The Evolution of Dragline Initiation in Spiders: Multiple Transitions from Multi- to Single-Gland Usage

Abstract
1. Introduction
- (1)
- The major ampullate glands are present in all araneomorphs. They produce the major component of the dragline and are the main subject of applied spider silk research. There are usually one to a few large major ampullate glands and their ducts lead to spigots on the inner margin of the ALS [8].
- (2)
- (3)
- (4)
- The piriform (sometimes spelled ‘pyriform’) glands produce glue-coated, fine fibres used to affix silk to substrates [14] or for thread joints during web building. There are usually a large number of small piriform glands, which are clustered on the ALS adjacent to the major ampullate spigots [7,8].
2. Material and Methods
2.1. Spider Collection
2.2. Observation of Spinning Behaviour
2.3. Phylogenetic Comparative Methods
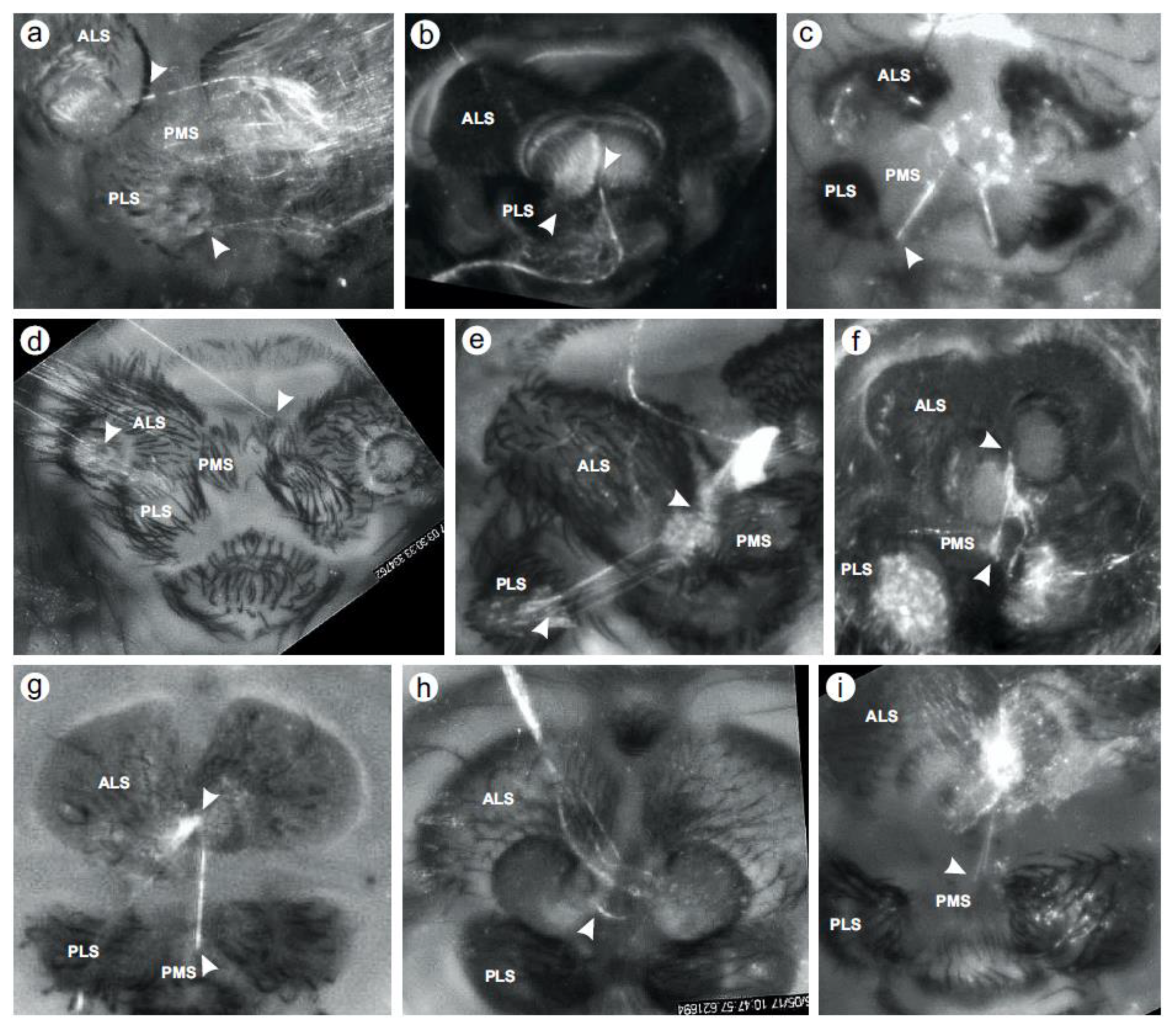
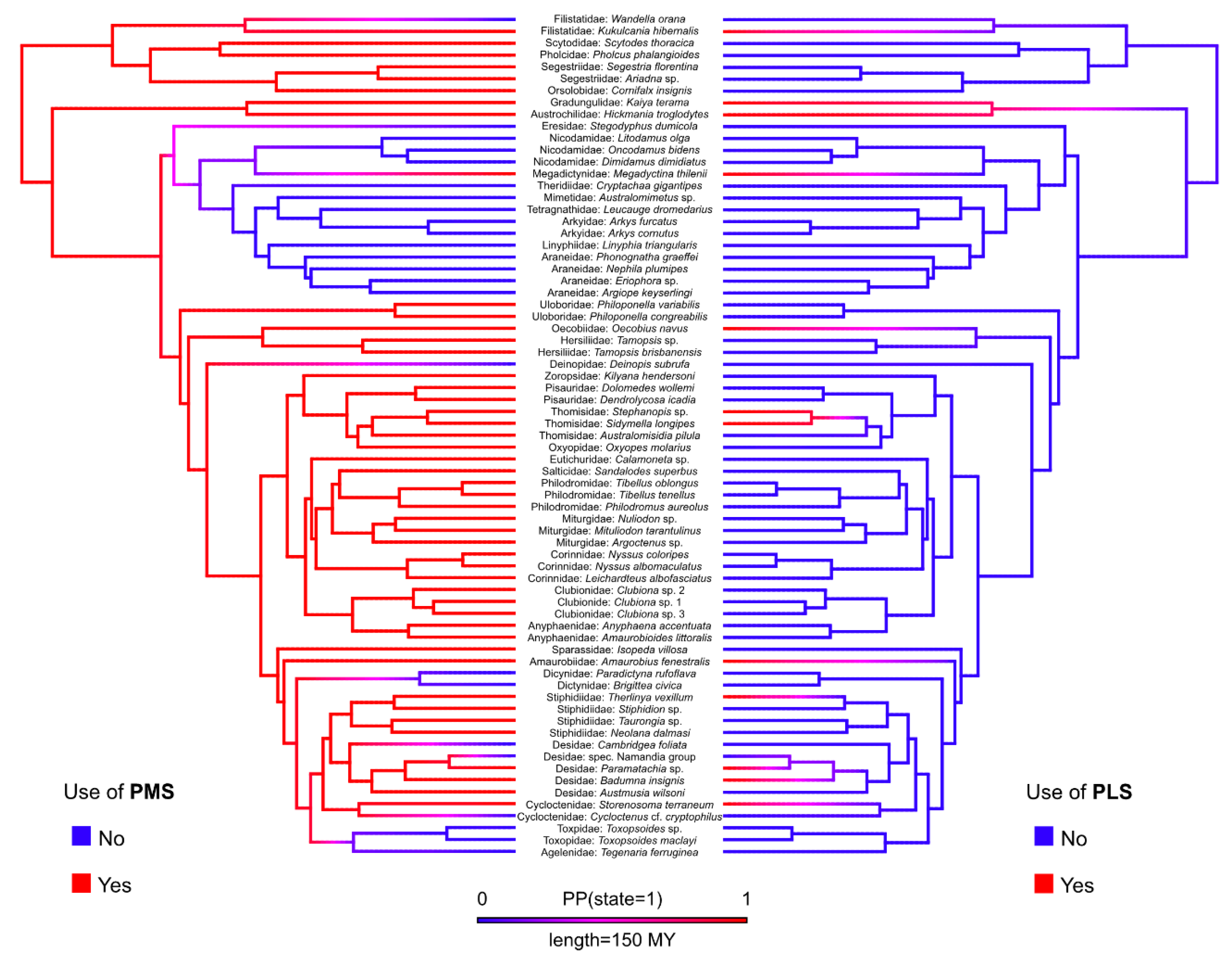
3. Results
4. Discussion
4.1. Most Draglines Are Compound Products
4.2. Evolutionary Trends
5. Conclusions and Outlook
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Vollrath, F.; Knight, D.P. Liquid crystalline spinning of spider silk. Nature 2001, 410, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.O.; Wells, D.; Reid, C.R.; Blamires, S.J. Clarity of objectives and working principles enhances the success of biomimetic programs. Bioinspir. Biomim. 2017, 12, 051001. [Google Scholar] [CrossRef] [PubMed]
- Vendrely, C.; Scheibel, T. Biotechnological production of spider-silk proteins enables new applications. Macromol. Biosci. 2007, 7, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Blamires, S.J. Biomechanical costs and benefits of sit-and-wait foraging traps. Isr. J. Ecol. Evol. 2019, 1, 1–10. [Google Scholar] [CrossRef]
- Blackledge, A.T.; Hayashi, C.Y. Silken toolkits: Biomechanics of silk fibers spun by the orb web spider Argiope argentata (Fabricius 1775). J. Exp. Biol. 2006, 209, 2452–2461. [Google Scholar] [CrossRef]
- Foelix, R.F. Biology of Spiders, 3rd ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2011; 419p. [Google Scholar]
- Apstein, C. Bau und Funktion der Spinndrüsen der Araneida. Arch. Naturg. 1889, 55, 29–74. [Google Scholar]
- Eberhard, W.G. Possible functional significance of spigot placement on the spinnerets of spiders. J. Arachnol. 2010, 38, 407–414. [Google Scholar] [CrossRef]
- Wolff, O.J.; Schneider, J.M.; Gorb, S.N. How to Pass the Gap-Functional Morphology and Biomechanics of Spider Bridging Threads. In Biotechnology of Silk; Asakura, T., Miller, T., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 165–177. [Google Scholar]
- Peters, H.M. On the structure and glandular origin of bridging lines used by spiders for moving to distant places. Act Zool Fenn 1990, 190, 309–314. [Google Scholar]
- Hayashi, Y.C.; Blackledge, T.A.; Lewis, R.V. Molecular and mechanical characterization of aciniform silk: Uniformity of iterated sequence modules in a novel member of the spider silk fibroin gene family. Mol. Biol. Evol. 2004, 21, 1950–1959. [Google Scholar] [CrossRef]
- Peakall, D.B. Composition, function and glandular origin of the silk fibroins of the spider Araneus diadematus Cl. J. Exp. Zool. Part A Ecol. Genet. Physiol. 1964, 156, 345–352. [Google Scholar] [CrossRef]
- Vasanthavada, K.; Hu, X.; Falick, A.M.; La Mattina, C.; Moore, A.M.F.; Jones, P.R.; Yee, R.; Reza, R.; Tuton, T.; Vierra, C. Aciniform spidroin, a constituent of egg case sacs and wrapping silk fibers from the black widow spider Latrodectus hesperus. J. Biol. Chem. 2007, 282, 35088–35097. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.O.; Grawe, I.; Wirth, M.; Karstedt, A.; Gorb, S.N. Spider’s super-glue: Thread anchors are composite adhesives with synergistic hierarchical organization. Soft Matter 2015, 11, 2394–2403. [Google Scholar] [CrossRef] [PubMed]
- Blackledge, T.A.; Pérez-Rigueiro, J.; Plaza, G.R.; Perea, B.; Navarro, A.; Guinea, G.V.; Elices, M.S. Sequential origin in the high performance properties of orb spider dragline silk. Sci. Rep. 2012, 2, 782. [Google Scholar] [CrossRef] [PubMed]
- Kovoor, J. Comparative structure and histochemistry of silk-producing organs in arachnids. In Ecophysiology of Spiders; Springer: Berlin, Germany, 1987; pp. 160–186. [Google Scholar]
- Řezáč, M.; Krejčí, T.; Goodacre, S.L.; Haddad, C.R.; Řezáčová, V. Morphological and functional diversity of minor ampullate glands in spiders from the superfamily Amaurobioidea (Entelegynae: RTA clade). J. Arachnol. 2017, 45, 198–209. [Google Scholar] [CrossRef]
- Wolff, J.O. Attachment Structures and Adhesive Secretions in Arachnids. Biologically-Inspired Systems; Gorb, S.N., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 7. [Google Scholar]
- Wolff, J.O.; Řezáč, M.; Krejčí, T.; Gorb, S.N. Hunting with sticky tape: Functional shift in silk glands of araneophagous ground spiders (Gnaphosidae). J. Exp. Biol. 2017, 220, 2250–2259. [Google Scholar] [CrossRef] [PubMed]
- Wolff, O.J.; Herberstein, M.E. 3D-printing spiders: Back-and-forth glue application yields silk anchorages with high pull-off resistance under varying loading situations. J. R. Soc. Interface 2017, 14, 20160783. [Google Scholar] [CrossRef]
- Wolff, J.O.; Paterno, G.B.; Liprandi, D.; Ramírez, M.J.; Bosia, F.; Van Der Meijden, A.; Michalik, P.; Smith, H.M.; Jones, B.R.; Ravelo, A.M.; et al. Evolution of aerial spider webs coincided with repeated structural optimization of silk anchorages. Evol. Dev. 2019, 73, 2122–2134. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, N.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2016. Available online: https://www.R-project.org/ (accessed on 17 December 2019).
- Paterno, G.B.; Penone, C.; Werner, G.D. sensiPhy: An r-package for sensitivity analysis in phylogenetic comparative methods. Methods Ecol. Evol. 2018, 9, 1461–1467. [Google Scholar] [CrossRef]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Huelsenbeck, P.J.; Nielsen, R.; Bollback, J.P. Stochastic mapping of morphological characters. Syst. Biol. 2003, 52, 131–158. [Google Scholar] [CrossRef] [PubMed]
- Work, R.W. Web components associated with the major ampullate silk fibers of orb-web-building spiders. Trans. Am. Microsc. Soc. 1981, 100, 1–20. [Google Scholar] [CrossRef]
- Braunitzer, G.; Wolff, D. Vergleichende chemische Untersuchungen über die Fibroine von Bombyx mori und Nephila madagascariensis. Z. Für Nat. B 1955, 10, 404–408. [Google Scholar] [CrossRef]
- Work, R.W. Mechanisms of major ampullate silk fiber formation by orb-web-spinning spiders. Trans. Am. Microsc. Soc. 1977, 96, 170–189. [Google Scholar] [CrossRef]
- Peters, M.H.; Kovoor, J. The silk-producing system ofLinyphia triangularis (Araneae, Linyphiidae) and some comparisons with Araneidae. Zoomorphology 1991, 111, 1–17. [Google Scholar] [CrossRef]
- Swanson, B.O.; Blackledge, T.A.; Beltrán, J.; Hayashi, C.Y. Variation in the material properties of spider dragline silk across species. Appl. Phys. A 2006, 82, 213–218. [Google Scholar] [CrossRef]
- Blackledge, T.A. Spider silk: A brief review and prospectus on research linking biomechanics and ecology in draglines and orb webs. J. Arachnol. 2012, 40, 1–13. [Google Scholar] [CrossRef]
- Madurga, R.; Plaza, G.R.; Blackledge, T.A.; Guinea, G.; Elices, M.; Pérez-Rigueiro, J. Material properties of evolutionary diverse spider silks described by variation in a single structural parameter. Sci. Rep. 2016, 6, 18991. [Google Scholar] [CrossRef]
- Lopardo, L.; Ramirez, M.J. The combing of cribellar silk by the prithine Misionella mendensis, with notes on other filistatid spiders (Araneae: Filistatidae). Am. Mus. Novit. 2007, 3563, 1–14. [Google Scholar] [CrossRef]
- Blamires, S.J.; Blackledge, T.A.; Tso, I.M. Physicochemical property variation in spider silk: Ecology, evolution, and synthetic production. Annu. Rev. Entomol. 2017, 62, 443–460. [Google Scholar] [CrossRef]
- Patek, S. Biomimetics and evolution. Science 2014, 345, 1448–1449. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Family | Species | Individuals Studied | Number of Observations | PMS Used | PLS Used |
|---|---|---|---|---|---|
| Agelenidae | Tegenaria ferruginea (Panzer, 1804) | 3 juv | 26 | − | − |
| Amaurobiidae | Amaurobius fenestralis (Ström, 1768) | 2 f, 1 juv | 15 | + | + |
| (to Cycloctenidae) | Storenosoma sp. | 1 juv | 6 | + | + |
| (to Cycloctenidae) | Storenosoma terraneum Davies, 1986 | 1 juv | 4 | + | + |
| Anyphaenidae | Amaurobioides litoralis Hickman, 1949 | 3 juv | 34 | + | − |
| Anyphaena accentuata (Walckenaer, 1802) | 2 juv | 16 | + | − | |
| Araneidae | Argiope keyserlingi Karsch, 1878 | 2 f, 1 juv | 21 | − | − |
| Cyrtophora hirta L. Koch, 1872 | 2 f, 1 juv | 24 | − | − | |
| Eriophora sp. | 1 f, 3 juv | 24 | − | − | |
| Nephila plumipes (Latreille, 1804) | 3 f, 1 juv | 15 | − | − | |
| Phonognatha graeffei (Keyserling, 1865) | 3 f | 28 | − | − | |
| Arkyidae | Arkys cornutus L. Koch, 1872 | 1 f | 9 | − | − |
| Arkys furcatus (Balogh, 1978) | 1 m | 7 | − | − | |
| Austrochilidae | Hickmania troglodytes (Higgins & Petterd, 1883) | 2 f, 1 m | 31 | + | + |
| Clubionidae | Clubiona sp. 1 | 1 m | 10 | + | − |
| Clubiona sp. 2 | 1 f | 5 | + | − | |
| Clubiona sp. 3 | 1 f | 8 | + | − | |
| Corinnidae | Leichhardteus albofasciatus Baehr & Raven, 2013 | 1 f, 1 juv | 18 | + | − |
| Nyssus coloripes Walckenaer, 1805 | 1 f, 1 m | 15 | + | − | |
| Nyssus albopunctatus (Hogg, 1896) | 1 f, 1 juv | 19 | + | − | |
| Cycloctenidae | Cycloctenus cf. crytophilus Hickman, 1981 | 1 f | 7 | − | − |
| Deinopidae | Deinopis subrufa L. Koch, 1879 | 2 f, 1 m | 34 | − | − |
| Desidae | Austmusia wilsoni Gray, 1983 | 3 juv | 15 | + | − |
| Badumna insignis (L. Koch, 1872) | 2 f, 2 juv | 33 | + | + | |
| Cambridgea foliata (L. Koch, 1872) | 2 f, 1 juv | 22 | − | − | |
| Paramatachia sp. 1 | 1 f | 7 | + | + | |
| Paramatachia sp. 2 | 1 f | 10 | + | + | |
| spec. Namandia group | 1 f, 1 m, 1 juv | 21 | − | − | |
| (to Stiphidiidae) | Taurongia sp. | 1 f | 7 | + | − |
| Dictynidae | Brigittea civica (Lucas, 1850) | 3 f | 27 | − | − |
| Paradictyna rufoflava (Chamberlain, 1946) | 3 f | 23 | − | − | |
| Eresidae | Stygodyphus dumicola Pocock, 1898 | 1 f, 3 juv | 37 | − | − |
| Eutichuridae | Calamoneta sp. | 3 juv | 32 | + | − |
| Filistatidae | Kulkulcania hibernalis (Hentz, 1842) | 3 juv | 24 | + | + |
| Wandella orana Gray, 1994 | 3 f | 24 | − | − | |
| Gradungulidae | Kaiya terama Gray, 1987 | 3 juv | 18 | + | + |
| Hersiliidae | Tamopsis brisbanensis Baehr & Baehr, 1987 | 1 f | 8 | + | − |
| Tamopsis sp. 2 | 2 juv | 20 | + | − | |
| Tamopsis sp. 3 | 1 juv | 8 | + | − | |
| Linyphiidae | Linyphia triangularis (Clerck, 1757) | 1 f, 1 m, 1 juv | 22 | − | − |
| Megadictynidae | Megadictyna thilenii Dahl, 1906 | 1 f, 1 m, 2 juv | 29 | + | + |
| Mimetidae | Australomimetus sp. | 3 f | 26 | − | − |
| Miturgidae | Argoctenus sp. | 1 m | 6 | + | − |
| Mituliodon tarantulinus (L. Koch, 1873) | 1 f, 1 m, 1 juv | 25 | + | − | |
| Nuliodon sp. | 3 f | 18 | + | − | |
| Nicodamidae | Dimidamus dimidiatus (Simon, 1897) | 1 f, 1 m | 18 | − | − |
| Litodamus olga Harvey, 1995 | 1 f | 7 | − | − | |
| Oncodamus bidens (Karsch, 1878) | 2 f, 1 juv | 33 | − | − | |
| Oecobiidae | Oecobius navus Blackwall, 1859 | 4 f | 30 | + | + |
| Orsolobidae | Cornifalx insignis Hickman, 1979 | 1 f | 6 | + | − |
| Oxyopidae | Oxyopes molarius L. Koch, 1878 | 2 f, 1 m | 21 | + | − |
| Philodromidae | Philodromus aureolus (Clerck, 1757) | 3 f | 25 | + | − |
| Tibellus tenellus (L. Koch, 1876) | 1 f | 9 | + | − | |
| Pholcidae | Pholcus phalangioides (Fuesslin, 1775) | 3 f | 27 | + | − |
| Pisauridae | Dolomedes wollemi Raven & Hebron, 2018 | 3 juv | 20 | + | − |
| Dendrolycosa icadia (L. Koch, 1876) | 2 m, 1 juv | 23 | + | − | |
| Salticidae | Sandalodes superbus (Karsch, 1878) | 2 f, 1 juv | 30 | + | − |
| Scytodidae | Scytodes thoracica (Latreille, 1802) | 1 f | 10 | + | − |
| Segestriidae | Ariadna sp. 1 | 1 juv | 8 | + | − |
| Ariadna sp. 2 | 1 juv | 10 | + | − | |
| Gippsicola sp. | 1 f | 8 | + | − | |
| Segestria florentina (Rossi, 1790) | 3 f | 27 | + | − | |
| Sparassidae | Isopeda villosa (L. Koch, 1875 | 1 f, 2 juv | 23 | + | − |
| Stiphidiidae | Neolana dalmasi (Marples, 1959) | 2 f | 21 | + | − |
| Stiphidion sp. | 1 f | 9 | + | − | |
| Therlinya vexillum Gray & Smith, 2002 | 3 f | 26 | + | + | |
| Tetragnathidae | Leucauge dromedaria (Thorell, 1881) | 3 f | 28 | − | − |
| Thomisidae | Australomisidia pilula (L. Koch, 1867) | 2 f, 1 juv | 26 | + | − |
| Sidymella longipes (L. Koch, 1874) | 1 f | 8 | + | + | |
| Stephanopis sp. 1 | 1 f | 7 | + | + | |
| Stephanopis sp. 2 | 1 juv | 8 | + | + | |
| Theridiidae | Cryptachaea gigantipes (Keyserling, 1890) | 3 f | 22 | − | − |
| Toxopidae | Toxopsoides sp. | 3 m | 25 | − | − |
| Toxopsoides macleayi Smith, 2013 | 2 m, 1 juv | 20 | − | − | |
| Uloboridae | Philoponella congregabilis (Rainbow, 1916) | 3 f | 23 | + | − |
| Philoponella variabilis (Keyserling, 1887) | 4 f, 1 juv | 28 | + | − | |
| Zoropsidae | Kilyana cf. hendersoni Raven & Stumkat, 2005 | 1 f, 1 m, 1 juv | 20 | + | − |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolff, J.O. The Evolution of Dragline Initiation in Spiders: Multiple Transitions from Multi- to Single-Gland Usage. Diversity 2020, 12, 4. https://doi.org/10.3390/d12010004
Wolff JO. The Evolution of Dragline Initiation in Spiders: Multiple Transitions from Multi- to Single-Gland Usage. Diversity. 2020; 12(1):4. https://doi.org/10.3390/d12010004
Chicago/Turabian StyleWolff, Jonas O. 2020. "The Evolution of Dragline Initiation in Spiders: Multiple Transitions from Multi- to Single-Gland Usage" Diversity 12, no. 1: 4. https://doi.org/10.3390/d12010004
APA StyleWolff, J. O. (2020). The Evolution of Dragline Initiation in Spiders: Multiple Transitions from Multi- to Single-Gland Usage. Diversity, 12(1), 4. https://doi.org/10.3390/d12010004