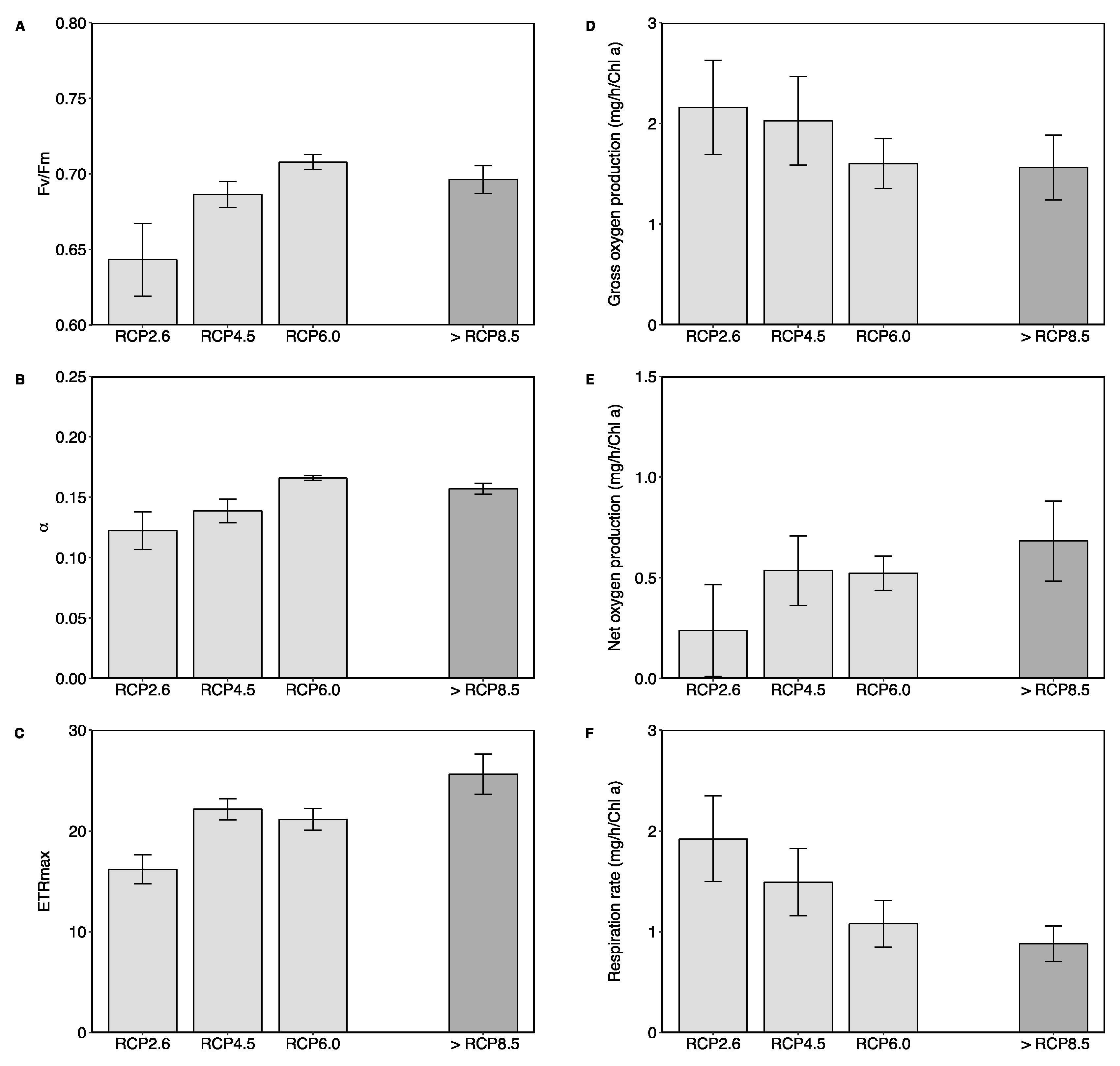{kind=link}
{kind=link}
{kind=link}
| Station | pHT | Temp (°C) | Salinity (psu) | AT (μmol kg−1) | pCO2 (μatm) | DIC (μmol kg−1) | HCO3− (μmol kg−1) | CO32− (μmol kg−1) | Ωcalcite | Ωaragonite |
|---|---|---|---|---|---|---|---|---|---|---|
| Reference | 8.041 | 23.086 | 34.129 | 2281.9 | 409.965 | 2007.341 | 1798.117 | 196.978 | 4.76 | 3.115 |
| 0.067 | 0.603 | 0.741 | 6.80 | 73.383 | 38.944 | 61.612 | 24.859 | 0.596 | 0.392 | |
| RCP 2.6 | 7.983 | 21.437 | 35.056 | 2282.93 | 493.011 | 2044.255 | 1855.972 | 173.103 | 4.144 | 2.703 |
| 0.119 | 1.273 | 0.125 | 6.57 | 158.004 | 53 | 81.439 | 32.771 | 0.781 | 0.501 | |
| RCP 4.5 | 7.809 | 22.701 | 34.455 | 2283.32 | 765.545 | 2122.447 | 1973.165 | 126.296 | 3.043 | 1.99 |
| 0.075 | 0.919 | 0.132 | 18.53 | 158.892 | 27.476 | 38.887 | 15.755 | 0.378 | 0.244 | |
| RCP 6.0 | 7.719 | 22.896 | 34.91 | 2271.84 | 970.706 | 2144.537 | 2008.7 | 106.928 | 2.568 | 1.681 |
| 0.095 | 0.937 | 0.211 | 3.03 | 257.68 | 33.169 | 43.845 | 17.716 | 0.423 | 0.274 | |
| >RCP 8.5 | 7.529 | 22.072 | 34.723 | 2277.62 | 1803.047 | 2218.975 | 2088.23 | 75.92 | 1.823 | 1.19 |
| 0.234 | 1.212 | 0.742 | 20.50 | 1287.448 | 82.982 | 82.43 | 33.368 | 0.799 | 0.519 |
| Site | C | Si | N | P |
|---|---|---|---|---|
| Reference | 9050 | 21 | 13 | 1 |
| RCP 2.6 | 10,564 | 39 | 15 | 1 |
| RCP 4.5 | 10,157 | 37 | 14 | 1 |
| RCP 6.0 | 11,049 | 40 | 15 | 1 |
| >RCP 8.5 | 11,469 | 128 | 12 | 1 |
| Redfield–Brzezinski | 106 | 15 | 16 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harvey, B.P.; Agostini, S.; Kon, K.; Wada, S.; Hall-Spencer, J.M. Diatoms Dominate and Alter Marine Food-Webs When CO2 Rises. Diversity 2019, 11, 242. https://doi.org/10.3390/d11120242
Harvey BP, Agostini S, Kon K, Wada S, Hall-Spencer JM. Diatoms Dominate and Alter Marine Food-Webs When CO2 Rises. Diversity. 2019; 11(12):242. https://doi.org/10.3390/d11120242
Chicago/Turabian StyleHarvey, Ben P., Sylvain Agostini, Koetsu Kon, Shigeki Wada, and Jason M. Hall-Spencer. 2019. "Diatoms Dominate and Alter Marine Food-Webs When CO2 Rises" Diversity 11, no. 12: 242. https://doi.org/10.3390/d11120242
APA StyleHarvey, B. P., Agostini, S., Kon, K., Wada, S., & Hall-Spencer, J. M. (2019). Diatoms Dominate and Alter Marine Food-Webs When CO2 Rises. Diversity, 11(12), 242. https://doi.org/10.3390/d11120242