Influence of Forest Structure and Composition on Summer Habitat Use of Wildlife in an Upland Hardwood Forest


Abstract
:1. Introduction
2. Materials and Methods
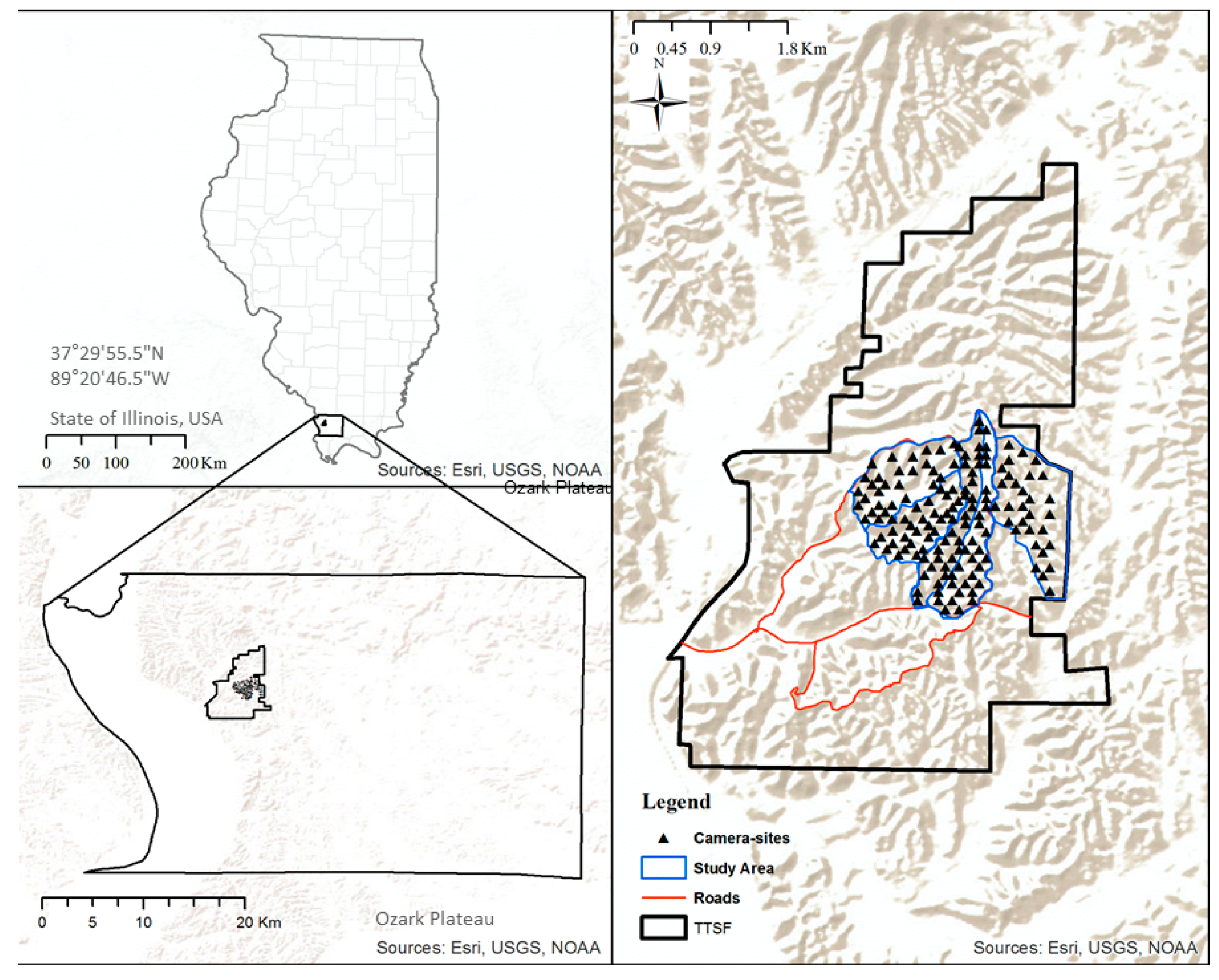
2.1. Study Area
2.2. Site Selection
2.3. Remote Camera Surveys
2.4. Habitat Characteristics
2.5. Occupancy Modeling
3. Results
3.1. Habitat Characteristics
3.2. Survey Effort and Wildlife Detections
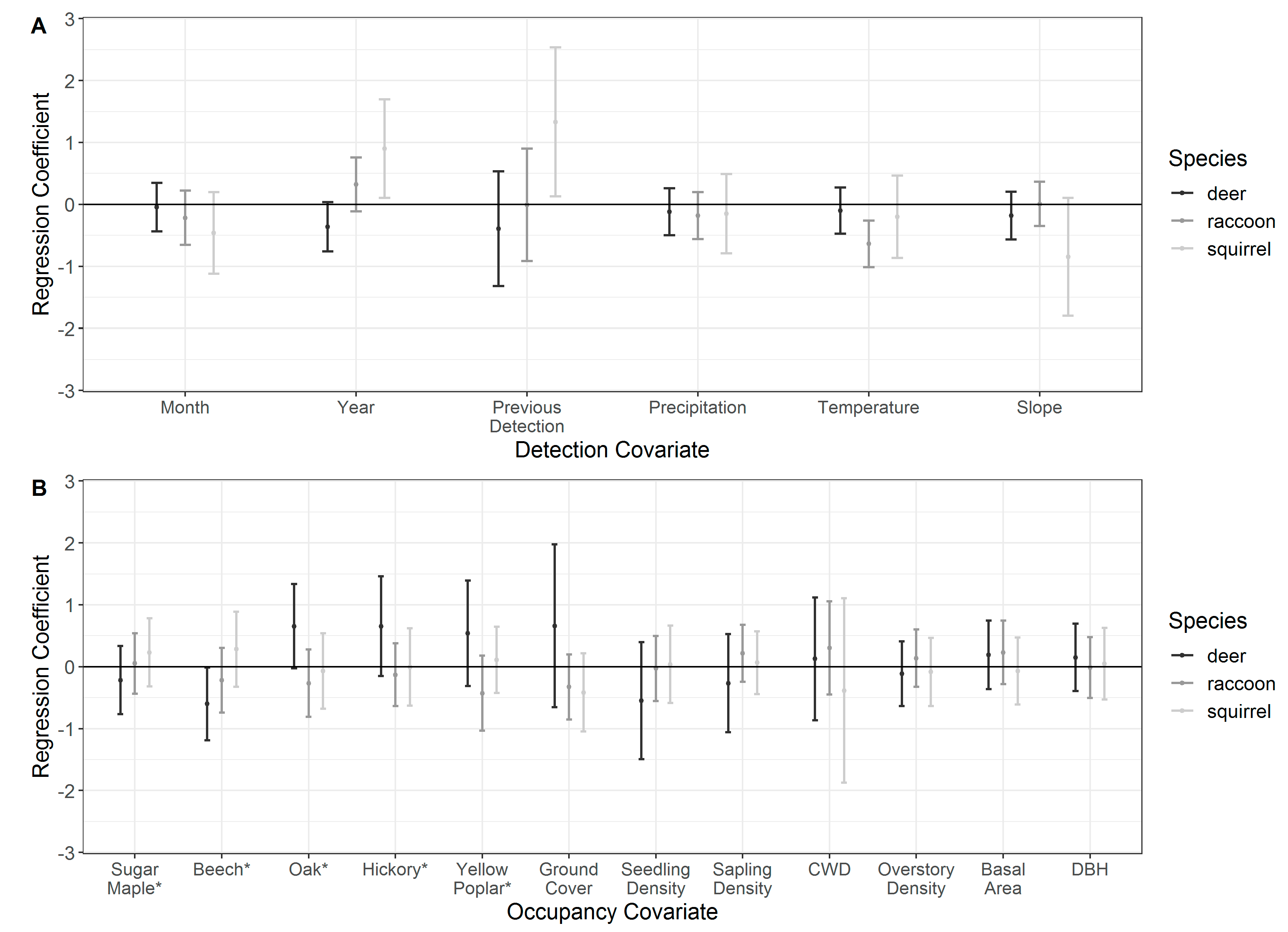
3.3. Detection
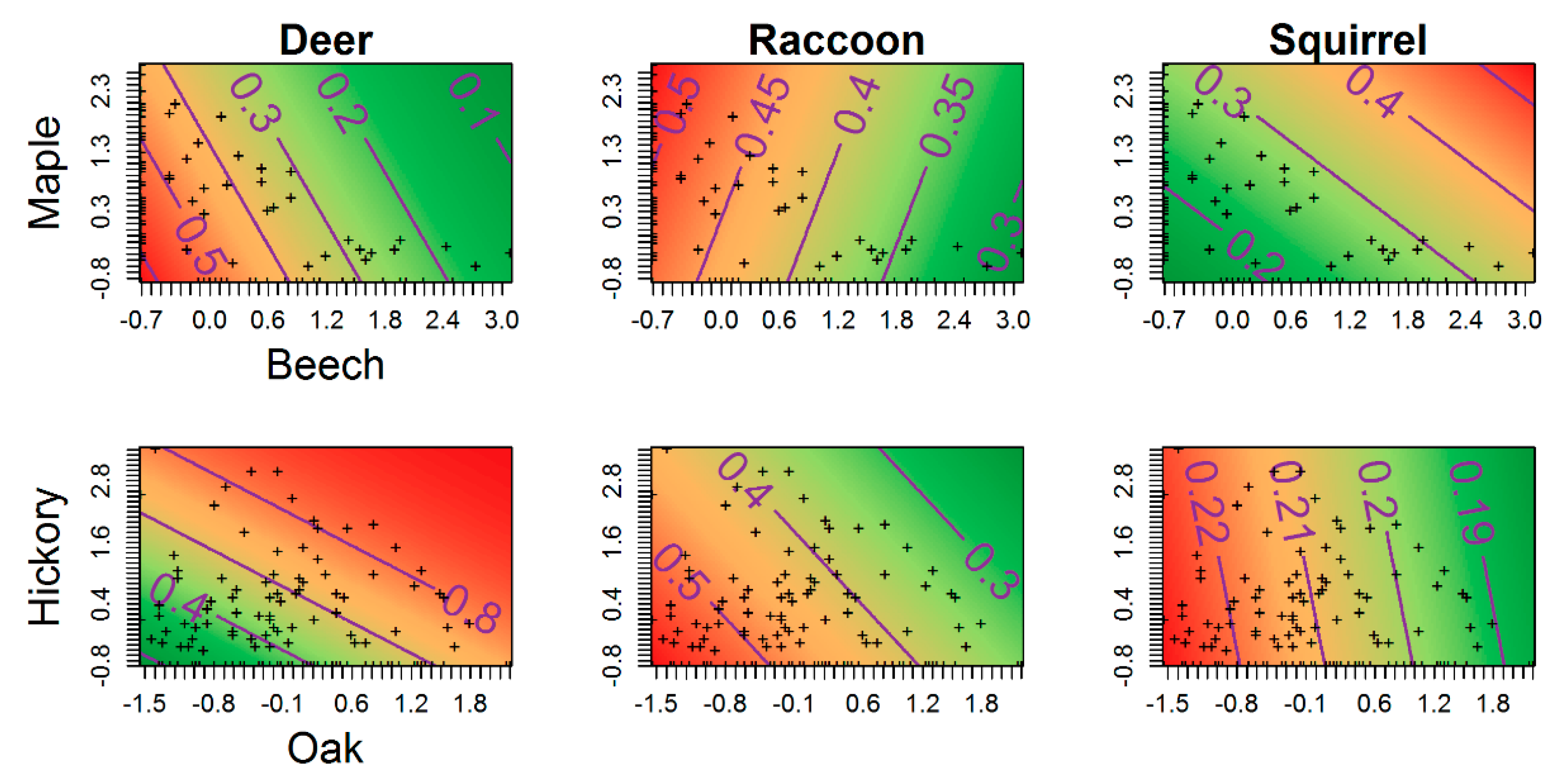
3.4. Occupancy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, P.S.; Shifley, S.R.; Rogers, R. The Ecology and Silviculture of Oaks; CABI: Boston, MA, USA, 2009. [Google Scholar]
- Abrams, M.D. Fire and the Development of Oak Forests—In Eastern North-America, Oak Distribution Reflects a Variety of Ecological Paths and Disturbance Conditions. Bioscience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Lorimer, C.G. Development of the Red Maple Understory in Northeastern Oak Forests. Forest Sci. 1984, 30, 3–22. [Google Scholar]
- Ozier, T.B.; Groninger, J.W.; Ruffner, C.M. Community composition and structural changes in a managed Illinois Ozark Hills Forest. Am. Midl. Nat. 2006, 155, 253–269. [Google Scholar] [CrossRef]
- Abrams, M.D. Where has all the white oak gone? Bioscience 2003, 53, 927–939. [Google Scholar] [CrossRef]
- McEwan, R.W.; Dyer, J.M.; Pederson, N. Multiple interacting ecosystem drivers: Toward an encompassing hypothesis of oak forest dynamics across eastern North America. Ecography 2011, 34, 244–256. [Google Scholar] [CrossRef]
- Van de Gevel, S.; Ozier, T.B.; Ruffner, C.M.; Groninger, J.W. Relating past land-use, topography, and forest dynamics in the Illinois Ozark Hills. US Serv. T R Nc 2003, 234, 246. [Google Scholar]
- Holzmueller, E.J.; Gibson, D.J.; Suchecki, P.F. Accelerated succession following an intense wind storm in an oak-dominated forest. Forest Ecol. Manag. 2012, 279, 141–146. [Google Scholar] [CrossRef]
- Fralish, J.S. The keystone role of oak and hickory in the central hardwood forest. In Upland Oak Ecology Symposium: History, Current Conditions, and Sustainability; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2004; p. 311. [Google Scholar]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- McShea, W.J.; Healy, W.M.; Devers, P.; Fearer, T.; Koch, F.H.; Stauffer, D.; Waldon, J. Forestry matters: Decline of oaks will impact wildlife in hardwood forests. J. Wild. Manag. 2007, 71, 1717–1728. [Google Scholar] [CrossRef]
- Rodewald, A.D. Decline of oak forests and implications for forest wildlife conservation. Nat. Area J. 2003, 23, 368–371. [Google Scholar]
- Martin, A.C.; Zim, H.S.; Nelson, A.L. American Wildlife and Plants: A guide to Wildlife Food Habits; McGraw-Hill Book Company, Inc.: New York, NY, USA, 1951; p. 500. [Google Scholar]
- Van Dersal, W.R. Utilization of oaks by birds and mammals. J. Wild. Manag. 1940, 4, 404–428. [Google Scholar] [CrossRef]
- Johnson, A.S.; Hale, P.E.; Ford, W.M.; Wentworth, J.M.; French, J.R.; Anderson, O.F.; Pullen, G.B. White-Tailed Deer Foraging in Relation to Successional Stage, Overstory Type and Management of Southern Appalachian Forests. Am. Midl. Nat. 1995, 133, 18–35. [Google Scholar] [CrossRef]
- Feldhamer, G. Acorns and White-Tailed Deer: Interrelationships in Forest Ecosystems. In Oak Forest Ecosystems: Ecology and Management for Wildlife; McShea, W.J., Healy, W.M., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2002; pp. 215–223. [Google Scholar]
- McShea, W.J.; Schwede, G. Variable Acorn Crops—Responses of White-Tailed Deer and Other Mast Consumers. J. Mammal. 1993, 74, 999–1006. [Google Scholar] [CrossRef]
- Ostfeld, R.S. Ecological Webs Involving Acorns and Mice. In Oak Forest Ecosystems: Ecology and Management for Wildlife; McShea, W.J., Healy, W.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2002; pp. 196–214. [Google Scholar]
- Rodewald, A.D.; Abrams, M.D. Floristics and avian community structure: Implications for regional changes in eastern forest composition. For. Sci. 2002, 48, 267–272. [Google Scholar]
- Grado, S.C.; Hunt, K.M.; Whiteside, M.W. Economic impacts of white-tailed deer hunting in Mississippi. Proc. South. Assoc. Fish Wildlife Agencies 2007, 61, 59–67. [Google Scholar]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Ford, W.M.; Johnson, A.S.; Hale, P.E.; Wentworth, J.M. Availability and use of spring and summer woody browse by deer in clearcut and uncut forests of the southern Appalachians. South. J. Appl. Forest. 1993, 17, 116–119. [Google Scholar] [CrossRef]
- Korschgen, L.J. Foods of Missouri deer, with some management implications. J. Wild. Manag. 1962, 26, 164–172. [Google Scholar] [CrossRef]
- Korschgen, L.J.; Porath, W.R.; Torgerson, O. Spring and summer foods of deer in the Missouri Ozarks. J. Wild. Manag. 1980, 89–97. [Google Scholar] [CrossRef]
- Yeager, L.E.; Elder, W.H. Pre-and post-hunting season foods of raccoons on an Illinois goose refuge. J. Wild. Manag. 1945, 9, 48–56. [Google Scholar] [CrossRef]
- Johnson, D.H. The Comparison of Usage and Availability Measurements for Evaluating Resource Preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence; Academic Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Welbourne, D.J.; Claridge, A.W.; Paull, D.J.; Lambert, A. How do passive infrared triggered camera traps operate and why does it matter? Breaking down common misconceptions. Remot. Sens. in Ecol. Conservat. 2016, 2, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Meek, P.D.; Ballard, G.; Claridge, A.; Kays, R.; Moseby, K.; O’Brien, T.; O’Connell, A.; Sanderson, J.; Swann, D.E.; Tobler, M.; et al. Recommended guiding principles for reporting on camera trapping research. Biodivers. Conserv. 2014, 23, 2321–2343. [Google Scholar] [CrossRef]
- Cusack, J.J.; Dickman, A.J.; Rowcliffe, J.M.; Carbone, C.; Macdonald, D.W.; Coulson, T. Random versus Game Trail-Based Camera Trap Placement Strategy for Monitoring Terrestrial Mammal Communities. PLoS ONE 2015, 10, e0126373. [Google Scholar] [CrossRef] [PubMed]
- Lesmeister, D.B.; Nielsen, C.K.; Schauber, E.M.; Hellgren, E.C. Spatial and Temporal Structure of a Mesocarnivore Guild in Midwestern North America. Wildlife Monogr. 2015, 191, 1–61. [Google Scholar] [CrossRef]
- VerCauteren, K.; Hygnstrom, S.E. Managing white-tailed deer: Midwest North America. In Biology and Management of White-Tailed Deer; Hewitt, D.G., Ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 501–535. [Google Scholar]
- McShea, W.J.; Healy, W.M. Oak Forest Ecosystems: Ecology and Management for Wildlife; Johns Hopkins University Press: Baltimore, MD, USA, 2002; p. xiii. 432p. [Google Scholar]
- Jenkins, M.A.; Webster, C.R.; Parker, G.R.; Spetich, M.A. Coarse woody debris in managed Central Hardwood Forests of Indiana, USA. For. Sci. 2004, 50, 781–792. [Google Scholar]
- Dee, D.P.; Uppala, S.; Simmons, A.; Berrisford, P.; Poli, P.; Kobayashi, S.; Andrae, U.; Balmaseda, M.; Balsamo, G.; Bauer, D.P. The ERA-Interim reanalysis: Configuration and performance of the data assimilation system. Quar. J. Royal Meteorol. Soc. 2011, 137, 553–597. [Google Scholar] [CrossRef]
- Mesinger, F.; DiMego, G.; Kalnay, E.; Mitchell, K.; Shafran, P.C.; Ebisuzaki, W.; Jović, D.; Woollen, J.; Rogers, E.; Berbery, E.H. North American regional reanalysis. Bull. Am. Meteorol. Soc. 2006, 87, 343–360. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Fiske, I.J.; Chandler, R.B. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q) AIC (c). Available online: https://cran.r-project.org/package=AICcmodavg (accessed on 5 January 2015).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. xxvi. 488p. [Google Scholar]
- Kéry, M.; Royle, J.A. Applied Hierarchical Modeling in Ecology: Analysis of Distribution, Abundance and Species Richness in R and BUGS. Volume 1, Prelude and Static Models; Academic Press & Elsevier: Cambridge, MA, USA, 2016; Volume 1, p. 808. [Google Scholar]
- MacKenzie, D.I.; Bailey, L.L. Assessing the fit of site-occupancy models. J. Agric. Biol. Environ. Stat. 2004, 9, 300–318. [Google Scholar] [CrossRef]
- Holzmueller, E.J.; Groninger, J.W.; Ruffner, C.M.; Ozier, T.B. Composition of Oak Stands in the Illinois Ozark Hills 2 Decades following Light Harvesting and No Cutting. North. J. Appl. For. 2011, 28, 50–53. [Google Scholar] [CrossRef] [Green Version]
- Fei, S.; Kong, N.; Steiner, K.C.; Moser, W.K.; Steiner, E.B. Change in oak abundance in the eastern United States from 1980 to 2008. For. Ecol. Manag. 2011, 262, 1370–1377. [Google Scholar] [CrossRef]
- Cove, M.V.; Jones, B.M.; Bossert, A.J.; Clever, D.R., Jr.; Dunwoody, R.K.; White, B.C.; Jackson, V.L. Use of camera traps to examine the mesopredator release hypothesis in a fragmented Midwestern landscape. Am. Mid. Nat. 2012, 168, 456–465. [Google Scholar] [CrossRef]
- Pease, B.S.; Nielsen, C.K.; Holzmueller, E.J. Single-camera trap survey designs miss detections: Impacts on estimates of occupancy and community metrics. PLoS ONE 2016, 11, e0166689. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, A.F.; Talancy, N.W.; Bailey, L.L.; Sauer, J.R.; Cook, R.; Gilbert, A.T. Estimating site occupancy and detection probability parameters for meso- and large mammals in a coastal ecosystem. J. Wild. Manag. 2006, 70, 1625–1633. [Google Scholar] [CrossRef]
- Rovero, F.; Martin, E.; Rosa, M.; Ahumada, J.A.; Spitale, D. Estimating Species Richness and Modelling Habitat Preferences of Tropical Forest Mammals from Camera Trap Data. PLoS ONE 2014, 9, e103300. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.L.; Gee, K.L.; Strickland, B.K.; Demarais, S.; DeYoung, R.W. Measuring fine-scale white-tailed deer movements and environmental influences using GPS collars. Int. J. Ecol. 2010, 2010, 459610. [Google Scholar] [CrossRef]
- Bowman, J.L.; Kochanny, C.O.; Demarais, S.; Leopold, B.D. Evaluation of a GPS collar for white-tailed deer. Wild. Soc. Bull. 2000, 141–145. [Google Scholar]
- Boyce, M.S.; McDonald, L.L. Relating populations to habitats using resource selection functions. Trends Ecol. Evol. 1999, 14, 268–272. [Google Scholar] [CrossRef]
- LaRue, M.A.; Nielsen, C.K.; Grund, M.D. Using distance sampling to estimate densities of white-tailed deer in south-central Minnesota. Prairie Nat. 2007, 39, 57–68. [Google Scholar]
- Canham, C.D.; Finzi, A.C.; Pacala, S.W.; Burbank, D.H. Causes and Consequences of Resource Heterogeneity in Forests—Interspecific Variation in Light Transmission by Canopy Trees. Can. J. For. Res. 1994, 24, 337–349. [Google Scholar] [CrossRef]
- Pedlar, J.H.; Fahrig, L.; Merriam, H.G. Raccoon habitat use at 2 spatial scales. J. Wild. Manag. 1997, 61, 102–112. [Google Scholar] [CrossRef]
- Gysel, L.W. An ecological study of tree cavities and ground burrows in forest stands. J. Wild. Manag. 1961, 25, 12–20. [Google Scholar] [CrossRef]
- Korschgen, L.J. Foods of fox and gray squirrels in Missouri. J. Wild. Manag. 1981, 45, 260–266. [Google Scholar] [CrossRef]
- Brown, L.G.; Yeager, L.E. Fox Squirrels and Gray Squirrels in Illinois; Illinois Natural History Survey Bulletin: Urbana, IL, USA, 1945; Volume 23, pp. 449–533. [Google Scholar]
- Taylor, G.J. Present Status and Habitat Survey of the Delmarva Fox Squirrel (Sciurus niger cinereus) with a Discussion of Reasons for its Decline; University of Maryland: College Park, MD, USA, 1973. [Google Scholar]
- Nixon, C.M.; Havera, S.P.; Greenberg, R.E. Distribution and Abundance of the Gray Squirrel in Illinois; Department of Registration and Education, Natural History Survey Division: Urbana, IL, USA, 1978; number 105; pp. 1–56. [Google Scholar]
- Brown, B.W.; Batzli, G.O. Habitat selection by fox and gray squirrels: A multivariate analysis. J. Wild. Manag. 1984, 48, 616–621. [Google Scholar] [CrossRef]
- Loeb, S.C. The role of coarse woody debris in the ecology of southeastern mammals. In Biodiversity and coarse woody debris in southern forests: Effects on biodiversity. In Proceedings of the Workshop on Coarse Woody Debris in Southern Forests, Athens, GA, USA, 18–20 October; 1993; pp. 108–118. [Google Scholar]
- Petit, L.J.; Petit, D.R.; Martin, T.E. Landscape-Level Management of Migratory Birds—Looking Past the Trees to See the Forest. Wild. Soc. Bull. 1995, 23, 420–429. [Google Scholar]
- Askins, R.A. Sustaining biological diversity in early successional communities: The challenge of managing unpopular habitats. Wild. Soc. Bull. 2001, 29, 407–412. [Google Scholar]
- Shinder, B.; Steel, B.; List, P. Public judgements of adaptive management: A response from forest communities. J. For. 1996, 94, 4–12. [Google Scholar]
- Pease, B.S. Summer Habitat Use by a Mammal Community of an Oak-Dominated Ecosystem in the Central Hardwood Region; Southern Illinois University at Carbondale: Carbondale, IL, USA, 2017. [Google Scholar]
- Lashley, M.A.; Harper, C.A.; Bates, G.E.; Keyser, P.D. Forage Availability for White-Tailed Deer Following Silvicultural Treatments in Hardwood Forests. J. Wild. Manag. 2011, 75, 1467–1476. [Google Scholar] [CrossRef]
- Beier, P.; McCullough, D.R. Factors influencing white-tailed deer activity patterns and habitat use. Wild. Monogr. 1990, 3–51. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Hypothesis | Process | Covariate | Description |
|---|---|---|---|
| Forest composition | Occupancy | Maple importance | Site-specific importance value 1 for sugar maple (Acer saccharum) |
| Forest composition | Occupancy | Beech importance | Site-specific importance value for American beech (Fagus grandifolia) |
| Forest composition | Occupancy | Oak importance | Site-specific importance value for oak species (Quercus spp.) |
| Forest composition | Occupancy | Hickory importance | Site-specific importance value for hickory species (Carya spp.) |
| Forest composition | Occupancy | Yellow poplar importance | Site-specific importance value for yellow poplar (Liriodendron tulipifera) |
| Forest structure | Occupancy | Ground cover | Percentage ground (<1 m) vegetation cover |
| Forest structure | Occupancy | Seedling density | The number of seedlings per m−1 (woody stems up to 2.5 cm in diameter and at least 30.5 cm in height) |
| Forest structure | Occupancy | Sapling density | The number of woody stems per m−1 with a diameter of 2.5 to 7.6 cm |
| Forest structure | Occupancy | Coarse woody debris (CWD) | Site-specific volume of dead and downed trees, calculated using tree length and diameter at midpoint 2 |
| Forest structure | Occupancy | Overstory density | The number of trees >1 m in height and with a diameter of ≥7.6 cm per m−1 |
| Forest structure | Occupancy | Basal area | Tree area in square meters of the cross-section at breast height, presented as m2 ha−1 |
| Forest structure | Occupancy | Mean DBH | Site-specific mean diameter at breast height (dbh) |
| Temporal | Detection | Month | Month of camera survey |
| Temporal | Detection | Year | Year of camera survey |
| Temporal | Detection | Previous detection | A categorical indicator of whether a given mammal species was previously detected at a given site |
| Environmental | Detection | Precipitation | NCEP NARR 3 precipitation rate at the surface, averaged over a 1-week survey period |
| Environmental | Detection | Temperature | ECMWF 4 Interim Full Daily Surface Temperature (2 m above-ground), averaged over a 1-week survey period |
| Environmental | Detection | Topographic slope | Topographic slope at a camera-site; included to account for variation in slopes (10%–42%) across camera-sites as a potential factor in varying detection probabilities |
| Overstory | Understory | |||||||
|---|---|---|---|---|---|---|---|---|
| Importance Value 1 | Basal Area (m2 ha−1) | Density (Stems ha−1) | Density (Stems ha−1) | |||||
| Species | MEAN | SE | MEAN | SE | MEAN | SE | MEAN | SE |
| Overall | - | - | 24 | 0.5 | 406 | 23 | 947 | 67 |
| White oak (Quercus alba) | 26 | 1.7 | 7 | 0.5 | 61 | 6 | 2 | 2 |
| Sugar maple (Acer saccharum) | 16 | 1.5 | 2 | 0.2 | 95 | 12 | 171 | 24 |
| American beech (Fagus grandifolia) | 12 | 1.4 | 2 | 0.2 | 79 | 13 | 554 | 47 |
| Black oak (Quercus velutina) | 9 | 1.1 | 3 | 0.3 | 15 | 2 | 0 | 0 |
| Yellow poplar (Liriodendron tulipifera) | 7 | 1.1 | 2 | 0.3 | 20 | 6 | 8 | 4 |
| N. red oak (Quercus rubra) | 6 | 0.8 | 2 | 0.3 | 13 | 2 | 0 | 0 |
| Mockernut hickory (Carya tomentosa) | 5 | 0.8 | 1 | 0.2 | 29 | 7 | 10 | 5 |
| Sweetgum (Liquidambar styraciflua) | 3 | 0.6 | 1 | 0.2 | 14 | 4 | 8 | 5 |
| Bitternut hickory (Carya cordiformis) | 2 | 0.6 | 1 | 0.1 | 13 | 4 | 0 | 0 |
| Species | ||||
|---|---|---|---|---|
| Model Name | AIC a | ΔAIC b | Ω c | K d |
| White-tailed deer | ||||
| Null | 296.95 | 0.00 | 0.64 | 2 |
| Temporal | 298.69 | 1.74 | 0.27 | 5 |
| Environmental | 301.41 | 4.47 | 0.07 | 5 |
| Temporal + Environmental | 303.51 | 6.57 | 0.02 | 8 |
| Raccoon | ||||
| Null | 319.97 | 7.73 | 0.02 | 2 |
| Temporal | 318.13 | 6.16 | 0.04 | 5 |
| Environmental | 311.97 | 0.00 | 0.82 | 5 |
| Temporal + Environmental | 315.64 | 3.66 | 0.13 | 8 |
| Eastern gray squirrel | ||||
| Null | 173.13 | 7.36 | 0.02 | 2 |
| Temporal | 165.76 | 0.00 | 0.76 | 5 |
| Environmental | 174.46 | 8.70 | 0.01 | 5 |
| Temporal + Environmental | 168.37 | 2.61 | 0.21 | 8 |
| Species | |||||
|---|---|---|---|---|---|
| Hypothesis | Model Name | AIC a | ΔAIC b | ω c | K d |
| White-tailed deer | |||||
| Null | Null | 296.95 | 0.73 | 0.30 | 2 |
| Composition | Beech + Maple | 297.77 | 1.56 | 0.20 | 5 |
| Composition | Oak + Hickory | 296.21 | 0.00 | 0.43 | 5 |
| Structure | Understory | 300.49 | 4.28 | 0.05 | 6 |
| Structure | Overstory | 301.79 | 5.58 | 0.03 | 5 |
| Raccoon | |||||
| Null | Null | 311.97 | 0.00 | 0.62 | 5 |
| Composition | Beech + Maple | 315.16 | 3.18 | 0.13 | 8 |
| Composition | Oak + Hickory | 315.02 | 3.05 | 0.14 | 8 |
| Structure | Understory | 316.64 | 4.66 | 0.06 | 9 |
| Structure | Overstory | 316.66 | 4.69 | 0.06 | 8 |
| Eastern gray squirrel | |||||
| Null | Null | 174.46 | 4.09 | 0.05 | 5 |
| Composition | Beech + Maple | 170.37 | 0.00 | 0.35 | 8 |
| Composition | Oak + Hickory | 171.50 | 1.12 | 0.20 | 8 |
| Structure | Understory | 171.27 | 0.90 | 0.22 | 9 |
| Structure | Overstory | 171.55 | 1.18 | 0.19 | 8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pease, B.S.; Holzmueller, E.J.; Nielsen, C.K. Influence of Forest Structure and Composition on Summer Habitat Use of Wildlife in an Upland Hardwood Forest. Diversity 2019, 11, 160. https://doi.org/10.3390/d11090160
Pease BS, Holzmueller EJ, Nielsen CK. Influence of Forest Structure and Composition on Summer Habitat Use of Wildlife in an Upland Hardwood Forest. Diversity. 2019; 11(9):160. https://doi.org/10.3390/d11090160
Chicago/Turabian StylePease, Brent S., Eric J. Holzmueller, and Clayton K. Nielsen. 2019. "Influence of Forest Structure and Composition on Summer Habitat Use of Wildlife in an Upland Hardwood Forest" Diversity 11, no. 9: 160. https://doi.org/10.3390/d11090160
APA StylePease, B. S., Holzmueller, E. J., & Nielsen, C. K. (2019). Influence of Forest Structure and Composition on Summer Habitat Use of Wildlife in an Upland Hardwood Forest. Diversity, 11(9), 160. https://doi.org/10.3390/d11090160