A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads


Abstract
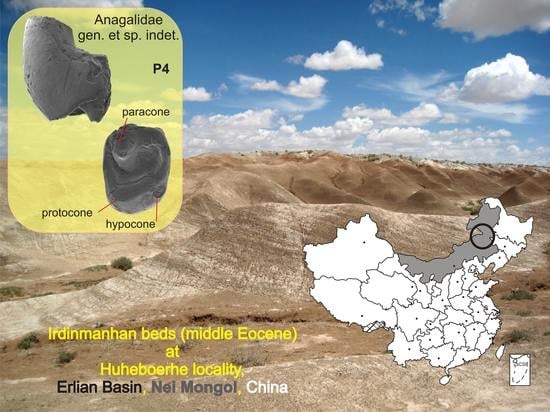
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
3. Systematic Paleontology
- Mammalia Linnaeus, 1758
- Euarchontoglires Murphy et al., 2001
- Anagalida Szalay and McKenna, 1971
- Anagalidae Simpson, 1931
- Gen. et sp. indet.
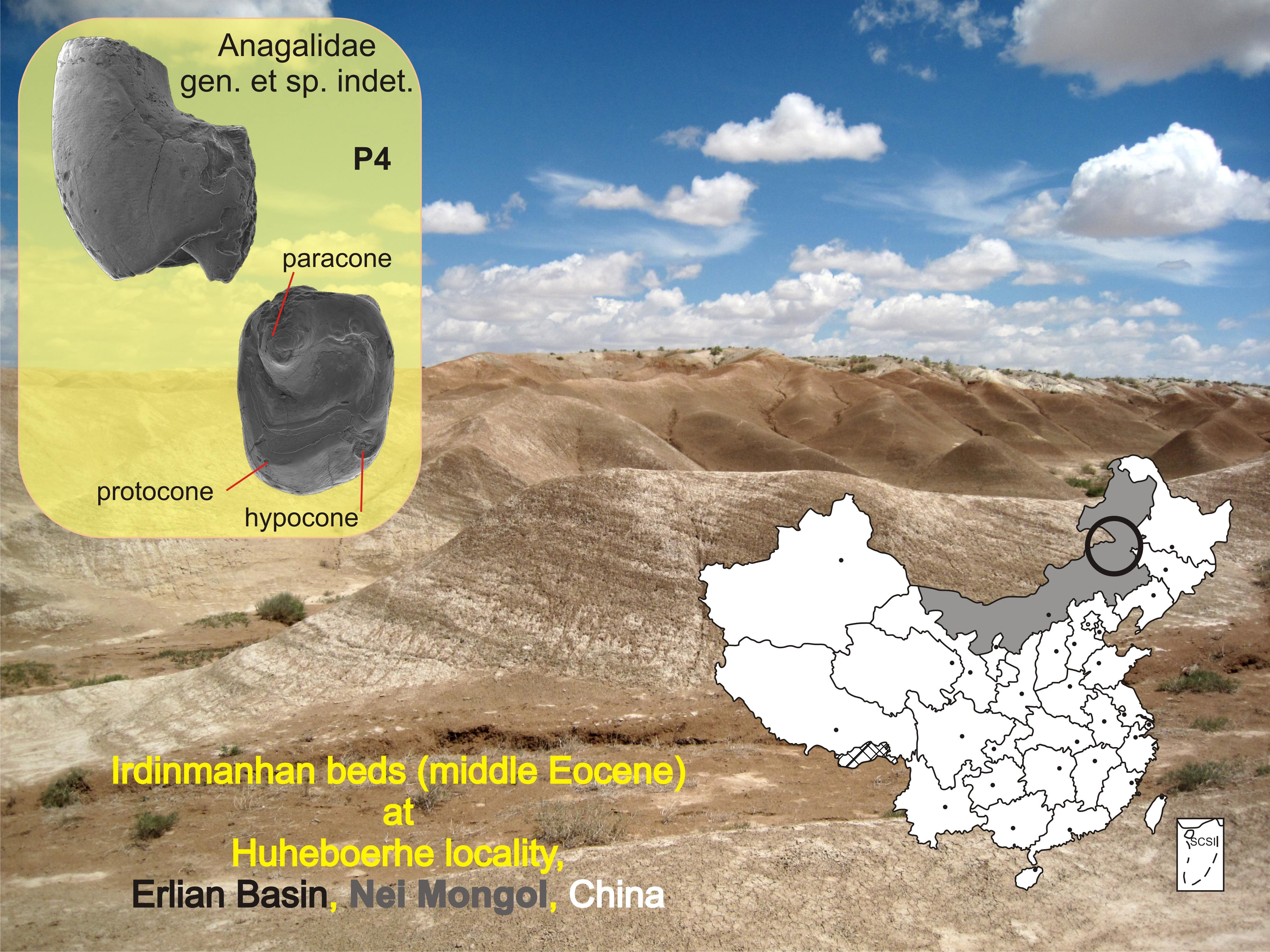
- (Figure 2)
Description and Comparisons
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murphy, W.J.; Eizirik, E.; Johnson, W.E.; Zhang, Y.P.; Ryder, O.A.; O’Brien, S.J. Molecular phylogenetics and the origins of placental mammals. Nature 2001, 409, 614–618. [Google Scholar] [CrossRef]
- Asher, R.J.; Bennett, N.; Lehmann, T. The new framework for understanding placental mammal evolution. Bioessays 2009, 31, 853–864. [Google Scholar] [CrossRef]
- Hautier, L.; Bennett, N.C.; Viljoen, H.; Howard, L.; Milinkovitch, M.C.; Tzika, A.C.; Goswami, A.; Asher, R.J. Patterns of ossification in southern versus northern placental mammals. Evolution 2013, 67, 1994–2010. [Google Scholar] [CrossRef]
- Asher, R.J.; Meng, J.; Wible, J.R.; McKenna, M.C.; Rougier, G.W.; Novacek, M.J. Stem Lagomorpha and the antiquity of Glires. Science 2005, 307, 1091–1094. [Google Scholar] [CrossRef]
- Fostowicz-Frelik, Ł. Convergent and parallel evolution in early Glires (Mammalia). In Evolutionary Biology: Self/Nonself Evolution, Species and Complex Traits Evolution, Methods and Concepts; Pontarotti, P., Ed.; Springer: Cham, Switzerland, 2017; pp. 199–216. [Google Scholar] [CrossRef]
- Fostowicz-Frelik, Ł. Most successful mammals in the making: A review of the Paleocene Glires. In Evolutionary Biology: A Transdisciplinary Approach; Pontarotti, P., Ed.; Springe: Cham, Switzerland, 2020; in press. [Google Scholar] [CrossRef]
- Meng, J.; Hu, Y.M.; Li, C.K. The osteology of Rhombomylus (Mammalia, Glires): Implications for phylogeny and evolution of Glires. Bull. Am. Mus. Nat. Hist. 2003, 275, 1–247. [Google Scholar] [CrossRef]
- Szalay, F.S.; McKenna, M.C. Beginning of the age of mammals in Asia: The late Paleocene Gashato fauna, Mongolia. Bull. Am. Mus. Nat. Hist. 1971, 144, 273–317. [Google Scholar]
- McKenna, M.C. Toward a phylogenetic classification of Mammalia. In Phylogeny of the Primates. A Multidisciplinary Approach; Luckett, W.P., Szalay, F.S., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 21–46. [Google Scholar]
- Murphy, W.J.; Pringle, T.H.; Crider, T.A.; Springer, M.S.; Miller, W. Using genomic data to unravel the root of the placental mammal phylogeny. Genome Res. 2007, 17, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Wyss, A.R. The morphology of Tribosphenomys (Rodentiaformes, Mammalia): Phylogenetic implications for basal Glires. J. Mammal. Evol. 2001, 8, 1–71. [Google Scholar] [CrossRef]
- Missiaen, P.; Smith, T. The Gashatan (late Paleocene) mammal fauna from Subeng, Inner Mongolia, China. Acta Palaeontol. Pol. 2008, 53, 357–378. [Google Scholar] [CrossRef]
- López-Torres, S.; Fostowicz-Frelik, Ł. A new Eocene anagalid (Mammalia: Euarchontoglires) from Mongolia and its implications for the group’s phylogeny and dispersal. Sci. Rep. 2018, 8, e13955. [Google Scholar] [CrossRef] [PubMed]
- Flynn, L.J. Hystricognathi and Rodentia incertae sedis. In Evolution of Tertiary Mammals in North America. Volume 2: Small Mammals, Xenarthrans, and Marine Mammals; Janis, C.M., Gunnell, G.F., Uhen, M.D., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 498–506. [Google Scholar]
- Li, C.K. Order Anagalida. In Basal Synapsids and Mammals, Volume III, Eulipotyphlans, Proteutheres, Chiropterans, Euarchontans, and Anagalids; Li, C.K., Qiu, Z., Eds.; Science Press: Beijing, China, 2016; pp. 390–411. [Google Scholar]
- Simpson, G.G. A new insectivore from the Oligocene, Ulan Gochu Horizon, of Mongolia. Am. Mus. Novit. 1931, 505, 1–22. [Google Scholar]
- Bohlin, B. Some mammalian remains from Shih-ehr-ma-ch’eng, Hui-hui-p’u area, Western Kansu. Reports from the Scientific Expedition to the North-Western Provinces of China under Leadership of Dr Sven Hedin. The Sino-Swedish Expedition Publication 35, VI. Vert. Paleontol. 1951, 5, 1–48. [Google Scholar]
- McKenna, M.C. New evidence against tupaioid affinities of the mammalian family Anagalidae. Am. Mus. Novit. 1963, 2158, 1–16. [Google Scholar]
- Bai, B.; Wang, Y.Q.; Li, Q.; Wang, H.B.; Mao, F.Y.; Gong, Y.X.; Meng, J. Biostratigraphy and diversity of Paleogene perissodactyls from the Erlian Basin of Inner Mongolia, China. Am. Mus. Novit. 2018, 3914, 1–60. [Google Scholar] [CrossRef]
- Russell, D.E.; Zhai, R.J. The Paleogene of Asia: Mammals and stratigraphy. Mém. Mus. Natl. Hist. Nat. Série C Sci. Terre. 1987, 52, 1–488. [Google Scholar]
- Wang, Y.Q.; Meng, J.; Beard, K.C.; Li, Q.; Ni, X.; Gebo, D.L.; Bai, B.; Jin, X.; Li, P. Early Paleogene stratigraphic sequences, mammalian evolution and its response to environmental changes in Erlian Basin, Inner Mongolia, China. Sci. China Earth Sci. 2010, 53, 1918–1926. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Meng, J.; Jin, X. Comments on Paleogene localities and stratigraphy in the Erlian Basin, Nei Mongol, China. Vert. Palasiat. 2012, 50, 181–203. [Google Scholar]
- Meng, J.; Wang, Y.; Ni, X.; Beard, K.C.; Sun, C.; Li, Q.; Jin, X.; Bai, B. New stratigraphic data from the Erlian Basin: Implications for the division, correlation, and definition of Paleogene lithological units in Nei Mongol (Inner Mongolia). Am. Mus. Novit. 2007, 3570, 1–31. [Google Scholar] [CrossRef]
- Fostowicz-Frelik, Ł.; Li, C.K.; Mao, F.Y.; Meng, J.; Wang, Y.Q. A large mimotonid from the Middle Eocene of China sheds light on the evolution of lagomorphs and their kin. Sci. Rep. 2015, 5, 9394. [Google Scholar] [CrossRef]
- Meng, J.; Bowen, G.J.; Ye, J.; Koch, P.L.; Ting, S.Y.; Li, Q.; Jin, X. Gomphos elkema (Glires, Mammalia) from the Erlian Basin: Evidence for the Early Tertiary Bumbanian Land Mammal Age in Nei-Mongol, China. Am. Mus. Novit. 2004, 3425, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Fostowicz-Frelik, Ł.; Li, Q. A new genus of stem lagomorphs (Mammalia: Glires) from the Middle Eocene of the Erlian Basin, Nei Mongol, China. Acta Zool. Cracov. 2014, 57, 29–42. [Google Scholar] [CrossRef]
- Li, Q.; Meng, J. New ctenodactyloid rodents from the Erlian Basin, Nei Mongol, China and the phylogenic relationships of Eocene Asian ctenodactyloids. Am. Mus. Novit. 2015, 3828, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Li, Q. Eocene fossil rodent assemblages from the Erlian Basin (Inner Mongolia, China): Biochronological implications. Palaeoworld 2016, 25, 95–103. [Google Scholar] [CrossRef]
- Dashzeveg, D.; Russell, D.E. Palaeocene and Eocene Mixodontia (Mammalia, Glires) of Mongolia and China. Palaeontology 1988, 31, 129–164. [Google Scholar]
- Li, Q.; Wang, Y.Q.; Fostowicz-Frelik, Ł. Small mammal fauna from Wulanhuxiu (Nei Mongol, China) implies faunal turnover across the Irdinmanhan–Sharamurunian boundary. Acta Palaeontol. Pol. 2016, 61, 759–776. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; McKenna, M.C. Faunal turnovers of Palaeogene mammals from the Mongolian Plateau. Nature 1998, 394, 364–367. [Google Scholar] [CrossRef]
- Sun, J.; Ni, X.; Bi, S.; Wu, W.; Ye, J.; Meng, J.; Windley, B.F. Synchronous turnover of flora, fauna, and climate at the Eocene-Oligocene boundary in Asia. Sci. Rep. 2014, 4, 7463. [Google Scholar] [CrossRef] [Green Version]
- Li, J.X.; Yue, L.P.; Roberts, A.P.; Hirt, A.M.; Pan, F.; Guo, L.; Xu, Y.; Xi, R.G.; Guo, L.; Qiang, X.K. Global cooling and enhanced Eocene Asian mid-latitude interior aridity. Nat. Commun. 2018, 9, 3026. [Google Scholar] [CrossRef]
- Wasiljeff, J.; Kaakinen, A.; Salminen, J.N.; Zhang, Z.Q. Magnetostratigraphic constraints on the fossiliferous Ulantatal sequence in Inner Mongolia, China: Implications for Asian aridification and faunal turnover before the Eocene-Oligocene boundary. Earth Planet. Sci. Lett. 2020, 535, 116125. [Google Scholar] [CrossRef]
- Li, Q.; Meng, J. Eocene ischyromyids (Rodentia, Mammalia) from the Erlian Basin, Nei Mongol, China. Vert. Palasiat. 2013, 51, 289–304. [Google Scholar]
- Ni, X.; Li, Q.; Li, L.; Beard, K.C. Oligocene primates from China reveal divergence between African and Asian primate evolution. Science 2016, 352, 673–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Ni, X. An early Oligocene fossil demonstrates treeshrews are slowly evolving “living fossils”. Sci. Rep. 2016, 6, 18627. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Q.; Wang, J. On the geological age of mammalian fossils from Shanmacheng, Gansu Province. Vert. Palasiat. 2016, 54, 351–357. [Google Scholar]
- Seo, H.; Kim, J.; Hwang, J.J.; Jeong, H.G.; Han, S.S.; Park, W.; Ryu, K.; Seomun, H.; Kim, J.Y.; Cho, E.S.; et al. Regulation of root patterns in mammalian teeth. Sci. Rep. 2017, 7, 12714. [Google Scholar] [CrossRef] [Green Version]
- Fostowicz-Frelik, Ł.; Li, C.K.; Meng, J.; Wang, Y.Q. New Gobiolagus (Mammalia: Lagomorpha) material from the Middle Eocene of Erden Obo (Nei Mongol, China). Vert. Palasiat. 2012, 50, 219–236. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fostowicz-Frelik, Ł.; Li, Q.; Saha, A. A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads. Diversity 2020, 12, 420. https://doi.org/10.3390/d12110420
Fostowicz-Frelik Ł, Li Q, Saha A. A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads. Diversity. 2020; 12(11):420. https://doi.org/10.3390/d12110420
Chicago/Turabian StyleFostowicz-Frelik, Łucja, Qian Li, and Anwesha Saha. 2020. "A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads" Diversity 12, no. 11: 420. https://doi.org/10.3390/d12110420
APA StyleFostowicz-Frelik, Ł., Li, Q., & Saha, A. (2020). A Gliriform Tooth from the Eocene of the Erlian Basin (Nei Mongol, China) and the Premolar Morphology of Anagalidan Mammals at a Crossroads. Diversity, 12(11), 420. https://doi.org/10.3390/d12110420