Invasive Goldenrod (Solidago gigantea) Influences Soil Microbial Activities in Forest and Grassland Ecosystems in Central Europe


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Soil Sampling
- Forest Uninvaded (FU): Ten study sites were located in stands dominated by Quercus sp., Fagus sp., Carpinus sp. and Betula sp. (deciduous forests).
- Grassland Uninvaded (GU): Ten study sites with indigenous multispecies vegetation dominated by Dactylis glomerata, Lolium perenne, Trifolium pratense, Capsella bursa-pastoris and Taraxacum officinale.
- Forest Invaded (FI): Ten adjacent study sites with monospecific stand of S. gigantea on forest edge, with an estimated time of invasion of 10–15 years.
- Grassland Invaded (GI): Ten adjacent study sites invaded by S. gigantea, with an estimated time of invasion of 10–20 years.
2.2. Soil Physicochemical Analysis and Enzyme Assays
2.3. Statistical Analyses
3. Results
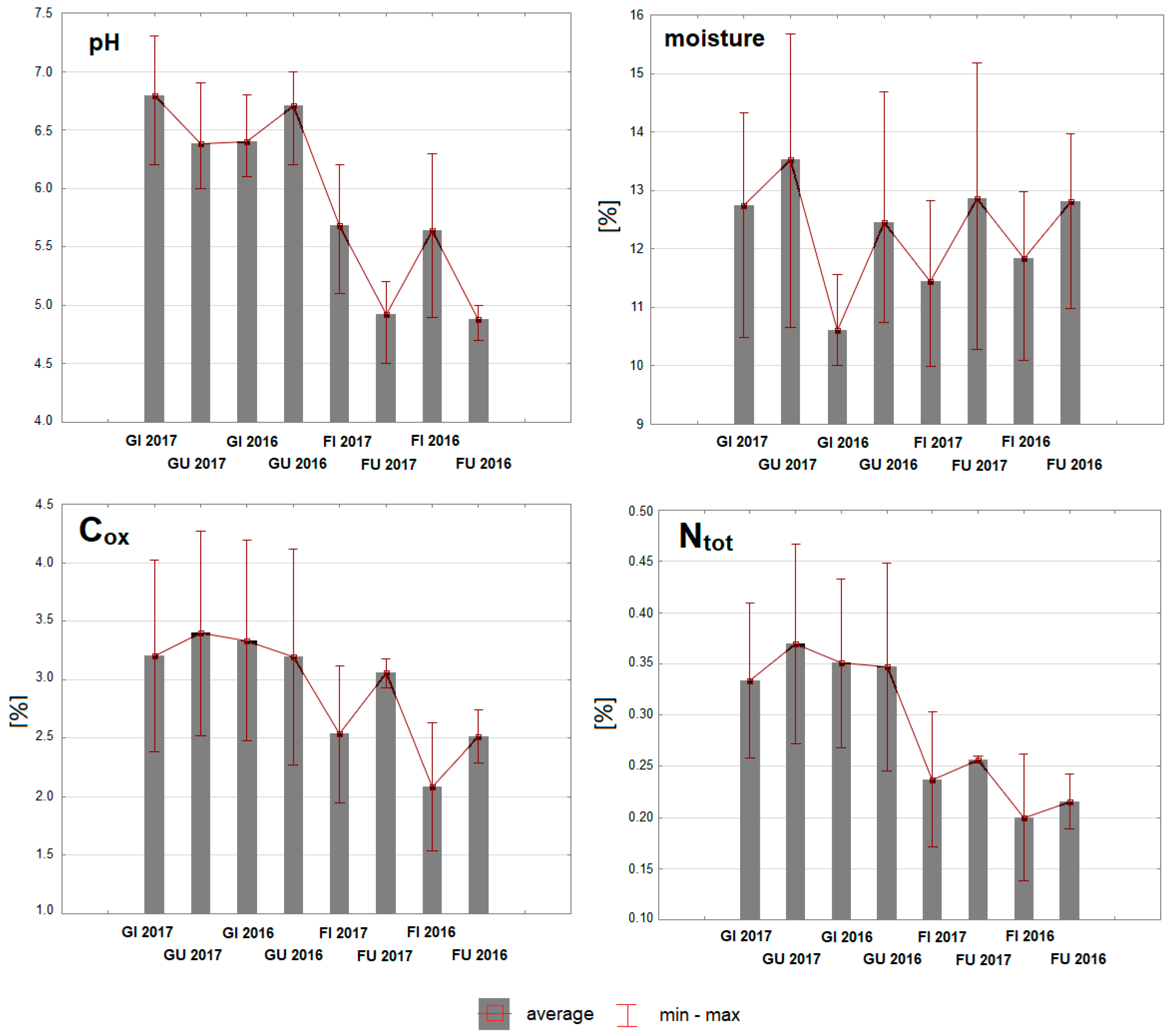
3.1. Soil Physicochemical Properties
3.2. Microbial Indicators and Enzyme Assays
3.3. Correlation Analysis of Soil Characteristics between Ecosystem Invasive Status and Microbial Parameters with Environmental Variables
4. Discussion
4.1. Soil Physicochemical Properties
4.2. FDA and Soil Enzymes Activity
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Herrera, I.; Ferrer-Paris, J.R.; Benzo, D.; Flores, S.; García, B.; Nassar, J.M. An invasive succulent plant (Kalanchoe daigremontiana) influences soil carbon and nitrogen mineralization in a neotropical semiarid zone. Pedosphere 2018, 28, 632–643. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Häggblom, M. Exotic plant species alter the microbial community structure and function in the soil. Ecology 2002, 83, 3152–3166. [Google Scholar] [CrossRef]
- Aguilera, A.G.; Alpert, P.; Dukes, J.S.; Harrington, R. Impact of the invasive plant Fallopia japonica (Houtt.) on plant communities and ecosystem processes. Biol. Invasions 2010, 12, 1243–1252. [Google Scholar] [CrossRef]
- Peltzer, D.A.; Bellingham, P.J.; Kurokawa, H.; Walker, L.R.; Wardle, D.A.; Yeates, G.W. Punching above their weight: Low-biomass non-native plant species alter soil properties during primary succession. Oikos 2009, 118, 1001–1014. [Google Scholar] [CrossRef]
- Moles, A.T.; Gruber, M.A.M.; Bonser, S.P. A new framework for predicting invasive plant species. J. Ecol. 2008, 96, 13–17. [Google Scholar] [CrossRef]
- Scharfy, D.; Güsewell, S.; Gessner, M.O.; Venterink, H.O. Invasion of Solidago gigantea in contrasting experimental plant communities: Effects on soil microbes, nutrients and plant-soil feedbacks. J. Ecol. 2010, 98, 1379–1388. [Google Scholar] [CrossRef]
- Vinhal-Freitas, I.C.; Corrêa, G.F.; Wendling, B.; Bobuľská, L.; Ferreira, A.S. Soil textural class plays a major role in evaluating the effects of land use on soil quality indicators. Ecol. Indic. 2017, 74, 182–190. [Google Scholar] [CrossRef]
- Batlle-Bayer, L.; Batjes, N.H.; Bindraban, P.S. Changes in organic carbon stocks upon land conversion in the Brazilian Cerrado: A review. Agric. Ecosyst. Environ. 2010, 137, 47–58. [Google Scholar] [CrossRef]
- Feng, J.; Wang, S.; Wang, S.; Ying, R.; Yin, F.; Jiang, L.; Li, Z. Effects of invasive Spartina alternifolia Loisel. and subsequent ecological replacement by Sonneratia apetala Buch.-Ham. on soil organic carbon fraction and stock. Forests 2019, 10, 171. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, W.; Luo, J.; Donnison, A. Changes in soil organic carbon dynamics in an Eastern Chinese coastal wetland following invasion by a C4 plant Spartina alterniflora. Soil Biol. Biochem. 2010, 42, 1712–1720. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Daigh, A.L.; Veenstra, J.J.; Engle, D.M.; Miller, J.R.; Debinski, D.M. Connecting soil organic carbon and root biomass with land-use and vegetation in temperate grassland. Sci. World J. 2014, 1, 487563. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sardans, J.; Wang, C.; Zeng, C.; Tong, C.; Chen, G.; Huang, J.; Pan, H.; Peguero, G.; Vallicrosa, H.; et al. The response of stock of C, N, and P to plant invasion in the coastal wetlands of China. Glob. Chang. Biol. 2019, 25, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Bobuľská, L.; Fazekašová, D.; Angelovičová, L.; Kotorová, D. Impact of ecological and conventional farming systems on chemical and biological soil quality indices in a cold mountain climate in Slovakia. Biol. Agric. Hortic. 2015, 31, 205–218. [Google Scholar] [CrossRef]
- Constán-Nava, S.; Soliveres, S.; Torices, R.; Serra, L.; Bonet, A. Direct and indirect effects of invasion by the alien tree Ailanthus altissima on riparian plant communities and ecosystem multifunctionality. Biol. Invasions 2015, 17, 1095–1108. [Google Scholar] [CrossRef]
- Pauková, Ž. Invasive plant species in the three microregions of Nitra region, South-West Slovakia. Ekológia Bratisl. 2013, 32, 262–266. [Google Scholar] [CrossRef]
- Zhang, C.B.; Wang, J.; Qian, B.Y.; Li, W.H. Effects of the invader Solidago canadensis on soil properties. Appl. Soil Ecol. 2009, 43, 163–169. [Google Scholar] [CrossRef]
- Pimentel, D.; McNair, S.; Janecka, J.; Wightman, J.; Simmonds, C.; OʹConnell, C.; Wong, E.; Russel, L.; Zern, J.; Aquino, T.; et al. Economic and environmental threats of alien plant, animal, and microbe invasions. Agric. Ecosyst. Environ. 2001, 84, 1–20. [Google Scholar] [CrossRef]
- Baranová, B.; Manko, P.; Jászay, T. Differences in surface-dwelling beetles of grasslands invaded and non-invaded by goldenrods (Solidago canadensis, S. gigantea) with special reference to Carabidae. J. Insect Conserv. 2014, 18, 623–635. [Google Scholar] [CrossRef]
- Roháčová, M.; Drozd, P. How many heteropteran species can live on alien goldenrods Solidago canadensis and S. gigantea in Europe? Biologia 2009, 64, 981–993. [Google Scholar] [CrossRef]
- Jakobs, G.; Weber, E.; Edwards, P.J. Introduced plants of the invasive Solidago gigantea (Asteraceae) are larger and grow denser than conspecifics in the native range. Divers. Distrib. 2004, 10, 11–19. [Google Scholar] [CrossRef]
- Guo, S.L.; Fang, F. Physiological adaptation of the invasive plant Solidago canadensis to environments. Acta Physiol. Sin. 2003, 27, 47–52. [Google Scholar]
- Bobuľská, L.; Čekanová, K.; Demková, L.; Oboňa, J.; Sarvaš, J. Evaluation of the phytoremediation properties of the invasive species Solidago genus. Ann. Univ. Craiova 2018, 23, 314–320. [Google Scholar]
- Stefanowicz, A.M.; Stanek, M.; Nobis, M.; Zubek, S. Species-specific effects of plant invasions on activity, biomass, and composition of soil microbial communities. Biol. Fertil. Soils 2016, 52, 841–852. [Google Scholar] [CrossRef] [Green Version]
- Fiala, K.; Barančikova, G.; Brečkova, V.; Burik, V.; Houškova, B.; Chomaničova, A.; Kobza, J.; Litavec, T.; Makovnikova, L.; Pechova, B.; et al. Partial Monitoring System–Soil. Binding Methods, 1st ed.; VÚPOP: Bratislava, Slovakia, 1999; 139p. [Google Scholar]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Green, V.S.; Stott, D.E.; Diack, M. Assay for fluorescein diacetate hydrolytic activity: Optimization for soil samples. Soil Biol. Biochem. 2006, 38, 693–701. [Google Scholar] [CrossRef]
- Grejtovský, A. Influence of soil improvers on enzymatic activity of heavy alluvial soil. Plant Soil Environ. 1991, 37, 289–295. [Google Scholar]
- Khaziev, F.K. Soil Enzyme Activity, 1st ed.; Nauka: Moscow, Russia, 1976; 180p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing, 1st ed.; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 22 February 2019).
- Van der Putter, W.H.; Klironomos, J.N.; Wardle, D.A. Microbial ecology of biological invasions. ISME 2007, 1, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusterholz, H.P.; Schneuwly, J.; Baur, B. Invasion of the alien shrub Prunus laurocerasus in suburban deciduous forests: Effects on native vegetation and soil properties. Acta Oecol. 2018, 92, 44–51. [Google Scholar] [CrossRef]
- Richard, M.; Tallamy, D.W.; Mitchell, A.B. Introduced plants reduce species interactions. Biol. Invasions 2019, 21, 983–992. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Fisher, R.A.; Wardle, D.A. Plant communities as drivers of soil respiration: Pathways, mechanisms, and significance for global changes. Biogeosciences 2011, 8, 2047–2061. [Google Scholar] [CrossRef]
- Koutika, L.S.; Vanderhoeven, S.; Chapuis-Lardy, L.; Dassonville, N.; Meerts, P. Assessment of changes in soil organic matter after invasion by exotic plant species. Biol. Fertil. Soils 2007, 44, 331–341. [Google Scholar] [CrossRef]
- Liao, C.Z.; Peng, R.H.; Luo, Y.Q.; Zhou, X.H.; Wu, X.W.; Fang, C.M.; Chen, J.K.; Li, B. Altered ecosystem carbon and nitrogen cycles by plant invasion: A meta-analysis. New Phytol. 2008, 177, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yan, Y.; Jiang, F.; Cheng, X.; An, S. Response of the soil microbial community composition and biomass to a short-term Spartina alternifolia invasion in a coastal wetland of eastern China. Plant Soil 2016, 408, 443–456. [Google Scholar] [CrossRef]
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Driscoll, D.A. Disturbance maintains native and exotic plant species richness in invaded grassy woodlands. J. Veg. Sci. 2017, 28, 573–584. [Google Scholar] [CrossRef]
- Stark, J.M.; Firestone, M.K. Mechanisms for soil moisture effects on activity of nitrifying bacteria. Appl. Environ. Microb. 1995, 61, 218–221. [Google Scholar] [Green Version]
- Stefanowicz, A.M.; Stanek, M.; Nobis, M.; Zubek, S. Few effects of invasive plants Reynoutria japonica, Rudbeckia laciniata and Solidago gigantea on soil physical and chemical properties. Sci. Total Environ. 2017, 574, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Sheahan, C.M.; Bray, D.B.; Bhat, M.G.; Jayachandran, K. Ecological, economic, and organizational dimension of organic farming in Miami-Dade country. J. Sustain. Agric. 2012, 36, 83–105. [Google Scholar] [CrossRef]
- Baranová, B.; Fazekašová, D.; Manko, P. Variations of selected soil properties in the grass fields invaded and uninvaded by invasive goldenrod (Solidago canadensis L.). Ekológia Bratisl. 2017, 36, 101–111. [Google Scholar] [CrossRef]
- Quist, C.W.; Vervoort, M.T.W.; Van Megen, H.; Gort, G.; Bakker, J.; Van der Putten, W.H.; Helder, J. Selective alteration of soil food web components by invasive giant goldenrod Solidago gigantea in two distinct habitat types. Oikos 2014, 123, 837–845. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Chacón, N.; Herrera, I.; Flores, S.; Gonzáles, J.A.; Nassar, J.M. Chemical, physical, and biochemical soil properties and plant roots as affected by native and exotic plants in neotropical arid zones. Biol. Fertil. Soils 2009, 45, 321–328. [Google Scholar] [CrossRef]
- Sicardi, M.; Garcìa-Prèchac, F.; Frioni, L. Soil microbial indicators sensitive to land use conversion from pastures to commercial Eucalyptus grandis (Hill ex Maiden) plantations in Uruguay. Appl. Soil Ecol. 2004, 27, 125–133. [Google Scholar] [CrossRef]
- Wang, X.; Lu, Q. Beta-glucosidase activity in paddy soils of the Taihu lake region, China. Pedosphere 2006, 16, 118–124. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Classen, A.T.; Simberloff, D. Two co-occurring invasive woody shrubs alter soil properties and promote subdominant invasive species. J. Appl. Ecol. 2014, 51, 124–133. [Google Scholar] [CrossRef]
- Sheng, Q.; Zhao, B.; Huang, M.; Wang, L.; Quan, Z.; Fang, C.; Li, B.; Wu, J. Greenhouse gas emission following an invasive eradication program. Ecol. Eng. 2014, 73, 229–237. [Google Scholar] [CrossRef]
- Speir, T.W.; Van Schaik, A.P.; Lloyd-Jones, A.R. Temporal response of soil biochemical properties in a pastoral soil alter cultivation following high application rates of undigested sewage sludge. Biol. Fertil. Soils 2003, 38, 377–385. [Google Scholar] [CrossRef]
- Corstanaje, R.; Schulin, R.; Lark, R.M. Scale-dependent relationships between soil organic carbon and urease activity. Eur. J. Soil Sci. 2007, 58, 1087–1095. [Google Scholar] [CrossRef]
- Ge, Y.; Wang, Q.; Wang, L.; Liu, W.; Liu, X.; Huang, Y.; Christie, P. Response of soil enzymes and microbial communities to root extracts of the alien Alternanthera philoxeroides. Arch. Agron. Soil Sci. 2018, 64, 708–717. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Häggblom, M. Experimental analysis of the effect of exotic and native plant species on the structure and function of soil microbial communities. Soil Biol. Biochem. 2003, 35, 895–905. [Google Scholar] [CrossRef]
- Bobuľská, L.; Macková, D.; Malina, R.; Demková, L. Occurrence and dynamics of Impatiens parviflora depending on various environmental conditions in the protected areas in Slovakia. Eur. J. Ecol. 2016, 2, 87–98. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Enzyme | Incubation Condition | Reference | ||||
|---|---|---|---|---|---|---|
| Substrate | Buffer (pH) | Temperature (°C) | Soil (g) | Time (h) | ||
| BGL | 4-Nitrophenyl glucopyranoside | MUB (6.0) | 37 | 1 | 3 | [25] |
| FDA | Fluorescein diacetate | PPB (7.6) | 30 | 1 | 1 | [26] |
| PHOSAL | p-Nitrophenyl phosphate | AB (5.0) | 37 | 5 | 3 | [27] |
| PHOSAC | p-Nitrophenyl phosphate | BB (10.0) | 37 | 5 | 3 | [27] |
| URE | Urea | PB (6.7) | 37 | 5 | 24 | [28] |
| Parameter | U | z | p Value | |
|---|---|---|---|---|
| pH | Between ecosystems | 49.5 | −7.2 | 0.0001 * |
| MOIS | 798 | −0.1 | 0.98 | |
| Cox | 525 | −2.64 | 0.008 * | |
| Ntot | 225 | −5.52 | 0.0001 | |
| BGL | 659 | 1.34 | 0.17 | |
| FDA | 624 | 1.68 | 0.05 * | |
| URE | 717 | 0.79 | 0.42 | |
| PHOSAC | 544 | 2.45 | 0.01 * | |
| PHOSAL | 248 | −5.8 | 0.0001* |
| Parameter | F | G | |||||
|---|---|---|---|---|---|---|---|
| U | z | p Value | U | z | p Value | ||
| pH | Between invaded and uninvaded sides | 30 | 4.58 | 0.0001 * | 190 | 0.25 | 0.79 |
| MOIS | 103 | −2.61 | 0.009 * | 102 | −2.63 | 0.008 * | |
| Cox | 100 | −2.69 | 0.007 * | 175 | −0.66 | 0.50 | |
| Ntot | 200 | 0.02 | 1.0 | 150 | −1.33 | 0.18 | |
| BGL | 68 | 3.55 | 0.001 * | 96 | 2.79 | 0.005 * | |
| FDA | 186 | −3.6 | 0.71 | 64 | −3.66 | 0.0002 * | |
| URE | 55 | −3.9 | 0.0001 * | 105 | −2.54 | 0.01 * | |
| PHOSAC | 161 | −1.04 | 0.29 | 190 | −0.25 | 0.79 | |
| PHOSAL | 90 | 2.96 | 0.003 * | 95 | 2.85 | 0.004 * | |
| Parameter | pH | MOIST | Cox | Ntot | BLG | FDA | URE | PHOSAC | PHOSAL |
|---|---|---|---|---|---|---|---|---|---|
| pH | −0.08 | −0.07 | 0.30 ** | 0.51 ** | −0.01 | −0.28 * | −0.39 | −0.31 * | 0.59 |
| MOIS | 0.38 ** | 0.35 ** | 0.16 | 0.55 ** | 0.05 | 0.23 * | −0.22 | ||
| Cox | 0.96 ** | 0.17 | 0.21 | −0.11 | 0.17 | −0.06 | |||
| Ntot | 0.14 | 0.17 | −0.18 | 0.11 | 0.17 | ||||
| BGL | 0.12 | −0.31 ** | 0.12 | −0.14 | |||||
| FDA | −0.05 | 0.21 | −0.20 | ||||||
| URE | 0.08 | −0.33 ** | |||||||
| PHOSAC | −0.13 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bobuľská, L.; Demková, L.; Čerevková, A.; Renčo, M. Invasive Goldenrod (Solidago gigantea) Influences Soil Microbial Activities in Forest and Grassland Ecosystems in Central Europe. Diversity 2019, 11, 134. https://doi.org/10.3390/d11080134
Bobuľská L, Demková L, Čerevková A, Renčo M. Invasive Goldenrod (Solidago gigantea) Influences Soil Microbial Activities in Forest and Grassland Ecosystems in Central Europe. Diversity. 2019; 11(8):134. https://doi.org/10.3390/d11080134
Chicago/Turabian StyleBobuľská, Lenka, Lenka Demková, Andrea Čerevková, and Marek Renčo. 2019. "Invasive Goldenrod (Solidago gigantea) Influences Soil Microbial Activities in Forest and Grassland Ecosystems in Central Europe" Diversity 11, no. 8: 134. https://doi.org/10.3390/d11080134
APA StyleBobuľská, L., Demková, L., Čerevková, A., & Renčo, M. (2019). Invasive Goldenrod (Solidago gigantea) Influences Soil Microbial Activities in Forest and Grassland Ecosystems in Central Europe. Diversity, 11(8), 134. https://doi.org/10.3390/d11080134