4.1. Abundance Dynamics in Relation to Changing Conditions
The current density of the common buzzard in our study area was high when compared to other European studies [
8,
10,
13,
42,
43]. It was stable in the first study period (1982–1992; [
20]), and probably at least until 1978 [
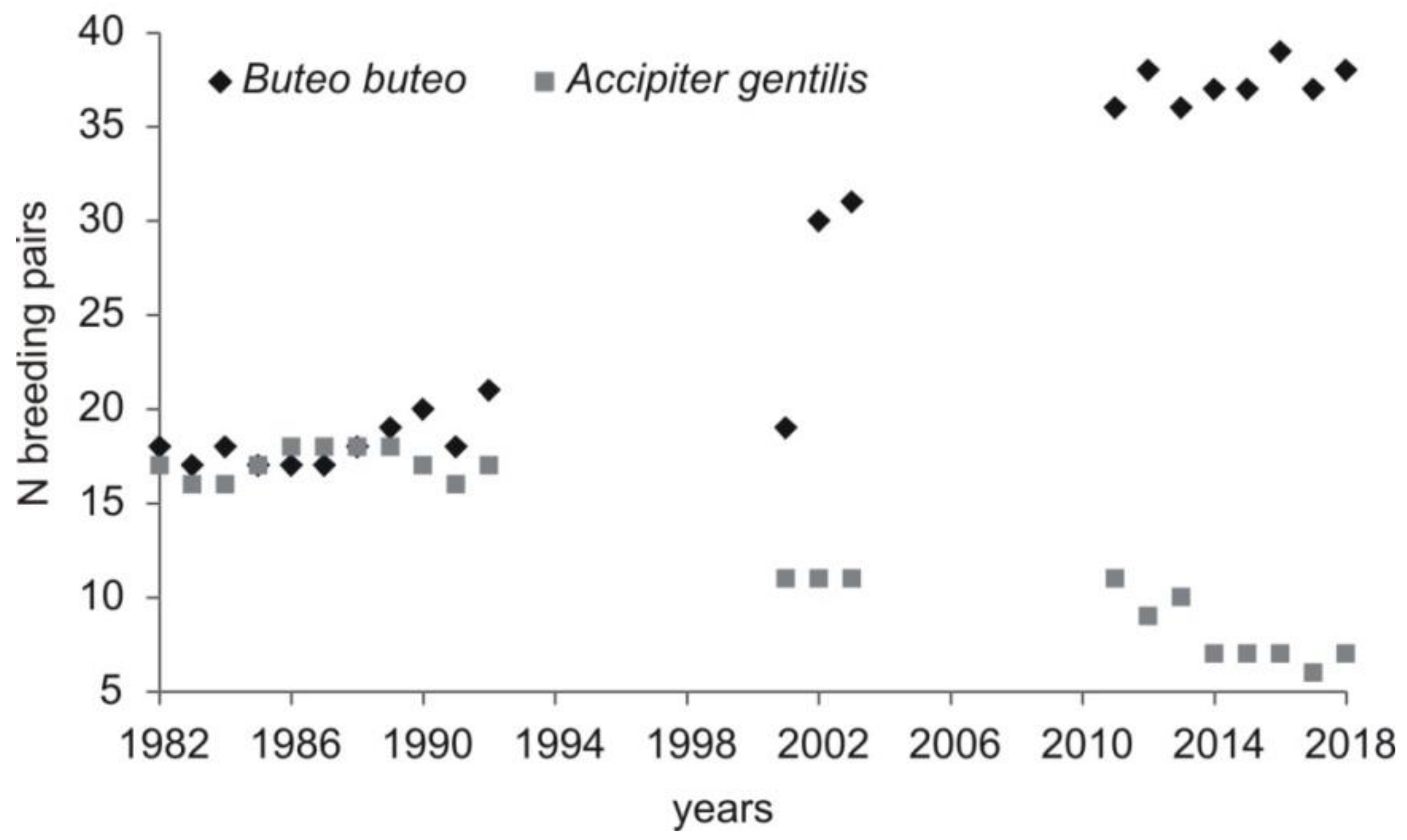
21]. Then, the population started to grow (in the beginning of the 1990s), and this has continued until today. One of the reasons that might have allowed for the population growth was the decrease in the northern goshawk number. Up to 1989, the abundance of the two species was similar, but since 1989, buzzards have always been in larger numbers, and this disproportion was higher every breeding season. The abundance of goshawk dropped to 6–7 pairs [
38], while that of the common buzzard peaked at 37–39 pairs in the last few years (this study). The situation from one of the forest complexes in our study area (no. 7; 70 ha;
Figure 1) shows relations between the two species very well. When goshawk was present in this forest (one pair), two pairs of buzzards nested there. However, before and after this period, with goshawk absent from the forest complex, the number of pairs of buzzards increased to four [
24,
44,
45]. Goshawks can prey on juvenile and adult buzzards (review in Reference [
46]). Indeed, in our study area in 2001–2002, buzzard (mostly juveniles) constituted almost 1% of goshawks’ prey items [
47]. The two species may also compete for food, as the current share of birds in a total consumed biomass by common buzzards was 60%, with more than 18% being domestic pigeons
Columba livia f.
domestica and poultry, the staple food of goshawks in this area [
38]. Niche overlap between the two species was reported to be especially strong in times of low vole abundance, when buzzards switched to birds. This resulted in lower breeding performance of the two species [
20]. Further, in our studies, a relatively high share of birds in diet was noted in time of low rodent availability. The presence of goshawks is known to be the factor that affects nest place choice by buzzards [
17] and negatively influence their breeding parameters [
16,
18,
19]. The other species that may compete with common buzzard is raven. In fact, its population decreased in our study area in previous years [
48]. Moreover, in contrast to the previous study periods [
26,
41], no cases of buzzard persecution (i.e., shooting, cutting of nesting trees, trapping of birds) were recorded in the last few years. In the UK, reduced persecution was one of the factors that allowed a rapid population increase [
9]. Finally, the factor which might have influenced the population of the common buzzard positively was the relatively mild winters in the last decades [
49], as it was shown that snow cover in spring is negatively related to breeding performance [
13], while changing winter weather resulted in higher survival rates, which seemed to account for an increase in population size in Germany [
50].
4.2. Diet Composition and Changes in Prey Availability
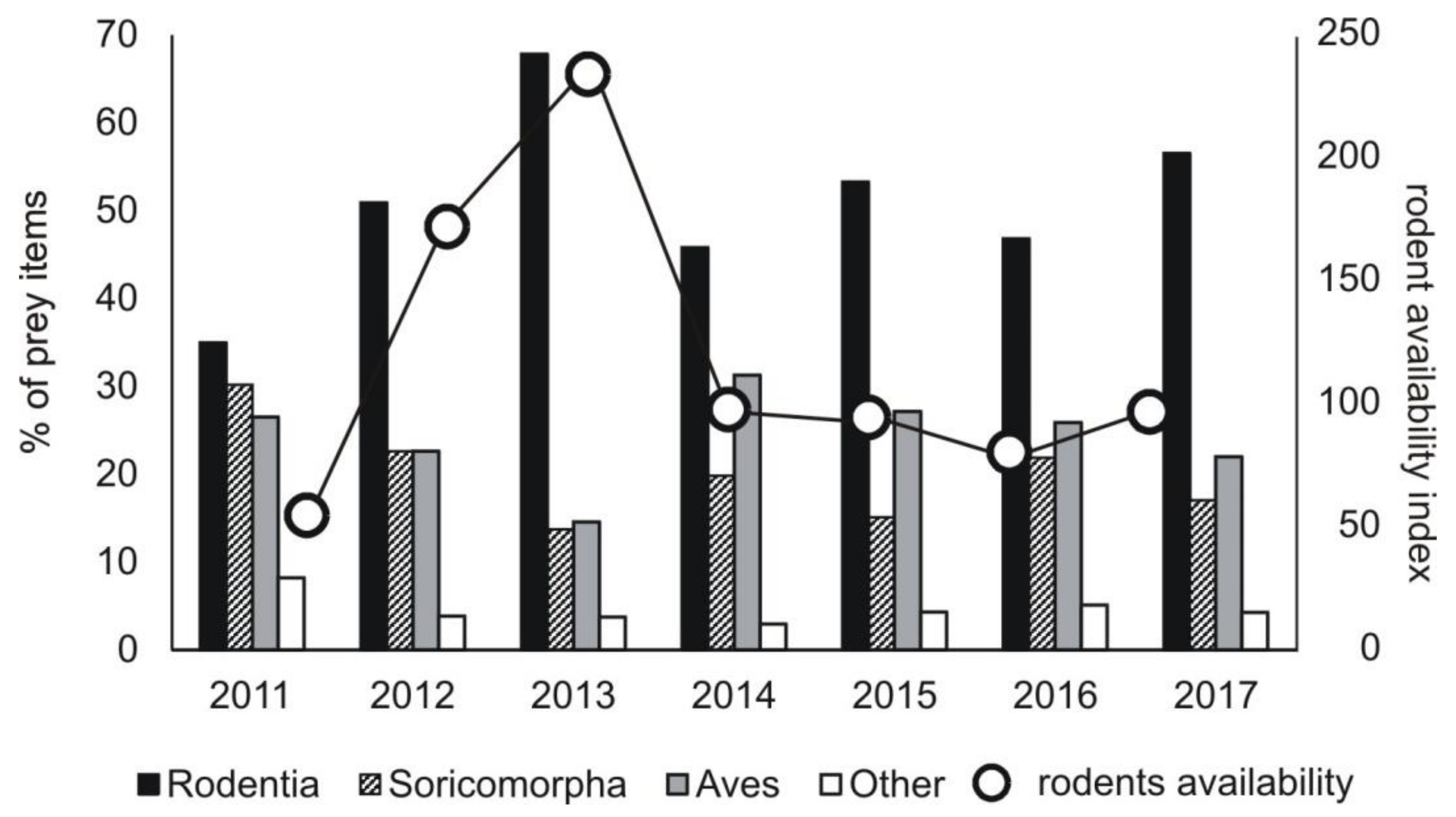
Our results showed important changes in the diet composition of the common buzzard in the last 30 years, which were the result of changes in rodent availability, mainly a decrease in the abundance of voles of
Microtus genus, and dampening of their population cycles. Such a collapse in vole cycles was reported in many countries [
51,
52,
53,
54], including North–Eastern and Western Poland [
55], and referred to
Microtus arvalis,
M. agrestis and
M. oeconomus [
55,
56]. The reasons for this are not fully explained, yet they are usually connected to climatic changes [
57,
58] (but see Reference [
59]). When voles are available and abundant, they may form the bulk of the common buzzard’s diet, i.e., in the farmland of Western Poland, where common vole reached densities of more than 300 ind./ha in the 1970–1980s, buzzards fed mainly on those rodents (over 80% of consumed biomass) [
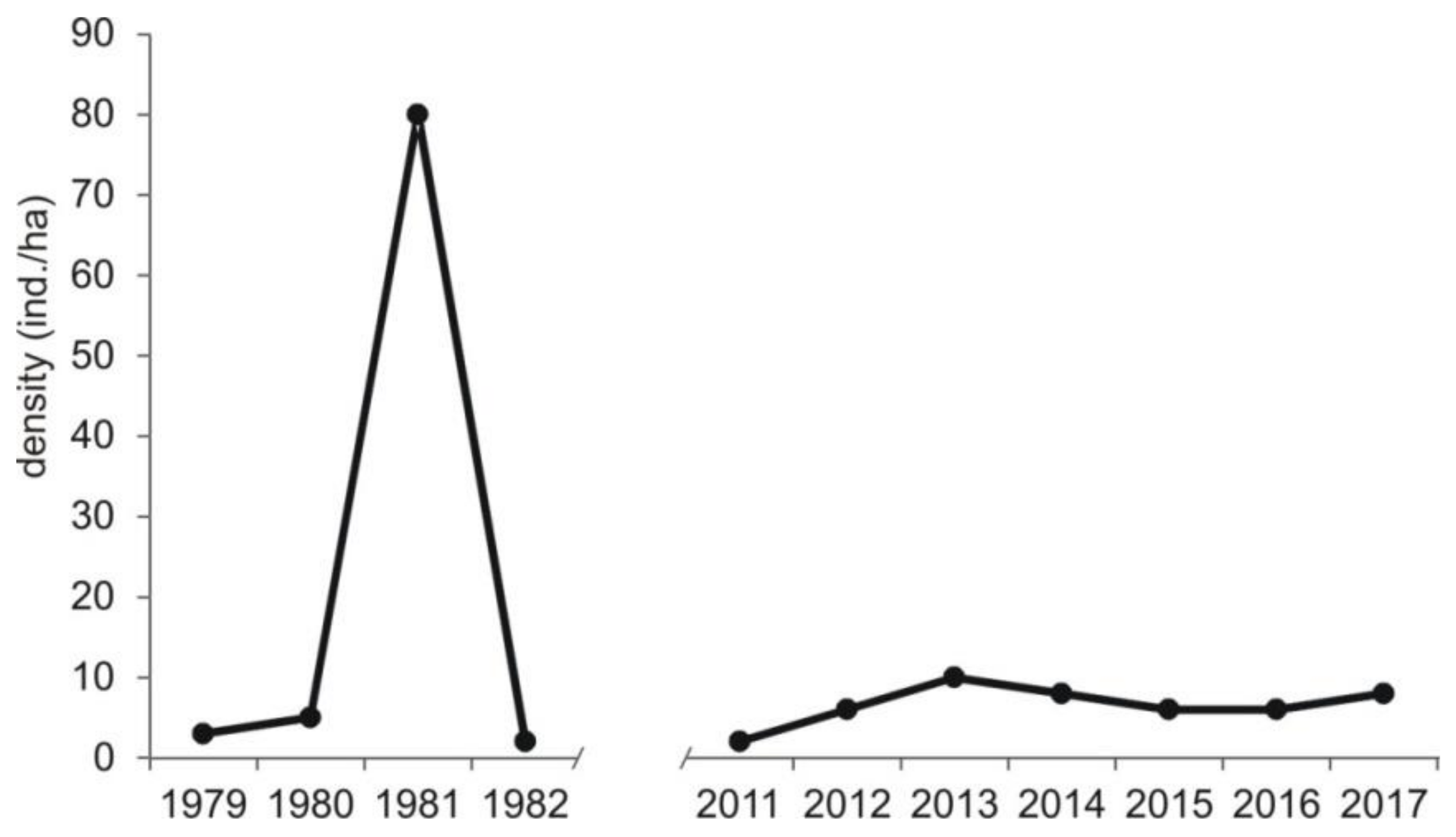
60]. In our study area, the densities of common voles were never that high; nevertheless, in 1981, this species density was higher than 80 ind./ha [
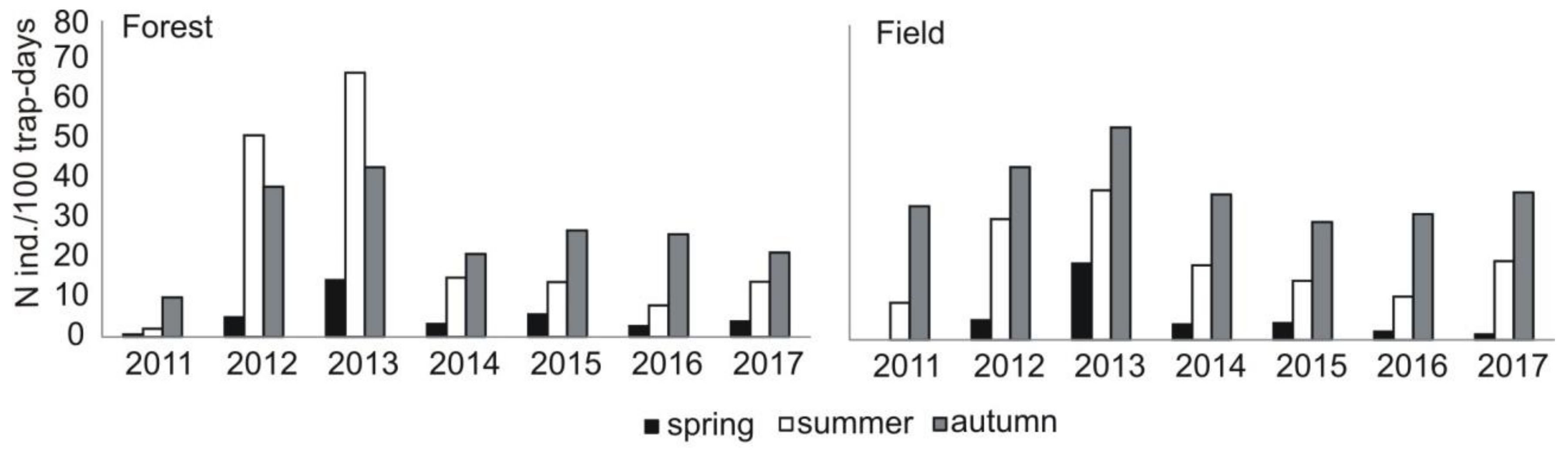
21]. In times of peak density, voles migrated from open to forest areas and even in forest complexes were one of the dominant rodent species [
61]. In the 1980s, mice (
Apodemus and
Mus species) were rather rare in the field rodent community. This changed in the last decade, i.e., in 2011–2017 (current study), the maximum density of voles (three
Microtus species pooled together) did not exceed 10 ind./ha. Further, live trapping of rodents in the first decade of the 20th century did not show high densities of voles [
62]; by contrast, striped-field mice dominated in open areas [
62]. This decrease in vole abundance was reflected by a drop in their share in the common buzzard diet, which decreased by over 40% in the last 35 years, and an increase in the share of striped-field mouse. The striped-field mouse is a highly adaptable species, inhabiting various habitats, i.e., fields, meadows, small woodlots, field-forest ecotone, and also urbanized areas, and expanding in the geographical range and spectrum of habitats where it lives. It is also daytime-active, which correlates with the activity pattern of the common buzzard [
63,
64,
65,
66]. The other mouse species, yellow-necked mouse, was a dominant species among rodents trapped in the forest. What is interesting is that in the field area, yellow-necked mice were trapped in considerable numbers. This was the case mostly in 2013, when their peak abundance was recorded, and mice moved from the forest to neighboring open fields. However, in the diet of the common buzzard, this species was represented marginally, i.e., its share was a bit over 1%.
The observed changes in field rodent community may be the result of climate warming. According to data from local weather station in Rogów, mean temperature increased in the last decades, while snow cover lasted a shorter time [
50,
67]. The other factor may be unfavorable changes in farming practices. In the 1980s, plants preferred by voles, i.e., medicago
Medicago spp., other alfalfa crops, and vegetables [
68] accounted for more than 11% of the whole cultivated area [
21]. Currently, crop structure has changed and the aforementioned plants are not cultivated anymore; the same applies to the cultivation of oats, while potato cultivation was severely reduced. Today, cultivated areas are dominated by wheat, rape, and maize; the last two were unknown in the area 40 years ago.
We showed that common buzzards reacted to changes in rodent availability by switching to other food categories. This raptor is a food opportunist, hunting a wide spectrum of prey species [
69], but focusing on those most abundant and thus most available ones [
70]. In European conditions, these are usually voles
Microtus spp., but in times of drop in their abundance, buzzards hunt an alternative prey, such as soricomorphs, lagomorphs, birds, including forest grouses, amphibians, reptiles, and invertebrates, including earthworms [
7,
71,
72,
73,
74]. The diet composition of the common buzzard depends on the landscape structure. In Belarus, the share of voles
Microtus spp. decreased, while the share of amphibians, reptiles, soricomorphs, and bank vole grew with an increasing forest cover. In agricultural areas, main prey were voles and striped-field mice, while in the suburbs, almost one fourth of consumed biomass were young domestic cats
Felis catus [
75]. In the Białowieża primeval forest, a big forest area, a primeval habitat for common buzzard, Soricomopha accounted for 22.5% of prey items, and bank vole (15% prey items) was the rodent species most commonly caught [
35]. In areas, where voles occur in low densities (North Spain), buzzards preyed mainly on rabbits and reptiles [
76]. In North Ireland, where voles were absent, main prey were rabbits
Oryctolagus cuniculus, rats
Rattus norvegicus, and corvids [
77]. Rabbits may be the primary prey for common buzzards [
14,
78], and their high abundance can be responsible for the buzzard population growth [
9]. In our study area, a sharp drop in the abundance of small game living in agrocenoses (i.e., brown hare, grey partridge, pheasant) was reported recently, while European rabbits went extinct [
38]. However, these species were never an important prey for common buzzard in our study area, and their share in a diet dropped in the last decades. Similarly, a decrease in the share of domestic pigeons and poultry was recorded, which reflected changes in their availability due to socioeconomic changes in Polish farmland [
38]. The other prey that were caught less often now were amphibians. This can also be connected to climate warming and the disappearance of most (known to us) small water bodies being the place for breeding.
4.3. Breeding Parameters and Nesting Tree Choice
Our results showed that in the last decades, the breeding success of the common buzzard did not change and was around 75%. In the 1980s, productivity per breeding pair and per successful pair was slightly higher, yet a dynamic growth of the common buzzard population was observed in the last decades. This slight drop in productivity may be density-dependent [
79] or can be the result of changes in prey availability [
6,
80], as the first study period (1982–1992) was the only one when abundance peaks of voles were observed. Indeed, in Poland, the highest breeding parameters (1.7 juv./breeding pair, 2.3 juv./per successful pair, breeding success 72%) were shown in the 1980s in the agrocenoses of the Western part of the country, where cyclic peaks in the abundance of common voles were reported [
12]. To compare, in the Białowieża National Park (vast, pristine forest), productivity was lower, i.e., 1.3 juv./breeding pair [
69]. Even lower breeding parameters were shown for a continuous forest complex in South-Eastern Poland, i.e., 0.8 juv./per breeding pair and 1.6 juv./successful pair [
81]. A lather low breeding success was also reported in the Italian pre-Alps in the 1990s (breeding success 72%, 1.1 juveniles per breeding pair, 1.5 juveniles per successful pair) [
10].
We also showed that common buzzards adapted to changes in stand structure, i.e., when the share of Scots pine decreased, it was chosen as a nesting tree less frequently. A higher proportion of larch in stands could result in a higher availability of convenient nesting trees. This species grows fast and was used for nesting much earlier than pine, so with a felling age similar to Scots pine, larch stands were available for nesting for a longer period. Moreover, we observed that nests built on larches were much more durable than on other tree species. In Poland, Scots pine was often a dominant nesting tree [
82,
83,
84,
85], as it is the main species in the structure of Polish forests [
86]. By contrast, in the Białowieża National Park, a small-leaved lime
Tilia cordata dominating in stands, was also chosen most often (60% of nests), with Norway spruce
Picea abies and Norway maple
Acer platanoides taking the next places [
69]. In various areas, different tree species can be chosen as nesting places. In Estonia, buzzards placed their nests on Norway spruces (42.3%) and on birch trees
Betula spp. (27.4%) most often [
87]. Norway spruce was also a dominant nesting tree in Finland [
88]. In the pre-Alps area in Italy, birds built their nests on sweet chestnut
Castanea sativa (tree species dominating in this area) [
10,
15], but they may also place their nests on rock stony ledges [
10,
15,
89]. In the agrocenoses of Western Poland, buzzards nested in parks, roadside tree alleys, or even on single trees in the fields [
12]. Such an expansion outside forest complexes was also observed in the last years in our study area. Finally, a case of common buzzard nesting in a building was reported in Spain [
90]. This shows that common buzzards are very adaptable when breeding site choice is concerned.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




