1. Introduction
The knowledge of thermophiles biodiversity could be very useful for the discovery of new molecules and/or more thermostable biocatalysts with thermophilic and thermostability properties superior to those currently known and in use [
1,
2]. Recent exploitations have clearly shown that thermophilic microorganisms are industrial providers of novel catalysts, and in some cases, these enzymes have been cloned and expressed in suitable mesophilic hosts [
1,
3].
The biggest part of thermophiles studied belongs to the genus
Geobacillus, and several strains have been found in thermophilic and mesophilic environments [
1,
4]. The development of extremophiles from geothermal areas, as a resource for novel thermostable enzymes, has received attention even in Antarctica, where the presence of thermophiles has been indicated by the discovery of several geothermal areas [
5]. Thermophilic enzymes (also called thermozymes) work and remain stable at high temperatures, and they are to be preferred to the mesophilic counterpart for their biotechnological potential [
6].
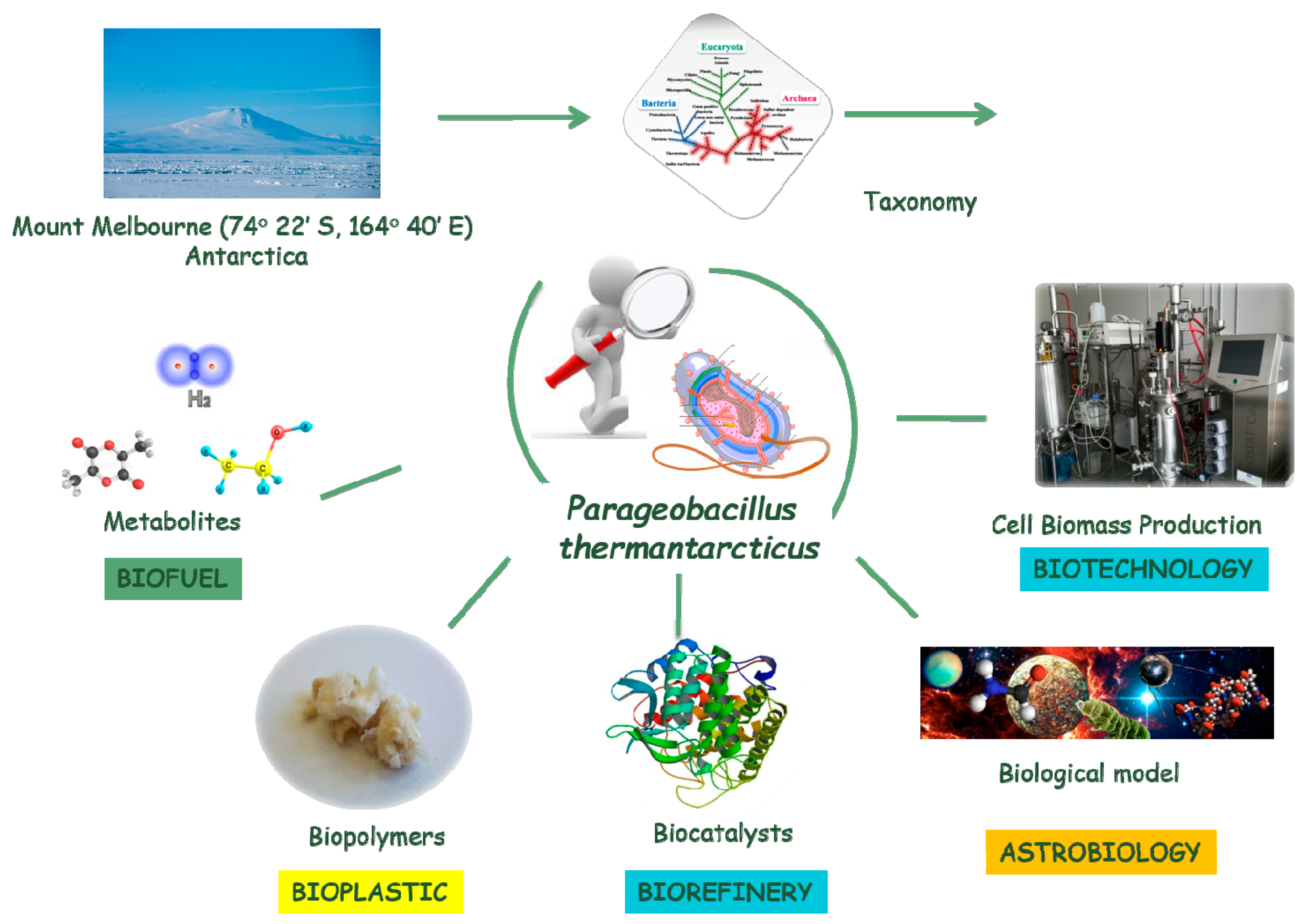
In view of this, a thermophilic bacterium,
Parageobacillus thermantarcticus, (DSM 9572
T) strain M1 [
7,
8], isolated from geothermal soil of Mount Melbourne (74°22′ S, 164°40′ E) in Antarctica (Italian Antarctica Expedition during the Austral summer 1986–1987), was characterized for the production of thermostable enzymatic activities such as an extracellular xylanase, a beta-xylosidase, an intracellular xylose/glucose isomerase, and a protease.
The interesting enzymatic pathways present in
P. thermantarcticus make this Antarctic thermophile of great potential in biotechnological applications, not only for industrial biocatalysis but also for bioprocessing aims. In fact, the optimization of enzymatic procedures for the conversion of waste lignocellulosic materials in high added value molecules using the xylanolytic enzymatic set of
P. thermantarcticus produced extremely interesting results and was recently reported [
9,
10]. These enzymes, which were involved in the enzymatic digestions of hemicellulolitic fraction obtained from rhizomes of
Arundo donax and the waste stems and leaves biomasses of
Cynara cardunculus, have been used for the bioconversion of agroresidues into xylo-oligomers and fermentable sugars.
Moreover, it was demonstrated that
P. thermantarcticus produces two exopolysaccharides (called EPS 1 and EPS 2) that are responsible for typical mucous colonies. These EPSs were produced using mannose as a sole carbon and energy source with a yield of 400 mg/L. Nuclear magnetic resonance spectra established that EPS 1 was a xantan polymer, while EPS 2 was a mannan polysaccharide [
11].
P. thermantarcticus has also been used as a biological model for astrobiology, i.e., the multidisciplinary approach to the study of origin and evolution of life on Earth and in the Universe. It has been demonstrated that either the viable cells or the spores of
P. thermantarcticus to space simulation are interesting for the issue of the origin of life [
12,
13,
14]. Indeed, the finding of bacterial species fit to survive in space parameters open new scenarios for the so-called panspermia theory. According to this theory, life on Earth could have originated from bacterial species from other places transported by solar pressure. The identification of bacteria like
P. thermantarcticus that potentially could survive to the space transport is one main key point to support this theory.
Recently, thanks to participation to “The Genomic Encyclopedia of Bacteria and Archaea (GEBA) III Project”, a project of the Community Science Program (CSP) of the DOE Joint Genomes Institute, the draft genomic sequence of
P. thermantarcticus using the Illumina technology has been obtained. With the available genome sequence, the biotechnology potential and the use of
P. thermantarcticus will increase. For example, like a factory,
P. thermantarcticus could convert unexplored feedstocks into useful products such as biobased chemicals, from renewable resources, for biofuels production, bioplastic materials development, etc. The increase of genetic tools is a requirement for high-throughput engineering. There is the opportunity to explore and exploit the physiology of a really interesting bacterium like
P. thermantarcticus: This will necessarily involve an integrated approach of various disciplines (
Figure 1) [
15].
2. Phylogenomic Re-Assessment of Bacillus thermoantarcticus to Parageobacillus thermantarcticus
It was 1996 when Nicolaus et al. [
8] described the isolation and the taxonomic description of a new Antarctic thermophilic species of
Bacillus strain M1, isolated from the geothermal soil of Mount Melbourne, for which the name
Bacillus thermoantarcticus was indicated. The name
thermoantarcticus was subsequently corrected in
thermantarcticus (therm.ant.arc’ti.cus. Gr. n. thermệ heat; L. masc. adj.
antarcticus southern, belonging to Antarctica; N.L. masc. adj.
thermantarcticus, a thermophile from Antarctica) in the Validation List N. 84 published in IJSEM [
16]. Nicolaus et al. [
8] described strain M1 as able to utilize some sugars such as glucose, trehalose, and xylose as sole carbon sources and overall able to synthesize an exopolysaccharide and some very interesting enzymes: an exocellular xylanase, exo-and endocellular alpha-glucosidase(s) and an intracellular alcohol dehydrogenase. Already at the time of its first description, the M1 strain showed remarkable biotechnological potential thanks to its enzymatic pattern and biopolymer synthesis. The 16S rRNA gene sequence analysis placed
Bacillus thermantarcticus as a peripheral member of the thermophilic
Bacillus rRNA group 5 [
7,
17].
Bacillus rRNA group 5 comprised thermophilic bacilli with 16S rRNA gene sequences similarity ranging between 98 ± 5% and 99 ± 2% and comprised
B. thermoglucosidiasius,
B. stearothermophilus,
B. kaustophilus,
B. thermocatenulatus,
B. thermoleovorans,
B. thermodenitrificans, and
B. thermantarcticus, some isolates not yet validly published and
Saccharococcus thermophiles [
18,
19]. A few years later, in 2002, Nicolaus et al. [
7] reported that “although the genus
Bacillus is the largest and the best-known member of the family
Bacillaceae, its taxonomy is very confusing, and new species often have been described on the basis of only a few physiological or ecological features”, underlining the necessity of applying a polyphasic approach, including phenotypic, chemical, and genotypic characterization, in order to define a new taxonomic position of endospore forming thermophiles.
Nazina et al. [
20] isolated five hydrocarbon-oxidizing strains from formation waters of oilfields in China, Russia, and Kazakhstan and compared their 16S rRNA gene sequences with those available for other members of the
Bacillaceae. Strains were further studied for their physiological and biochemical aspects, including analysis of cell membrane fatty acid composition and DNA–DNA hybridization analysis with the most closely related
Bacillus species. Nazina et al. [
20], based on their studies, proposed creating a new genus,
Geobacillus gen. nov., including two new species denominated as
Geobacillus subterraneus sp. nov. and
Geobacillus uzenensis sp. nov., and proposed the shift of the validly described species of group 5,
B. kaustophilus,
B. thermoglucosidasius,
B. stearothermophilus,
B. thermoleovorans,
B. thermocatenulatus, and
B. thermodenitrificans, to this new genus, with
Geobacillus stearothermophilus (formerly
Bacillus stearothermophilus DSM 22
T) as the type species of the genus. Moreover, the authors suggested an in-depth study and analysis to establish the taxonomic position of the not yet validated species as ‘
B. caldotenax’, ‘
B. caldovelox’, ‘
B. caldolyticus’, ‘
B. caldoxylolyticus’, ‘
B. thermoantarcticus’, and
Saccharococcus thermophilus, present in group 5.
Subsequently,
Bacillus pallidus [
21],
Saccharococcus caldoxylosilyticus [
22], and
Bacillus vulcani [
23] were transferred to
Geobacillus [
24,
25,
26], and then Miñana-Galbis et al. [
27] suggested the further transfer of
Geobacillus pallidus to the new genus
Aeribacillus. Moreover, the following new species
Geobacillus toebii [
28],
Geobacillus gargensis [
26],
Geobacillus debilis [
25],
Geobacillus lituanicus [
29],
Geobacillus tepidamans [
30], and
Geobacillus jurassicus [
31] were proposed. Coorevitis et al. [
32] performed a modern polyphasic study to clarify the taxonomy of some members of the
Geobacillus genus, such as
Bacillus thermantarcticus, also including
G. stearothermophilus—the type of species of the genus
Geobacillus—whose taxonomic description was mainly provided by Claus and Berkeley [
33]. Even Zeigler in 2005 [
34] had suggested that
B. thermantarcticus should be transferred to the genus
Geobacillus, relying on recN and 16S rRNA gene sequences. Coorevitis et al. [
32] demonstrated that
B. thermantarcticus, from a phenotypic and phylogenetic point of view, was distant from the type strain of
G. thermoglucosidasius (now
G. thermoglucosidans), suggesting the transfer to the genus
Geobacillus as the separate species
Geobacillus thermantarcticus comb. nov.
Aliyu et al. [
35], in subsequent experiments, using the phylogenomic metrics AAI (average aminoacid identity), ANI (average nucleotide identity), and dDDH (digital DNA–DNA hybridization) applied to 63 genome sequences of
Geobacillus strains, came to the conclusion that the genus
Geobacillus comprised sixteen distinct genomospecies and, among these, also different and potential new species. Moreover, the authors stated that the genus
Geobacillus clusters into two monophyletic clades with different nucleotide base compositions: the clade I, containing a G + C content range of 48.8–53.1%, and the clade II, in which the G + C content range is 42.1–44.4%. The authors, therefore, suggested that all
Geobacillus species that fall in the clade II can be clustered into a new genus and created a new genus,
Parageobacillus (Pa.ra.ge.o.ba.cil’lus. Gr. prep. Para, beside or alongside of; n. Gê the Earth; L. dim. n. bacillus small rod; M.L. masc. n.
Parageobacillus, a genus nearest to
Geobacillus). The
Parageobacillus genus included five “genomospecies”,
P. toebii,
P. caldoxylosilyticus,
P. thermantarcticus,
P. thermoglucosidasius, and
Parageobacillus genomospecies 1 (strain NUB3621). The current position of
P. thermantarcticus is shown in
Figure 2, in which the Antarctic bacterium was analyzed using the EzTaxon-e server. The phylogenetic tree was reconstructed using the software MEGA version 5 after multiple alignments of the data by CLUSTAL_X. Distances (distance options according to Kimura’s two-parameter model) and clustering were based on the neighbor-joining method [
32].
4. Production of Xylo-Oligosaccharides as High Value-Added Molecules
β-1,4-xylan is the second most widespread polymer present in nature, representing one-third of the available renewable biomass [
10,
87]. Using proper chemical, autohydrolytic, or enzymatic procedures, it is possible to produce (XOS), D-xylose and, to a lesser extent, L-arabinose from crop waste residues and forestry biomass [
88,
89,
90].
The industrial interest for the xylo-oligosaccharides is increasing because of their nutraceutical properties. These molecules can work as selective prebiotics for the gut microflora, reduce the blood glucose and cholesterol, reduce the procarcinogenic enzymes in the gastrointestinal tract, and enhance mineral absorption from the large intestine. Xylobiose has been shown to have a stimulatory effect on the selective growth of human intestinal
Bifidobacteria, which are extremely useful for the maintenance of a healthy intestinal microflora. In addition, XOS, which can be used in anti-obesity diets, are noncariogenic and low-calorie [
90,
91,
92,
93,
94,
95]. These beneficial effects are caused by the chemical structure of the xylo-oligosaccharides, which in turn are affected by the adopted hemicellulose extraction procedures.
In this sense, xylanases and β-xylosidases enzymes isolated from thermophilic strains of bacteria and fungi can be considered essential from a biotechnological viewpoint, due to their higher thermostability, and other biochemical properties, which are fundamental for industrial applications [
96,
97].
The optimization of enzymatic procedures for the conversion of waste lignocellulosic materials in XOS molecules using the xylanolytic enzymatic set from
Geobacillus microorganisms produced extremely interesting results and was recently reported [
50,
51,
98].
In particular,
P. thermantarcticus produced an extracellular xylanase (1,4-β-D-xylan xylanohydrolase; E.C. 3.2.1.8) and a β-xylosidase (1,4-β-D-xylan xylohydrolase; E.C. 3.2.1.37), which were involved in the enzymatic digestions of hemicellulose extract obtained from rhizomes of
Arundo donax L. and the waste biomass of
Cynara cardunculus stems and leaves [
9,
10]. Among the several biomass crops for lignocellulosic materials, one of the most interesting is giant reed,
A. donax. The most useful and studied part of
A. donax L. is represented by shoots (reeds) of interest in bioethanol, biodiesel or biopolymer production [
9].
However, at the end of the cropping cycle of a giant reed stand (10–15 years), a huge number of rhizomes is left on the soils, and they have to be eliminated to allow further agricultural utilizations. These rhizomes could represent a source of useful molecules for bioenergy or biochemicals [
9,
51].
On the other hand,
C. cardunculus is another interesting crop due to its ability to produce both biomass from the whole plant, and oil from the seeds previously removed from the plants [
10].
For the extraction of the hemicellulosic fraction from both biomasses, a 72-h alkaline treatment was considered to be the most appropriate in comparison with other methods [
99]. In particular, starting from hemicellulose biomass from
A. donax rhizomes, an extracellular suspension of
P. thermantarcticus, which contained either xylanase and beta-xylosidase activities, was able to produce, at a temperature of 70 °C, a mixture of xylose and XOS up to five- and six-monosaccharide units; the oligosaccharides amount emerged prevailing in the case of higher ratios between the xylan substrate and the enzyme amounts (substrate/biocatalyst ratio, S/B), as reported in
Table 4 [
9]. However, the efficiency of these digestion procedures was evaluated as a percentage of the equivalent reducing xylose obtained (dinitrosalicyclic colorimetric assay–DNS–) [
100]; in any case, it was over 60%.
Marcolongo et al. [
51] reported the saccharification by an alkali-thermostable xylanase from
Geobacillus thermodenitrificans A333 of hemicellulose fractions from several agroderived biomasses (brewery’s spent grains, corn cobs, and
A. donax biomass). The polysaccharidic fractions were obtained after alkali treatment; in particular, the digestion of
A. donax hemicellulose produced an end-product profile made of xylopentaose, xylotriose, xylobiose (with a prevalence of X2 upon long time incubation), and similar to that recovered using
P. thermantarcticus xylanase [
9]. By contrast, the evaluation of the reducing sugars revealed that the percentage of
A. donax xylan degradation by
G. thermodenitrificans A333 xylanase, was very low, also reaching 7.17% after 72 h of reaction.
Instead, a yield bioconversion of 62% and 44%, in terms of reducing sugars (production of xylo-oligosaccharides) and xylose releases, respectively (
Table 4), was obtained through the simultaneous use of endoxylanase and β-xylosidase activities of
P. thermantarcticus at a temperature of 70 °C, starting from the hemicellulosic fraction isolated from
C. cardunculus [
10]. MS analysis of the reaction mixture after 24 h suggested the presence of xylose, xylotriose, xylotetraose, and also of two disaccharides: the first made of xylose and
O-methyl glucuronic acid and the second one of xylose and glucuronic acid.
These results were similar to those obtained from the commercial beechwood xylan digestion using the thermostable xylanase from the
Geobacillus sp. strain DUSELR13 [
50]. In a second bioprocess, the xylanase and β-xylosidase activities from
P. thermantarcticus were separated according to (i) a procedure for the xylanase recovery (ES-GTM endo-xyl) based on differences between the thermal stabilities of the two proteins [
55], for which the β-xylosidase activity could be eliminated by heat treatment at 80 °C for 20 min, and (ii) a further procedure for the beta-xylosidase recovery, which exploited the possibility of many xylanases to be absorbed on xylan polymers [
101]. It was possible to obtain a xylan-bound fraction that contained 100% of the inactive xylanase and an unbound fraction that contained 100% of the active β-xylosidase (ES-GTM beta-xyl) from the extracellular enzymatic pool of
P. thermantarcticus [
10].
These enzymatic preparations (ES-GTM endo-xyl, ES-GTM beta-xyl) were then used individually or consecutively in cascade hydrolysis reactions aiming to accumulate xylose or xylo-derivatives selectively into the reaction media, as reported in
Table 4. The polysaccharide bioconversion after the exclusive ES-GTM endo-xyl attack furnished a yield of 43.7% as a percentage of reducing sugars with only 6.8% of the xylose content. In addition, the hydrolysis products after 24 h were essentially composed of xylobiose (26.5%), xylotriose (5.5%), and oligomers (DP > 3), as indicated by the HPAE–PAD (High Performance Anion-Exchange. Chromatography with Pulsed. Amperometric Detection) analyses, being the composition of reaction products related to the absence of β-xylosidase activity. Furthermore, in these experimental conditions, two new tetrasaccharides were produced. Their structures were identified, and chemical shifts were totally assigned by NMR spectroscopy; they corresponded to the 2′-
O-(4-
O-methyl-α-D-glucuronosyl) xylotriose (compound 1, α-D-4OMeGlcA-(1-2)-β-D-Xyl-(1-4)-β-D-Xyl-(1-4)-β-D-Xyl)) and the 2′-
O-α-D-glucuronosyl xylotriose (compound 2, α-D-GlcA-(1-2)-β-D-Xyl-(1-4)-β-D-Xyl-(1-4)-β-D-Xyl), which were produced with yields of 22% and 13%, respectively (
Figure 3).
It is worth emphasizing that a xylotriose substituted by 4-O-methylglucuronic acid located at the non-reducing end, which was structurally similar to compound 2, was later identified by HPAEC analysis of a birchwood xylan depolymerization by the
Geobacillus thermodenitrificans T12 endo-xylanase, GtXynA1 [
48]. Alternatively, an enzymatic cascade reaction was carried out; in fact, the xylo-oligosaccharides mixture produced by the ES-GTM endo-xyl digestion (line 4,
Table 4) was involved in a consecutive hydrolysis step using the isolated β-xylosidase from
P. thermantarcticus (ES-GTM beta-xyl) (line 5,
Table 4) [
10]. In this way, 73% of reducing sugars were produced with a xylose yield of 62.6% in the reaction mixture. Exploiting the different optimal temperatures and thermostabilities of each enzyme, in the enzymatic cascade attack, reactions were carried out at a temperature of 70 °C for the first step (xylanase activity), and at 60 °C for the second step (β-xylosidase activity). In comparison with the digestion procedures involving both hemicellulolytic enzymes (lines 1–4,
Table 4), the enzymatic cascade result was more efficient, with an increase of 10% of reducing sugars and about 20% of xylose.
The combined use of the xylanase and β-xylosidase activities from the extremely thermophilic bacterium
Geobacillus thermodenitrificans TSAA1 was also previously reported [
40]. The endoxylanase was active over a broad range of pH (5.0–10.0) and temperatures (30–100 °C), with optima at pH 7.5 and 70 °C, while β-xylosidase was optimally active at pH 7.0 and 60 °C. In this case, the end products’ Thin Layer Chromatography analysis of hydrolysis of xylan from birchwood and xylan containing agroresidues revealed the presence of xylobiose, xylotriose, xylotetraose, xylopentaose, and other higher xylooligosaccharides by the exclusive endoxylanase action. Upon incubation of an enzyme preparation containing both xylanase and β-xylosidase on xylan containing agroresidues, xylose formation was recorded from the very beginning along with other xylooligosaccharides; on increasing the time of incubation, the xylose band got intensified on TLC.
The synergistic hydrolysis of xylan using a xylanase, β-xylosidases, and an α-L-arabinofuranosidase from
Geobacillus thermodenitrificans NG80-2 was described by Huang et al. [
102]. The reducing sugars released from birchwood, oat spelt, and beechwood xylan hydrolysis in the presence of xylanase, β-xylosidase, α-L-arabinofuranosidase, or their enzyme combinations suggested that a synergistic effect on each enzymatic activity on hemicellulose degradation was recorded, and the degree of synergy depended on the substrate. However, in any case, yields lower than 60% in terms of reducing sugars and for processes with a degree of enzymatic synergy of 1.55 were recorded.
Xylooligosaccharides (XOS) and arabinoxylooligosaccharides (AXOS) were produced from the insoluble arabinoxylan fraction of pretreated wheat bran by endoxylanases GsXyn10A from
Geobacillus stearothermophilus [
98]. The AXOS obtained by xylanase treatment of pretreated wheat bran were analyzed by HPAEC-PAD: The major arabinose-substituted products identified with
GsXyn10A were A
3X, minor amounts of A
2XX. Their structure was confirmed after enzymatic treatment of AXOS with specific arabinofuranosidases. The prebiotic effect of XOS, AXOS, and mixtures of XOS and AXOS has been reported by Broekaert et al. [
103]. In conclusion, the results reported for
P. thermantarcticus encouraged the application of these enzymatic separation procedures on the industrial scale. An efficient enzymatic employment in the bioconversion of agroresidues for obtaining both xylo-oligomers and fermentable sugars was here described. Moreover,
P. thermantarcticus β-xylosidase, showing a scarce substrate inhibition in agreement with some of the recently discovered β-xylosidase behavior [
104], could represent a useful enzyme for industrial purposes.
5. Relevance of P. thermantarcticus to Astrobiology
P. thermantarcticus has been used as a biological model for Astrobiology, i.e., the multidisciplinary approach to the study of origin and evolution of life on Earth and in the Universe. The study of extremophiles is particularly relevant for the search of life and for the issue of habitability in the universe—indeed, as also envisaged by the recent European Astrobiology Roadmap [
105]. Microbiology experiments, carried out either in space or in laboratories by simulating space conditions, are one main tool for the definition of boundaries of life on Earth or on other planetary bodies, i.e., the so-called exoplanets (Mars, Europa, Enceladus, etcetera).
On the basis of these considerations, we exposed cells of
P. thermantarcticus to the extreme environmental parameters that are typical of space and exoplanets [
12,
13]. Different laboratory simulations mimicking the extraterrestrial conditions were carried out as schematically reported in
Table 5. For all the conditions assayed, the resistance of
P. thermantarcticus to the space environment’s simulation was assessed by measuring the ability to restart cellular growth after the stressing treatments. The simulations performed included: the extreme temperature variations, by storing cells for 3 months at temperatures ranging from −196 °C to 80 °C; the absence of water, by storing desiccated cells at −20 °C or at room temperature for time intervals up to 1 year; exposition to UV rays at the bactericidal λ 254 nm, with doses ranging from 125 J/m
2 up to 1000 J/m
2; finally, the exposure to X-rays, with radiation doses from 100 Gy up to 1000 Gy. In all cases, cells of
P. thermantarcticus were able to grow as much as the not stressed cells. As shown in
Table 5, only in the case of desiccation and UV-rays exposure, there was a slight delay in the growth’s kinetics observed, while for other stressors, no appreciable difference was found with respect to the not stressed samples.
Moreover, in order to investigate the biological effectiveness of the space simulation, the analysis of some key cellular structures was performed [
13]. As reported in
Table 5, membranes, proteins, and DNA were the cellular components that were investigated after exposure to the stressing conditions mimicking the space environment. With regard to membranes, in all the simulations,
P. thermantarcticus adapted its membranes by increasing its fluidity, as suggested by the increase of the branched fatty acids; on the other hand, a partial decrease in the amount of the demethylated form of menaquinone was observed, except for the UV ray exposure. The effect of ultraviolet radiations was more marked for the DNA, since UV-rays induce the generation of different kinds of photo products [
106]; other stressors did not induce a significant modification in the nucleic acid. Finally, the protein component of this bacterium also did not undergo a significant modification to the space simulation.
P. thermantarcticus is a sporulating species, i.e., under unfavorable conditions, like the absence of water or nutrients, excess of pressure, temperature or radiations, its cells undergo conversion into metabolically dormant spores. The spores of different bacterial species, especially those of bacteria from the related
Bacillus genus, were also shown to be very resistant for a million years, and for this reason, they have been chosen as a biological model for astrobiology [
107]. The investigation of bacterial spores’ resistance to ionizing radiations, which represent a major part of the cosmic radiation spectrum, is one of the main issues for the study of habitability. Therefore, further studies were performed using exposure of the spores of
P. thermantarcticus to irradiation with UV and γ-rays [
14]. In both cases, resistance to irradiation was assessed by measuring D
10, which represents the radiation dose required to inactivate 90% of the starting spores’ population. As previously described for the viable cells, spores were irradiated with UV at λ 254 nm with doses from 125 J/m
2 up to 1000 J/m
2: Irradiation at 125 J/m
2 emerged as the D
10, thus confirming the hypothesis that UV resistance of viable cells was due to the onset of sporulation during irradiation [
13]. γ-ray irradiation, performed using
60Co, was carried out in the dose interval from 0 kGy to 15.0 kGy. The D
10 value was found at about 2.50 kGy, a value higher that the majority of spores of other species previously reported (
Table 6).
The investigation of the effect of γ-rays on some molecular cells’ structures was carried out by means of Raman spectroscopy analysis. Such a technique allowed showing that inactivation of the spores, taking place at doses higher than 2.50 kGy, followed two routes. Indeed, some spores underwent the loss of dipicolinic acid (DPA), the main component of the core. The main function of DPA is to protect proteins and DNA from the damages caused by reactive oxygen species (ROS), which in turn are produced by the radiolysis of water induced by radiations. The spores lacking DPA, which is lost after damage in the coat, are therefore not protected from the action of ROS and appeared as dark phase cells at the microscope analysis. On the other hand, the microscopy analysis allowed assessing the persistence of intact spores also at radiation levels higher than D
10. The Raman analysis showed that such spores, although still retaining their DPA content, presented a lowering in their DNA and protein integrity, as confirmed by the decrease in the relative Raman signals. In this case, the inactivation was caused by the direct interaction of γ-rays with the cellular structures and was not mediated by ROS formation [
14]. Moreover, the results obtained by subjecting either the viable cells or the spores of
P. thermantarcticus to space simulation are interesting for the issue of origin of life. Indeed, the finding of bacterial species able to resist space conditions opens new scenarios for the so-called
panspermia theory, according to which, early terrestrial life forms could have originated from bacteria transported by solar pressure from other planetary bodies. The identification of bacteria like
P. thermantarcticus that potentially could survive space transport is one main key point to support this theory.
6. Genome of P. thermantarcticus
The
P. thermantarcticus M1 genome was sequenced in 2014 by the U.S. Department of Energy Joint Genome Institute. The DOE mission is to advance genomics linked to clean energy production and environmental cleaning and characterization [
108]. The
P. thermantarcticus M1 genome was obtained under a program entitled: “Genomic Encyclopedia of Type Strains, Phase III the genomes of soil and plant-associated and newly described type strains” (
https://img.jgi.doe.gov/cgi-bin/m/main.cgi). The sequencing method applied was Illumina HiSeq 2500-1TB, and the NCBI Bioproject Accession number is PRJNA323262. The total number of sequenced bases was 3,448,881, of which 85.19% are coding bases; the DNA C + G % emerged at 43.63% and was calculated as a count of Gs and Cs divided by the total number of bases. The total genes number was 3714, of which the protein-coding genes were 96.85%, and the residual 3.15% were RNA genes. Protein-coding genes with function prediction were 2783 (79.93%), while those without function prediction were 814.
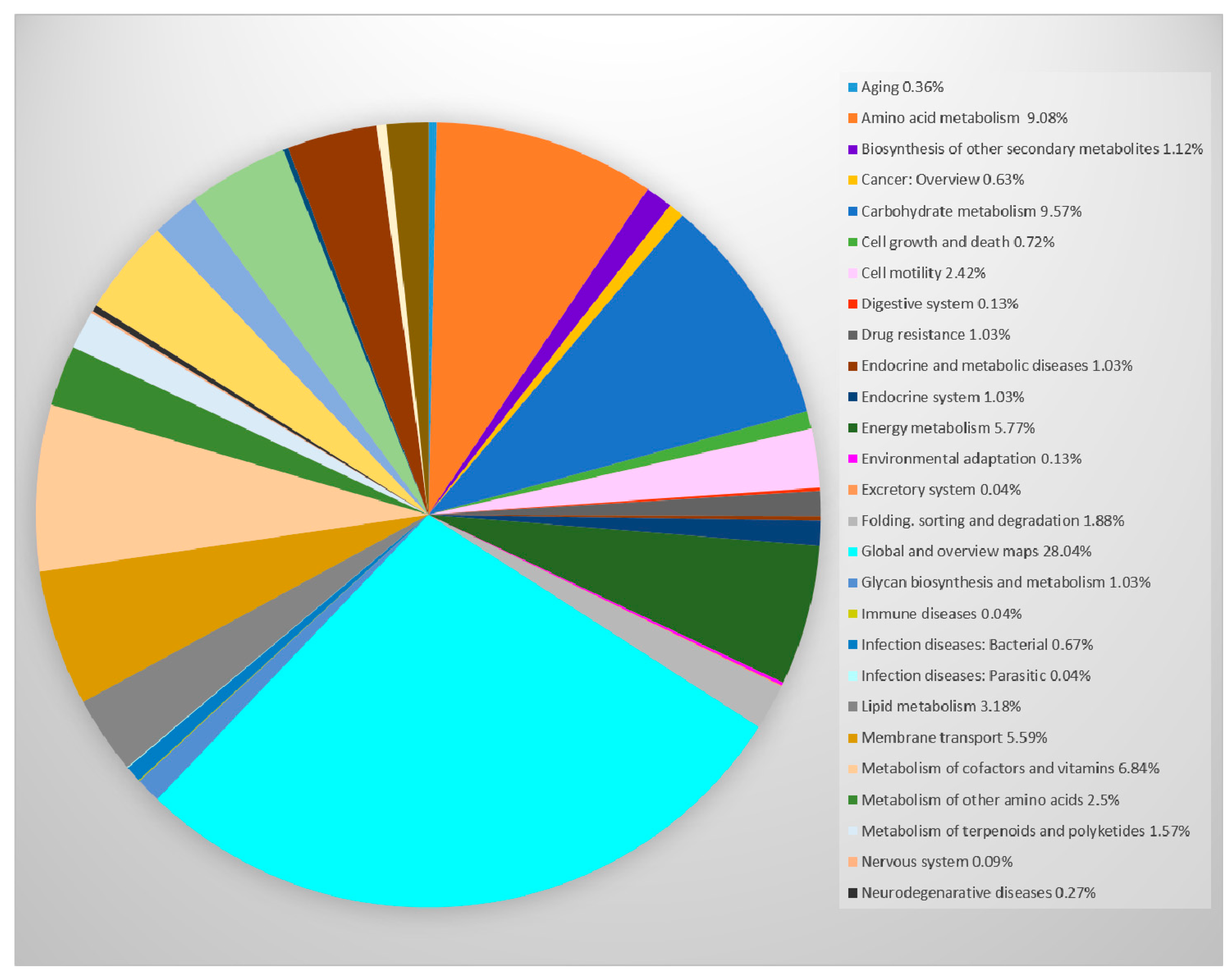
Enzyme-coding genes were 988, and protein-coding genes connected to Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways database were 1058 (
Figure 4). KEGG is a database that incorporates genomic, systemic, functional, and chemical information [
109].
As show in
Figure 4, the category “Global and overview maps” occupies the highest percentage of KEGG categories in which
P. thermantarcticus genome results are divided and relies on basal metabolism reactions such as 2-Oxocarboxylic acid metabolism; carbon metabolism; biosynthesis of antibiotics, amino acids, and secondary metabolites; degradation of aromatic compounds; fatty acid metabolism; metabolic pathways; etc. Moreover, the genes related to the carbohydrate, aminoacid, vitamin, energy metabolisms, membrane transport and signal transduction are also present. Protein-coding genes in the internal cluster were 852, while those in the chromosomal cassette were 3566. Genes found in biosynthetic clusters were 111; fused protein-coding genes were 281; protein-coding genes coding signal peptides were 108; protein-coding genes coding transmembrane proteins were 847.
Genome investigation of
P. thermantarcticus M1 showed an aerobic metabolism for the presence of ubiquinol oxidation with an oxygen (with proton transport) pathway (Pathway OID 768), as confirmed by the experimental data [
8]; in addition, the genome revealed the possibility to set up growth in anaerobic conditions thanks to the presence of a metabolism in which nitrate (NO
3−) becomes the electron acceptor for respiration in the dissimilatory nitrate reduction pathway (Pathway OID 764) and the presence of the gene coding for the alcohol dehydrogenase, class IV.
P. thermantarcticus M1 is a motile, rod-shaped bacterium, as previously reported [
8]. These morphological and mechanical properties have found confirmation in the genome reading. Indeed, there are many genes coding for the cell shape (rod shape-determining protein RodA), chemotaxis, and flagellar biosynthesis proteins.
From a chemotaxonomic point of view, Nicolaus et al. [
8] reported for
P. thermantarcticus M1 three phospholipids and one phosphoglycolipid, as major components. Genes coding for cardiolipin synthase and cardiolipin synthetase 2 (EC:2.7.8) were found.
Starting from the genome analysis, P. thermantarcticus M1 possesses a prototroph metabolism for L-aspartate, L-alanine, glycine, L-glutamate, L-glutamine, L-asparagine, Coenzyme A, and L-methionine synthesis while, an auxotroph metabolism emerged for L-phenylalanine, L-tryptophan, L-tyrosine, L-histidine, L-arginine, L-leucine, and L-serine synthesis.
P. thermantarcticus M1 produces spores [
8], and they represent a useful biological system that allows the microorganism to survive under extreme environmental conditions, including space simulation, as recently reported [
12,
13,
14]. The genome examination highlighted several genes involved in the sporulation system, ranging from genes coding for sporulation proteins and lipoprotein, prespore-specific regulator, to spore germination proteins.
Concerning sugars metabolism, sucrose-, glucose-, fructose-, and xylose-utilizing pathways were pointed out in the genome. In particular, sucrose conversion to fructose and glucose 6-phosphate and sucrose hydrolysis pathways were reported; in addition, evidence of metabolic dissimilation of glucose, the “Embden Meyerhof pathway” giving pyruvic acid, emerged; the fructose and xylose metabolisms dealing with D-Fructose 6-phosphate formation from D-fructose and D-xylose conversion to D-xylulose 5-phosphate were also found. Moreover,
P. thermantarcticus M1 utilizes glucose, trehalose, and xylose for growth, only weakly maltose and sucrose, while it does not utilize cellobiose, proprionate, citrate, and dextrin [
8]. Furthermore, the ability to utilize mannose for growth resulted in Manca et al. [
11]; in addition, xylan, arabinose, galactose, fructose can be added to the above-reported list of sugar utilization for growth as deducible from Lama et al. [
55]. Complementary information of the sugar metabolism comes from the presence in the genome of the carbohydrate ABC transporter membrane protein 2_CUT1 family, protein 1_CUT1 family, and substrate-binding protein_CUT1 family.
P. thermantarcticus M1 was described as an exopolysaccharide producer, when grown with mannose as a sole carbon and energy source [
11]. The secretion of polymers out of the cell represents an ecological manner to better support the extreme conditions in which extremophilic microorganisms live [
110]. In fact, the main ecological role of exopolysaccharides is their protective nature: EPSs, generating a layer around the cell, furnish a defense mechanism with respect to extremes of temperature and salinity or potential predators. They are indispensable in film construction, in the system of surface adhesion, and in nutrient uptake [
110,
111,
112]. Genomic evidence was found in the asserted metabolic pathway concerning acetyl–CoA assimilation via the glyoxylate cycle, responsible for the biosynthesis of polysaccharides during gluconeogenesis.
Dipasquale et al. [
80] described a proteolytic activity detected in the cell-free supernatant. Experimental results recommended that the enzyme belongs to the serine protease family (EC. 3.4.21) and had a presumed molecular mass of about 42 kDa. The analysis of genes coding for proteins revealed a huge number of genes promoting catalytic activities able to hydrolyze peptide bonds. Among them, a gene coding for serine protease S1-C subfamily, containing the C-terminal PDZ domain, was noted, for which the exact molecular weight was 43 kDa, by confirming the experimental results.
P. thermantarcticus M1 produced an intracellular xylose isomerase showing a molecular mass of about 47 kDa, with an isoelectric point of 4.8. Lama et al. [
61], going over the genome, detected the presence of a gene encoding for the abovementioned activity.
In addition,
P. thermantarcticus M1 produced, in the cell-free supernatant, xylanase and β-xylosidase activities [
55]. The genome study confirmed the experimental evidence for hemicellulose activities: Both genes encoding for endo-1,4 beta-xylanase and xylan 1,4-beta-xylosidase have been found.
At present, the P. thermantarcticus M1 genome investigation indicates the presence of many other biotechnologically interesting potentialities not yet studied. Indeed, at the time of writing, there is no experimental evidence relying on the presence of urease, pullulanase, beta-fructofuranosidase, endoglucanase, and polyhydroxyalkanoate synthase activities, which, by contrast, have been reported in genome sequencing.
Therefore, knowledge of the P. thermantarcticus M1 genome sequence supports the experimental results collected up to now; on the other hand, this suggests that different other research activities coupled with modern fermentation systems and innovative molecular biology techniques could open up new possible scenarios to better exploit the potential of this microorganism.
For instance, starting from metabolic pathways and the presence of specific genes, the search for the best enzymatic activity-inducing substrates could be performed more easily; in addition, gene cloning strategies in a suitable engineered host could improve the catalyst and the corresponding hydrolysis products’ productivities. In this view, in the literature, there are many examples of hemicellulolitic genes cloning aiming at higher activity expression [
113]. The gene coding for a β-xylosidase activity present in the genome of the thermophilic
Geobacillus stearothermophilus microorganism was cloned and expressed in
E. coli. In addition, an amino acid substitution at position 509 extended the enzymatic hydrolysis reaction to the long-chain xylo-oligomers [
53].

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}