Vacant Bivalve Boreholes Increase Invertebrate Species Richness in a Physically Harsh, Low Intertidal Platform


Abstract
1. Introduction
2. Materials and Methods
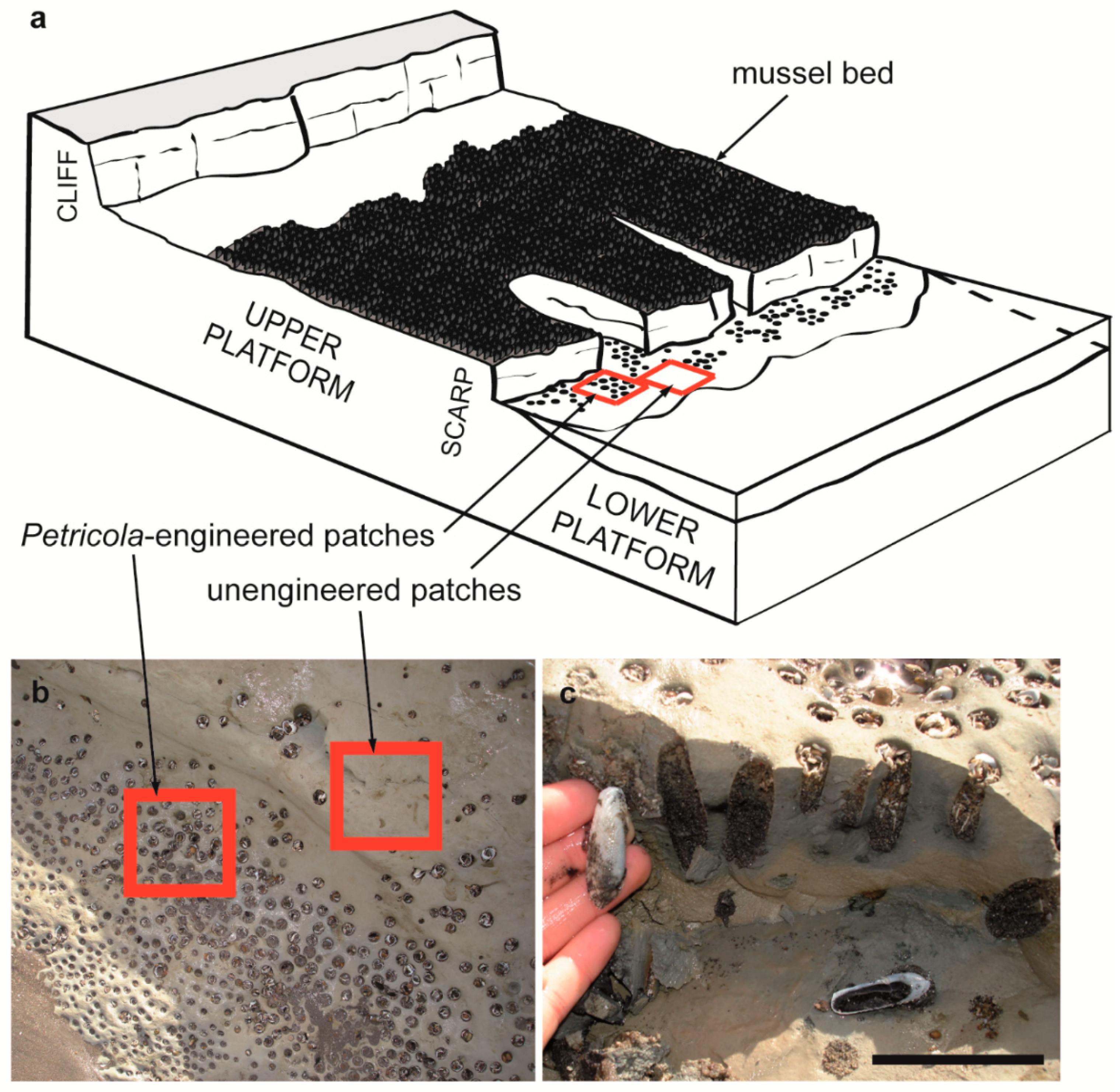
2.1. Study Site and Organisms
2.2. Sampling
2.3. Data Analysis
3. Results
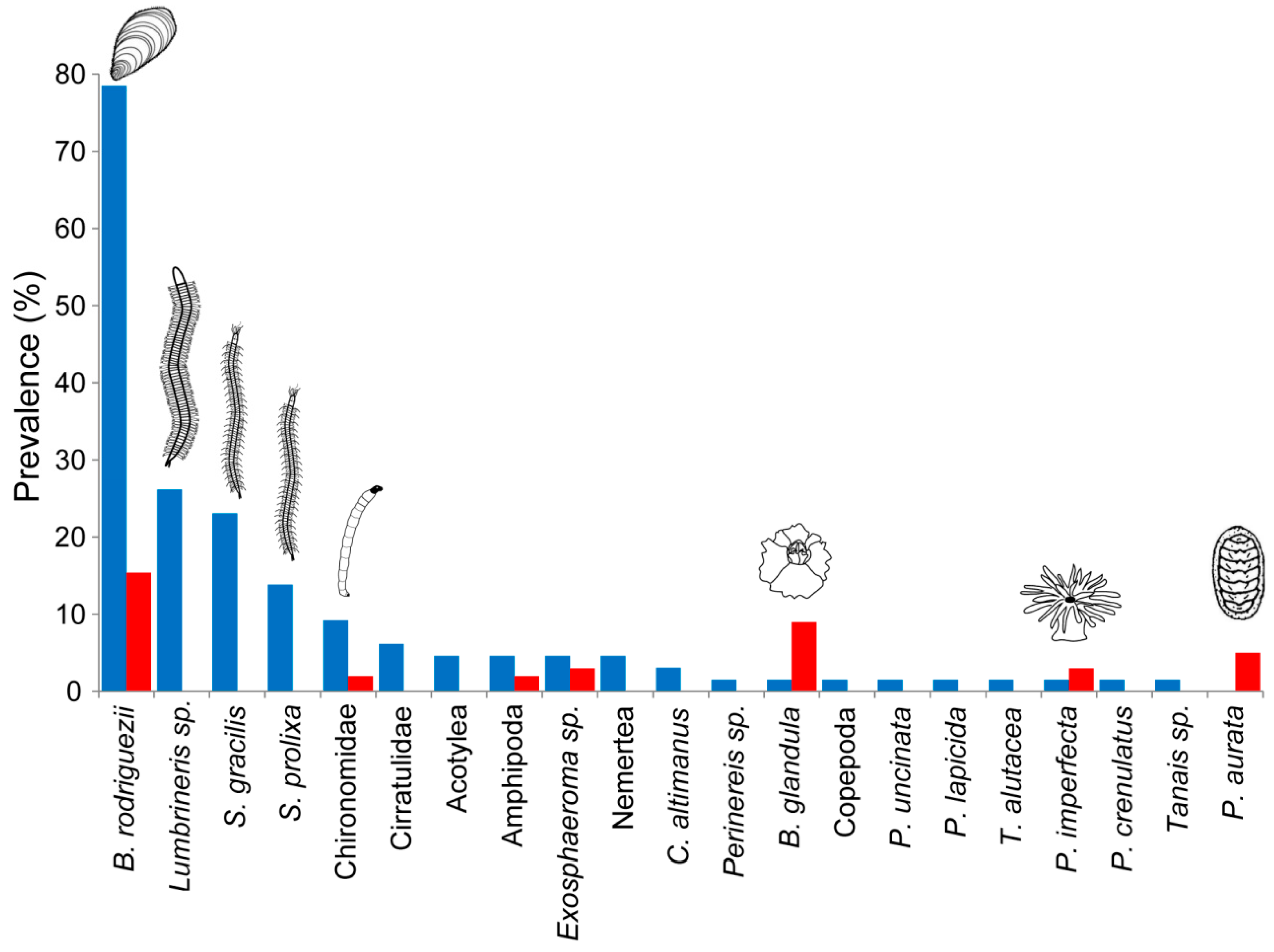
3.1. Invertebrate Species Composition and Richness in SAMPLEs from Petricola-Engineered and Unengineered Patches
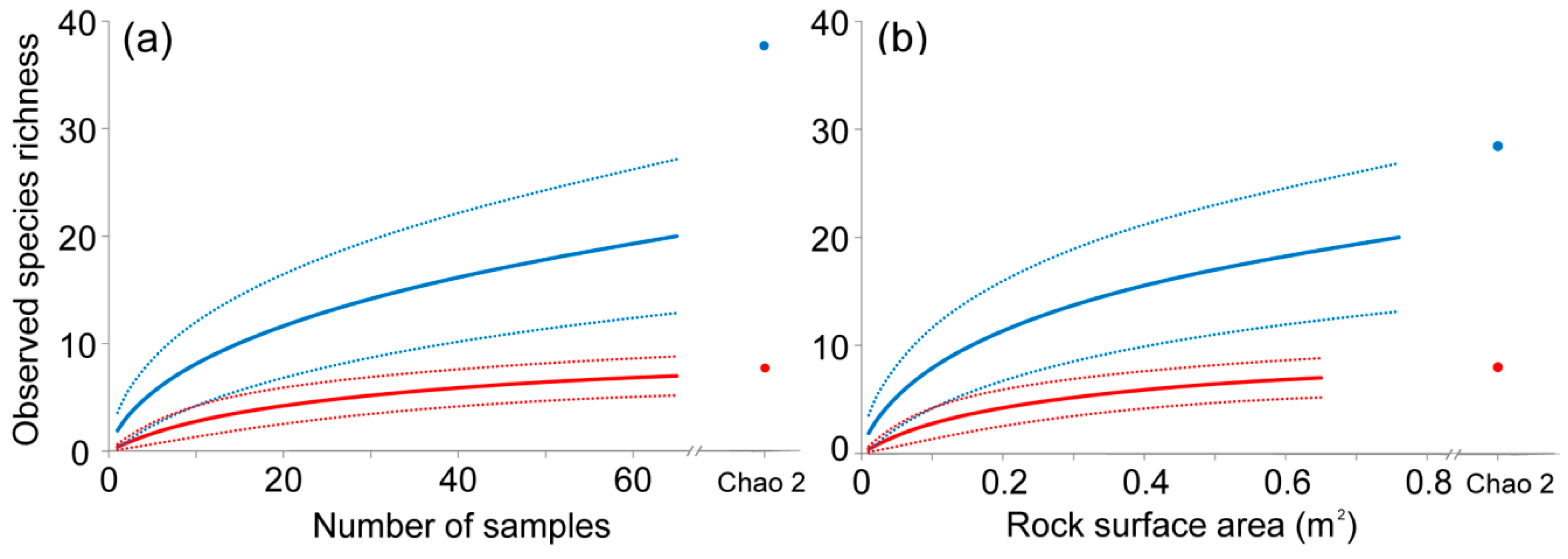
3.2. Invertebrate Species Richness as a Function of Sampled Area
3.3. Invertebrate Species Composition in Petricola-Engineered Patches and Mussel Beds
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bertness, M.D.; Hacker, S.D. Physical stress and positive associations among marsh plants. Am. Nat. 1994, 144, 363–372. [Google Scholar] [CrossRef]
- Crain, C.M.; Bertness, M.D. Ecosystem engineering across environmental gradients: Implications for conservation and management. BioScience 2006, 56, 211–218. [Google Scholar] [CrossRef]
- Badano, E.; Jones, C.; Cavieres, L.; Wright, J. Assessing impacts of ecosystem engineers on community organization: A general approach illustrated by effects of a high-Andean cushion plant. Oikos 2006, 115, 369–385. [Google Scholar] [CrossRef]
- Silliman, B.R.; Bertness, M.D.; Altieri, A.H.; Griffin, J.N.; Bazterrica, M.C.; Hidalgo, F.J.; Crain, C.M.; Reyna, M.V. Whole-community facilitation regulates biodiversity on Patagonian rocky shores. PLoS ONE 2011, 6, e24502. [Google Scholar] [CrossRef] [PubMed]
- Bagur, M.; Gutiérrez, J.L.; Arribas, L.P.; Palomo, M.G. Complementary influences of co-occurring physical ecosystem engineers on species richness: Insights from a Patagonian rocky shore. Biodivers. Conserv. 2016, 25, 2787–2802. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 1997, 78, 1946–1957. [Google Scholar] [CrossRef]
- Gálvez Bravo, L.; Belliure, J.; Rebollo, S. European rabbits as ecosystem engineers: Warrens increase lizard density and diversity. Biodivers. Conserv. 2009, 18, 869–885. [Google Scholar] [CrossRef]
- Bertness, M.D.; Leonard, G.H.; Levine, J.M.; Schmidt, P.R.; Ingraham, A.O. Testing the relative contribution of positive and negative interactions in rocky intertidal communities. Ecology 1999, 80, 2711. [Google Scholar] [CrossRef]
- Castilla, J.; Lagos, N.; Cerda, M. Marine ecosystem engineering by the alien ascidian Pyura praeputialis on a mid-intertidal rocky shore. Mar. Ecol. Prog. Ser. 2004, 268, 119–130. [Google Scholar] [CrossRef]
- Borthagaray, A.I.; Carranza, A. Mussels as ecosystem engineers: Their contribution to species richness in a rocky littoral community. Acta Oecol. 2007, 31, 243–250. [Google Scholar] [CrossRef]
- Foster, B.A. Desiccation as a factor in the intertidal zonation of barnacles. Mar. Biol. 1971, 8, 12–29. [Google Scholar] [CrossRef]
- Garrity, S.D. Some adaptations of gastropods to physical stress on a tropical rocky shore. Ecology 1984, 65, 559–574. [Google Scholar] [CrossRef]
- Harley, C.D.G.; Helmuth, B.S.T. Local- and regional-scale effects of wave exposure, thermal stress, and absolute versus effective shore level on patterns of intertidal zonation. Limnol. Oceanogr. 2003, 48, 1498–1508. [Google Scholar] [CrossRef]
- Teagle, H.; Hawkins, S.J.; Moore, P.J.; Smale, D.A. The role of kelp species as biogenic habitat formers in coastal marine ecosystems. J. Exp. Mar. Biol. Ecol. 2017, 492, 81–98. [Google Scholar] [CrossRef]
- Bertness, M.D.; Crain, C.M.; Silliman, B.R.; Bazterrica, M.C.; Reyna, M.V.; Hildago, F.; Farina, J.K. The community structure of Western Atlantic Patagonian rocky shores. Ecol. Monogr. 2006, 76, 439–460. [Google Scholar] [CrossRef]
- Connell, J.H. The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalus stellatus. Ecology 1961, 42, 710–723. [Google Scholar] [CrossRef]
- Connell, J.H. Community interactions on marine rocky intertidal shores. Annu. Rev. Ecol. Syst. 1972, 3, 169–192. [Google Scholar] [CrossRef]
- Dayton, P.K. Competition, disturbance, and community organization: The provision and subsequent utilization of space in a rocky intertidal community. Ecol. Monogr. 1971, 41, 351–389. [Google Scholar] [CrossRef]
- Bertness, M.D.; Leonard, G.H. The role of positive interactions in communities: Lessons from intertidal habitats. Ecology 1997, 78, 1976. [Google Scholar] [CrossRef]
- Coan, E.V. Recent species of the genus Petricola in the eastern Pacific (Bivalvia: Veneroidea). Veliger 1997, 40, 298–340. [Google Scholar]
- Bagur, M.; Gutiérrez, J.L.; Arribas, L.P.; Palomo, M.G. Endolithic invertebrate communities and bioerosion rates in southwestern Atlantic intertidal consolidated sediments. Mar. Biol. 2014, 161, 2279–2292. [Google Scholar] [CrossRef]
- Scarabino, F.; Zelaya, D.G.; Orensanz, J.M.; Ortega, L.; Defeo, O.; Schwindt, E.; Carranza, A.; Zaffaroni, J.C.; Martínez, G.; Scarabino, V. Cold, warm, temperate and brackish: Bivalve biodiversity in a complex oceanographic scenario (Uruguay, southwestern Atlantic). Am. Malacol. Bull. 2015, 33, 284–301. [Google Scholar] [CrossRef]
- Sunamura, T. Rocky coast processes: With special reference to the recession of soft rock cliffs. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2015, 91, 481–500. [Google Scholar] [CrossRef] [PubMed]
- Zavala, C.; Freije, H. Geología de los acantilados. In Las Mesetas Patagónicas que Caen al Mar: La Costa Rionegrina; Massera, R.F., Lew, J., Serra Pairano, G., Eds.; Gobierno de Río Negro: Viedma, Argentina, 2005; pp. 187–199. ISBN 987-22604-0-0. [Google Scholar]
- Andreis, R.R. Petrografía y Paleocorrientes de la Formación Río Negro (Tramo General Conesa—Boca del Río Negro). Ph.D. Thesis, Facultad de Ciencias Naturales y Museo, La Plata, Argentina, 1965; pp. 245–310. [Google Scholar]
- Arribas, L.P.; Gutiérrez, J.L.; Bagur, M.; Soria, S.A.; Penchazadeh, P.E.; Palomo, M.G. Variations in aggregate descriptors of rocky shore communities: A test of synchrony across spatial scales. Mar. Biol. 2019, 166, 44. [Google Scholar] [CrossRef]
- Arribas, L.P.; Bagur, M.; Klein, E.; Penchaszadeh, P.E.; Palomo, M.G. Geographic distribution of two mussel species and associated assemblages along the northern Argentinean coast. Aquat. Biol. 2013, 18, 91–103. [Google Scholar] [CrossRef]
- Arribas, L.P.; Bagur, M.; Gutiérrez, J.L.; Palomo, M.G. Matching spatial scales of variation in mussel recruitment and adult densities across southwestern Atlantic rocky shores. J. Sea Res. 2015, 95, 16–21. [Google Scholar] [CrossRef]
- Trovant, B.; Orensanz, J.L.; Ruzzante, D.E.; Stotz, W.; Basso, N.G. Scorched mussels (Bivalvia: Mytilidae: Brachidontinae) from the temperate coasts of South America: Phylogenetic relationships, trans-Pacific connections and the footprints of Quaternary glaciations. Mol. Phylogenet. Evol. 2015, 82, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Trenhaile, A.S.; Kanyaya, J.I. The role of wave erosion on sloping and horizontal shore platforms in macro-and mesotidal environments. J. Coast. Res. 2007, 298–309. [Google Scholar] [CrossRef]
- Dickson, M.E.; Pentney, R. Micro-seismic measurements of cliff motion under wave impact and implications for the development of near-horizontal shore platforms. Geomorphology 2012, 151, 27–38. [Google Scholar] [CrossRef]
- Paine, R.T. Food web complexity and species diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- Breitburg, D.L. Residual effects of grazing: Inhibition of competitor recruitment by encrusting coralline algae. Ecology 1984, 65, 1136–1143. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Wiens, J.A. Spatial scaling in ecology. Funct. Ecol. 1989, 3, 385–397. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9. 2013. Available online: http://purl.oclc.org/estimates (accessed on 30 January 2019).
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999; pp. 128–129. ISBN 0-13-081542-x. [Google Scholar]
- Chao, A. Non–parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Almeida-Neto, M.; Guimarães, P.; Guimarães, P.R., Jr.; Loyola, R.D.; Ulrich, W. A consistent metric for nestedness analysis in ecological systems: Reconciling concept and measurement. Oikos 2008, 117, 1227–1239. [Google Scholar] [CrossRef]
- Guimarães, P.R., Jr.; Guimarães, P. Improving the analyses of nestedness for large sets of matrices. Environ. Model. Softw. 2006, 21, 1512–1513. [Google Scholar] [CrossRef]
- Menge, B.A.; Lubchenco, J. Community organization in temperate and tropical rocky intertidal habitats: Prey refuges in relation to consumer pressure gradients. Ecol. Monogr. 1981, 51, 429–450. [Google Scholar] [CrossRef]
- Pinn, E.H.; Thompson, R.C.; Hawkins, S.J. Piddocks (Mollusca: Bivalvia: Pholadidae) increase topographical complexity and species diversity in the intertidal. Mar. Ecol. Prog. Ser. 2008, 355, 173–182. [Google Scholar] [CrossRef]
- Hendy, I.W.; Michie, L.; Taylor, B.W. Habitat creation and biodiversity maintenance in mangrove forests: Teredinid bivalves as ecosystem engineers. PeerJ 2014, 2, e591. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.W. The role of Penitella penita (Conrad 1837) (Family Pholadidae) as eroders along the Pacific coast of North America. Ecology 1968, 49, 156–159. [Google Scholar] [CrossRef]
- Hutchings, P.A. Biological destruction of coral reefs, a review. Coral Reefs 1986, 4, 239–252. [Google Scholar] [CrossRef]
- Horonjeff, R.; Patrick, D. Action of marine borers and protective measures against attack. Coast. Eng. Proc. 1951, 1, 8. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Taxa 1 | Adhesion 2 | Unengineered | Petricola-Engineered | Mussel-Engineered |
|---|---|---|---|---|---|
| Parabunodactys imperfecta | CnA | YES | X | X | |
| Acotylea indet. | P | NO | X | ||
| Lumbrineris sp. | AP | NO | X | X | |
| Perinereis sp. | AP | NO | X | X | |
| Syllis gracilis | AP | NO | X | X | |
| Syllis prolixa | AP | NO | X | X | |
| Cirratulidae | AP | NO | X | ||
| Protoarcinella uncinata | AP | NO | X | X | |
| Platynereis sp. | AP | NO | X | ||
| Boccardia polybranchia | AP | NO | X | ||
| Capitella capitata? | AP | NO | X | ||
| Nereididae (Allita?) | AP | NO | X | ||
| Brachidontes rodriguezii | MB | YES | X | X | X |
| Petricola lapicida | MB | NO | X | ||
| Mytilus edulis | MB | YES | X | ||
| Plaxiphora aurata | MP | YES | X | ||
| Siphonaria lessoni | MG | YES | X | ||
| Themiste alutacea | S | NO | X | ||
| Nemertea indet. | N | NO | X | X | |
| Chironomidae indet. | ID | YES | X | X | X |
| Balanus glandula | CrCi | YES | X | X | X |
| Copepoda indet. | CrCo | NO | X | ||
| Hyale grandicornis | CrA | NO | X | ||
| Melitidae indet. | CrA | NO | X | ||
| Ampithoidae indet. | CrA | NO | X | ||
| Corophiidae indet. | CrA | NO | X | X | X |
| Exosphaeroma sp. | CrI | NO | X | X | X |
| Tanais sp. | CrT | NO | X | X | |
| Cyrtograpsus altimanus | CrD | NO | X | X | |
| Platyxanthus crenulatus | CrD | NO | X | ||
| Total | 7 | 20 | 22 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagur, M.; Gutiérrez, J.L.; Arribas, L.P.; Palomo, M.G. Vacant Bivalve Boreholes Increase Invertebrate Species Richness in a Physically Harsh, Low Intertidal Platform. Diversity 2019, 11, 39. https://doi.org/10.3390/d11030039
Bagur M, Gutiérrez JL, Arribas LP, Palomo MG. Vacant Bivalve Boreholes Increase Invertebrate Species Richness in a Physically Harsh, Low Intertidal Platform. Diversity. 2019; 11(3):39. https://doi.org/10.3390/d11030039
Chicago/Turabian StyleBagur, María, Jorge L. Gutiérrez, Lorena P. Arribas, and M. Gabriela Palomo. 2019. "Vacant Bivalve Boreholes Increase Invertebrate Species Richness in a Physically Harsh, Low Intertidal Platform" Diversity 11, no. 3: 39. https://doi.org/10.3390/d11030039
APA StyleBagur, M., Gutiérrez, J. L., Arribas, L. P., & Palomo, M. G. (2019). Vacant Bivalve Boreholes Increase Invertebrate Species Richness in a Physically Harsh, Low Intertidal Platform. Diversity, 11(3), 39. https://doi.org/10.3390/d11030039