First Complete Wing of a Stem Group Sphenisciform from the Paleocene of New Zealand Sheds Light on the Evolution of the Penguin Flipper

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
3. Systematic Paleontology
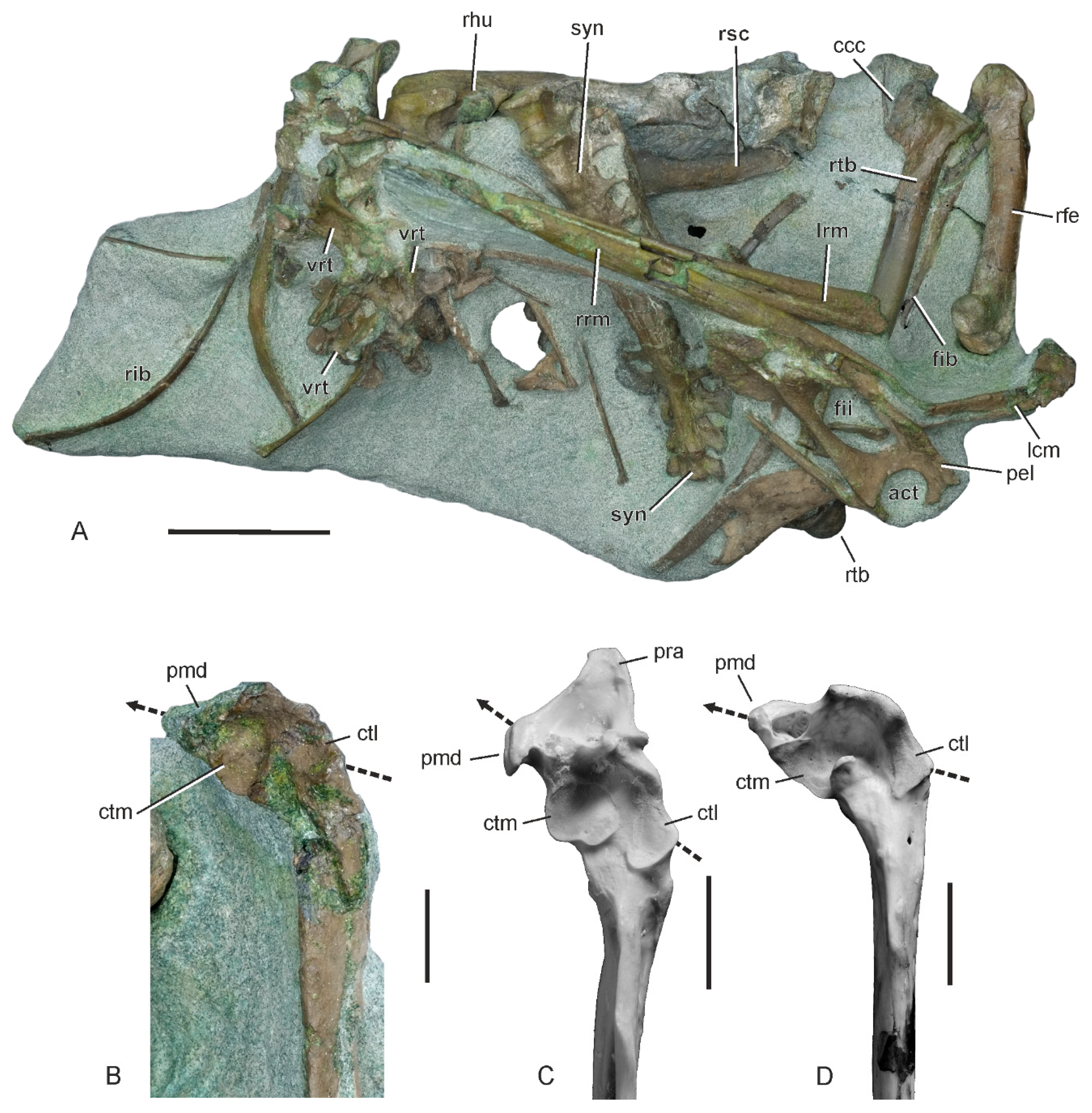
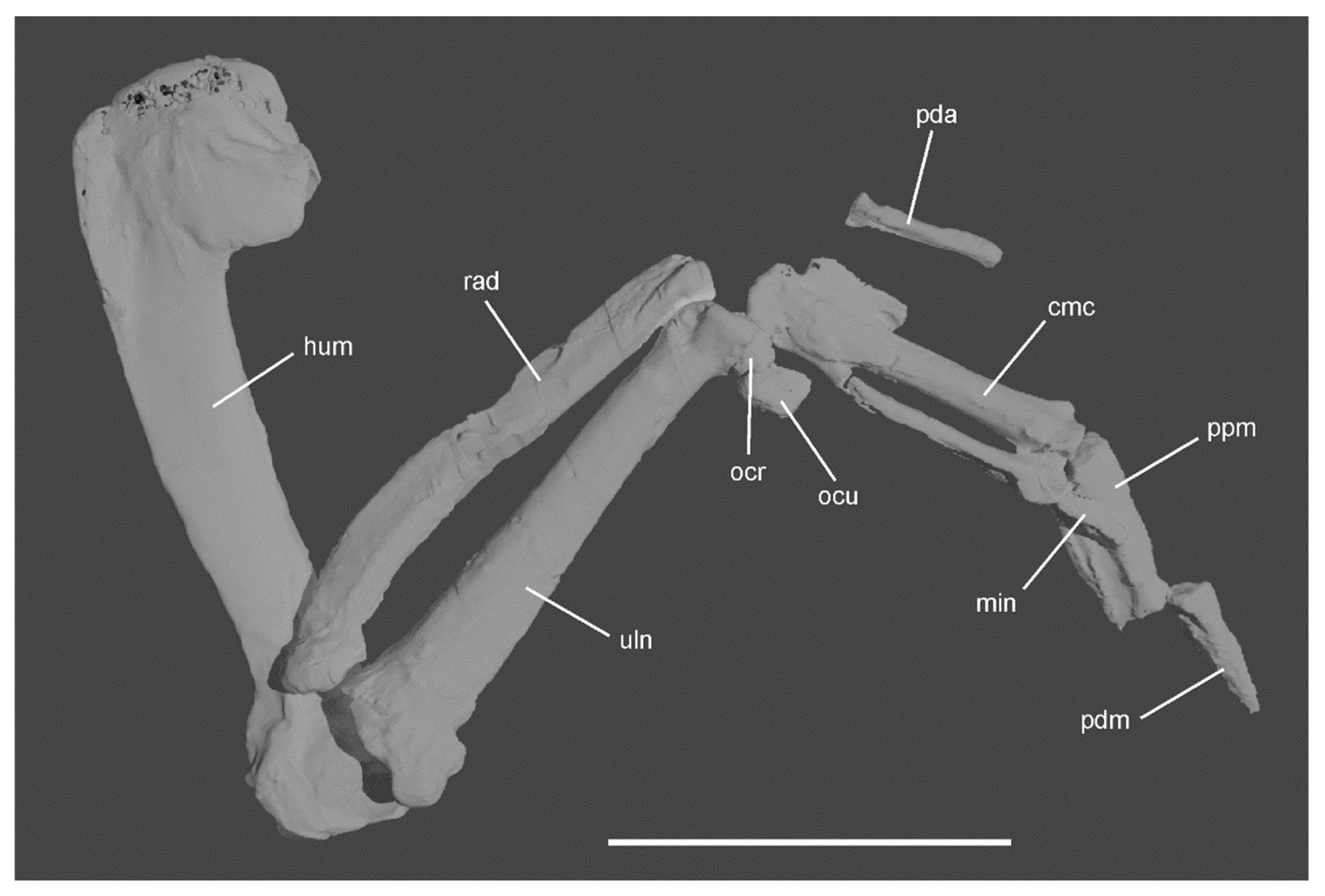
3.1. Referred Specimen
3.2. Locality and Horizon
3.3. Measurements (in mm, in Brackets the Dimensions of other Muriwaimanu tuatahi Specimens)
3.4. Taxonomic Remarks
3.5. Description and Comparisons
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Raikow, R.J.; Bicanovsky, L.; Bledsoe, A.H. Forelimb joint mobility and the evolution of wing-propelled diving in birds. Auk 1988, 105, 446–451. [Google Scholar] [CrossRef]
- Schreiweis, D.O. A comparative study of the appendicular musculature of penguins (Aves, Sphenisciformes). Smithson. Contrib. Zool. 1982, 341, 1–46. [Google Scholar] [CrossRef]
- Ksepka, D.T.; Ando, T. Penguins past, present, and future: Trends in the evolution of the Sphenisciformes. In Living Dinosaurs. The Evolutionary History of Modern Birds; Dyke, G., Kaiser, G., Eds.; John Wiley and Sons: Chichester, UK, 2011; pp. 155–186. [Google Scholar]
- Mayr, G. Avian Evolution: The Fossil Record of Birds and Its Paleobiological Significance; Wiley-Blackwell: Chichester, UK, 2017; pp. 1–293. [Google Scholar]
- Slack, K.E.; Jones, C.M.; Ando, T.; Harrison, G.L.; Fordyce, R.E.; Arnason, U.; Penny, D. Early penguin fossils, plus mitochondrial genomes, calibrate avian evolution. Mol. Biol. Evol. 2006, 23, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Mayr, G.; De Pietri, V.L.; Love, L.; Mannering, A.A.; Scofield, R.P. A well-preserved new mid-Paleocene penguin (Aves, Sphenisciformes) from the Waipara Greensand in New Zealand. J. Vertebr. Paleontol. 2017, 37, e139816915. [Google Scholar] [CrossRef]
- Mayr, G.; De Pietri, V.L.; Scofield, R.P. A new fossil from the mid-Paleocene of New Zealand reveals an unexpected diversity of world’s oldest penguins. Sci. Nat. 2017, 104, 9. [Google Scholar] [CrossRef] [PubMed]
- Mayr, G.; De Pietri, V.L.; Love, L.; Mannering, A.A.; Scofield, R.P. Leg bones of a new penguin species from the Waipara Greensand add to the diversity of very large-sized Sphenisciformes in the Paleocene of New Zealand. Alcheringa 2020. [Google Scholar] [CrossRef]
- Blokland, J.C.; Reid, C.M.; Worthy, T.H.; Tennyson, A.J.D.; Clarke, J.A.; Scofield, R.P. Chatham Island Paleocene fossils provide insight into the palaeobiology, evolution, and diversity of early penguins (Aves, Sphenisciformes). Palaeontol. Electron. 2019, 22, 1–92. [Google Scholar] [CrossRef]
- Jadwiszczak, P. Partial limb skeleton of a “giant penguin” Anthropornis from the Eocene of Antarctic Peninsula. Pol. Polar Res. 2012, 33, 259–274. [Google Scholar] [CrossRef]
- Clarke, J.A.; Ksepka, D.T.; Stucchi, M.; Urbina, M.; Giannini, N.; Bertelli, S.; Narváez, Y.; Boyd, C.A. Paleogene equatorial penguins challenge the proposed relationship between penguin biogeography, diversity, and Cenozoic climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 11545–11550. [Google Scholar] [CrossRef]
- Ksepka, D.T.; Clarke, J.A.; de Vries, T.J.; Urbina, M. Osteology of Icadyptes salasi, a giant penguin from the Eocene of Peru. J. Anat. 2008, 213, 131–147. [Google Scholar] [CrossRef]
- Ksepka, D.T.; Fordyce, R.E.; Ando, T.; Jones, C.M. New fossil penguins (Aves, Sphenisciformes) from the Oligocene of New Zealand reveal the skeletal plan of stem penguins. J. Vertebr. Paleontol. 2012, 32, 235–254. [Google Scholar] [CrossRef]
- Clarke, J.A.; Ksepka, D.T.; Salas-Gismondi, R.A.; Altamirano, J.; Shawkey, M.D.; D’Alba, L.; Vinther, J.; DeVries, T.J.; Baby, P. Fossil evidence for evolution of the shape and color of penguin feathers. Science 2010, 330, 954–957. [Google Scholar] [CrossRef] [PubMed]
- Bengtson, P. Open nomenclature. Palaeontology 1988, 31, 223–227. [Google Scholar]
- Garbe, U.; Randall, T.; Hughes, C.; Davidson, G.; Pangelis, S.; Kennedy, S.J. A new neutron radiography/tomography/imaging station DINGO at OPAL. Phys. Procedia 2015, 69, 27–32. [Google Scholar] [CrossRef]
- Mays, C.; Bevitt, J.; Stilwell, J. Pushing the limits of neutron tomography in palaeontology: Three-dimensional modelling of in situ resin within fossil plants. Palaeontol. Electron. 2017, 20.3.57A, 1–12. [Google Scholar] [CrossRef]
- Crouch, E.M.; Willumsen, P.S.; Kulhanek, D.K.; Gibbs, S.J. A revised Paleocene (Teurian) dinoflagellate cyst zonation from eastern New Zealand. Rev. Palaeobot. Palynol. 2014, 202, 47–79. [Google Scholar] [CrossRef]
- Haidr, N.; Acosta Hospitaleche, C. Feeding habits of Antarctic Eocene penguins from a morphofunctional perspective. N. Jahrb. Geol. Paläontol. Abh. 2012, 263, 125–131. [Google Scholar] [CrossRef]
- Ksepka, D.T.; Bertelli, S. Fossil penguin (Aves: Sphenisciformes) cranial material from the Eocene of Seymour Island (Antarctica). Hist. Biol. 2006, 18, 389–395. [Google Scholar] [CrossRef]
- Kim, J.; Park, H.; Jabłoński, P.G.; Choi, H. The function of the alula in avian flight. Sci. Rep. 2015, 5, 9914. [Google Scholar]
- Nachtigall, W.; Kempf, B. Vergleichende Untersuchungen zur flugbiologischen Funktion des Daumenfittichs (Alula spuria) bei Vögeln. Z. vergl. Physiol. 1971, 71, 326–341. [Google Scholar] [CrossRef]
- Mayr, G. Vergleichend-morphologische Untersuchung der Befiederung des Daumens. Mitt. Zool. Mus. Berlin 72, Suppl.: Ann. Ornithol. 1996, 20, 31–100. [Google Scholar]
- Johansson, L.C.; Aldrin, B.S.W. Kinematics of diving Atlantic puffins (Fratercula arctica L.): Evidence for an active upstroke. J. Exp. Biol. 2002, 205, 371–378. [Google Scholar] [PubMed]
- Williams, C.L.; Hagelin, J.C.; Kooyman, G.L. Hidden keys to survival: The type, density, pattern and functional role of emperor penguin body feathers. Proc. R. Soc. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Barker, P.F.; Filippelli, G.M.; Florindo, F.; Martin, E.E.; Scher, H.D. Onset and role of the Antarctic Circumpolar current. Deep Sea Res. 2007, 54, 2388–2398. [Google Scholar] [CrossRef]
- Thomas, D.B.; Ksepka, D.T.; Fordyce, R.E. Penguin heat-retention structures evolved in a greenhouse Earth. Biol. Lett. 2011, 7, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Near, T.J.; Dornburg, A.; Kuhn, K.L.; Eastman, J.T.; Pennington, J.N.; Patarnello, T.; Zane, L.; Fernández, D.A.; Jones, C.D. Ancient climate change, antifreeze, and the evolutionary diversification of Antarctic fishes. Proc. Natl. Acad. Sci. USA 2012, 109, 3434–3439. [Google Scholar] [CrossRef]
- Rogers, A.D. Evolution and biodiversity of Antarctic organisms: A molecular perspective. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 2191–2214. [Google Scholar] [CrossRef]
- Townsend, C.W. The use of the wings and feet by diving birds. Auk 1909, 26, 234–248. [Google Scholar] [CrossRef]
- Buono, M.R.; Fernández, M.S.; Reguero, M.A.; Marenssi, S.A.; Santillana, S.N.; Mörs, T. Eocene basilosaurid whales from the la Meseta formation, Marambio (Seymour) Island, Antarctica. Ameghiniana 2016, 53, 296–316. [Google Scholar] [CrossRef]
- Lambert, O.; Bianucci, G.; Salas-Gismondi, R.; di Celma, C.; Steurbaut, E.; Urbina, M.; de Muizon, C. An amphibious whale from the middle Eocene of Peru reveals early South Pacific dispersal of quadrupedal cetaceans. Curr. Biol. 2019, 29, 1352–1359. [Google Scholar] [CrossRef]
- Ando, T.; Fordyce, R.E. Evolutionary drivers for flightless, wing-propelled divers in the Northern and Southern Hemispheres. Palaeogeogr., Palaeoclimatol. Palaeoecol. 2014, 400, 50–61. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayr, G.; De Pietri, V.L.; Love, L.; Mannering, A.A.; Bevitt, J.J.; Scofield, R.P. First Complete Wing of a Stem Group Sphenisciform from the Paleocene of New Zealand Sheds Light on the Evolution of the Penguin Flipper. Diversity 2020, 12, 46. https://doi.org/10.3390/d12020046
Mayr G, De Pietri VL, Love L, Mannering AA, Bevitt JJ, Scofield RP. First Complete Wing of a Stem Group Sphenisciform from the Paleocene of New Zealand Sheds Light on the Evolution of the Penguin Flipper. Diversity. 2020; 12(2):46. https://doi.org/10.3390/d12020046
Chicago/Turabian StyleMayr, Gerald, Vanesa L. De Pietri, Leigh Love, Al A. Mannering, Joseph J. Bevitt, and R. Paul Scofield. 2020. "First Complete Wing of a Stem Group Sphenisciform from the Paleocene of New Zealand Sheds Light on the Evolution of the Penguin Flipper" Diversity 12, no. 2: 46. https://doi.org/10.3390/d12020046
APA StyleMayr, G., De Pietri, V. L., Love, L., Mannering, A. A., Bevitt, J. J., & Scofield, R. P. (2020). First Complete Wing of a Stem Group Sphenisciform from the Paleocene of New Zealand Sheds Light on the Evolution of the Penguin Flipper. Diversity, 12(2), 46. https://doi.org/10.3390/d12020046