New Records of Antarctic Tardigrada with Comments on Interpopulation Variability of the Paramacrobiotus fairbanksi Schill, Förster, Dandekar and Wolf, 2010

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Processing
2.2. Microscopy, Morphometrics, and Morphological Nomenclature
2.3. Comparative Material
2.4. DNA Extraction and Genotyping
2.5. Comparative Molecular Analysis
2.6. Statistical Analysis
3. Results
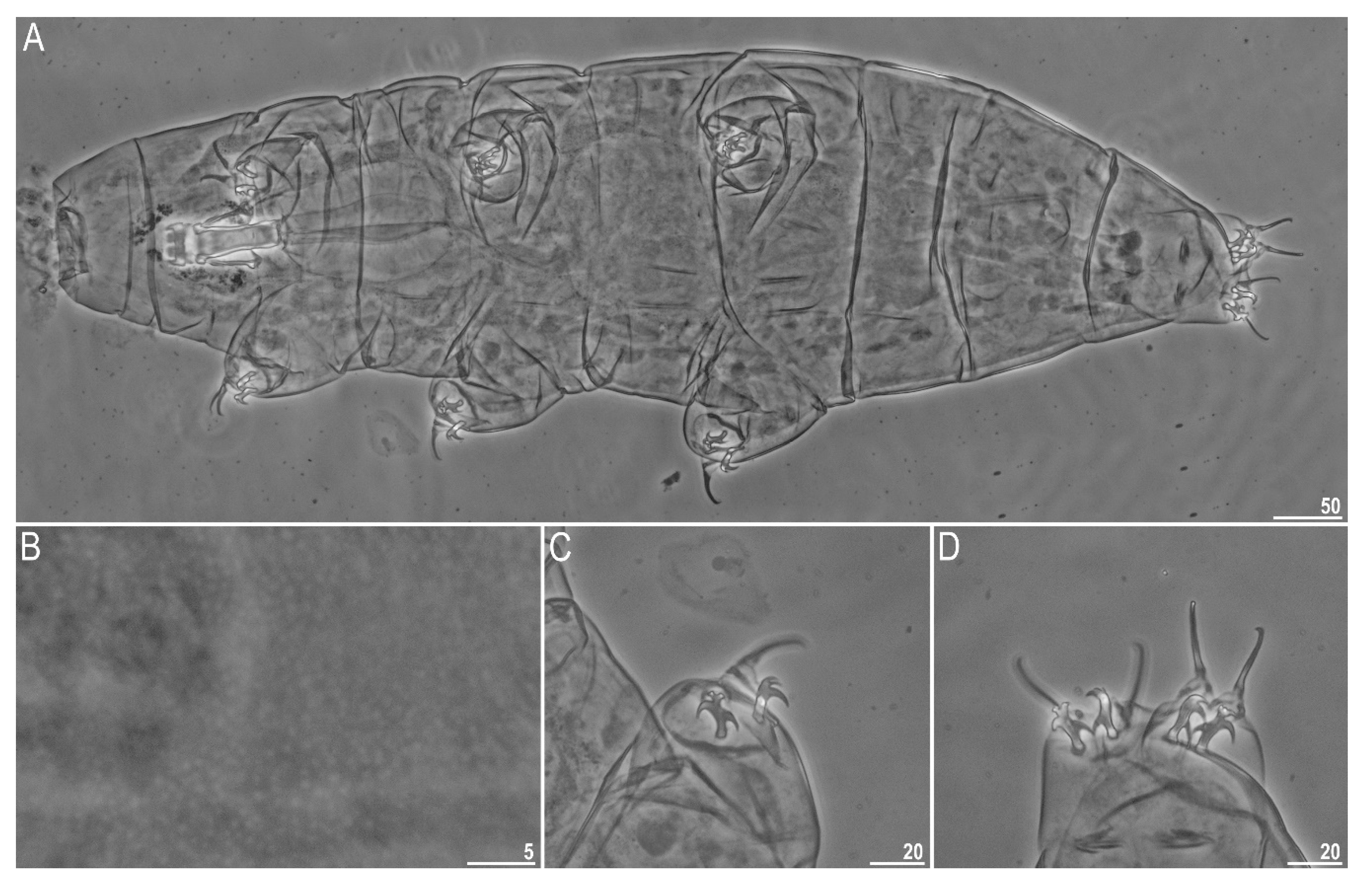
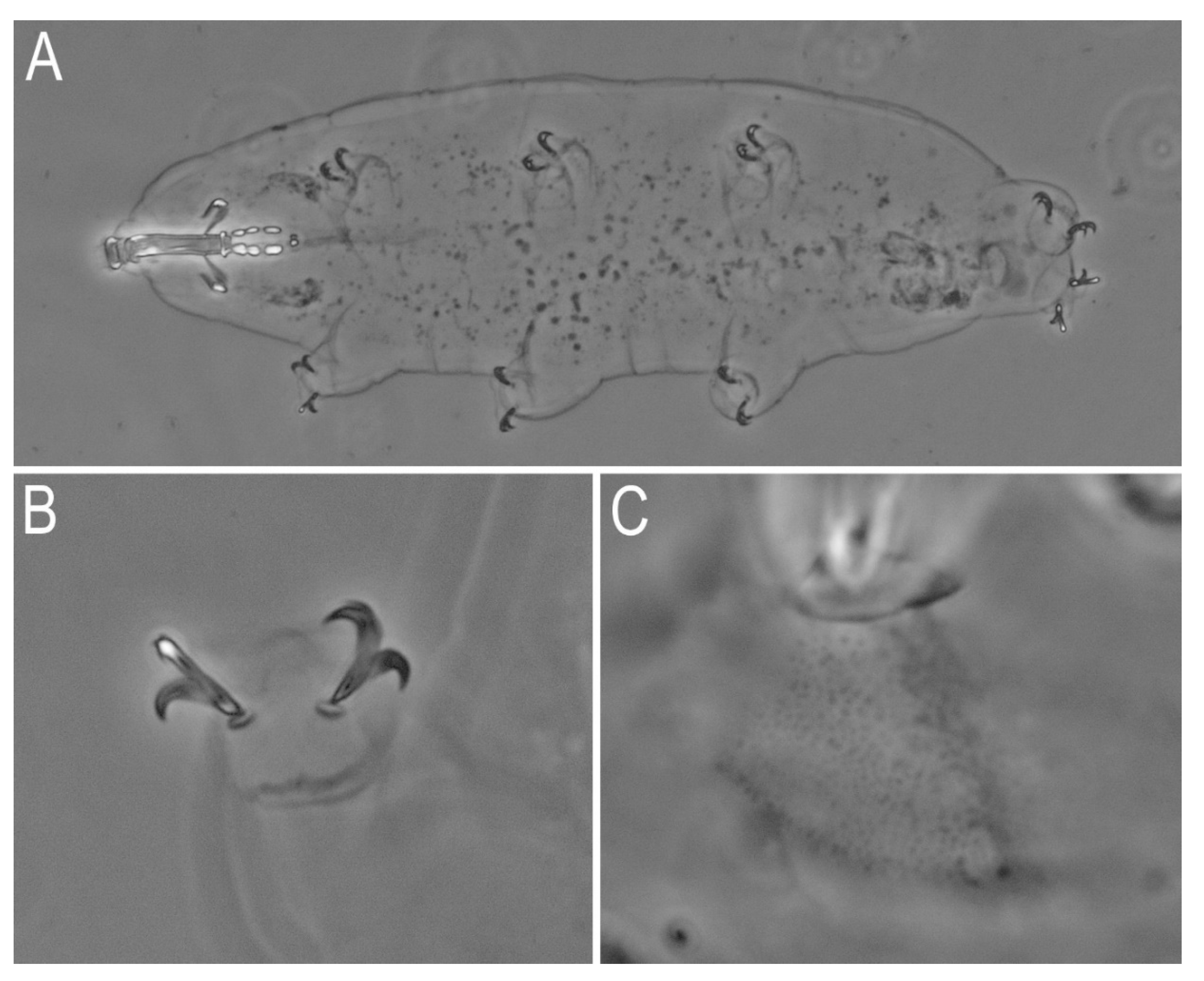
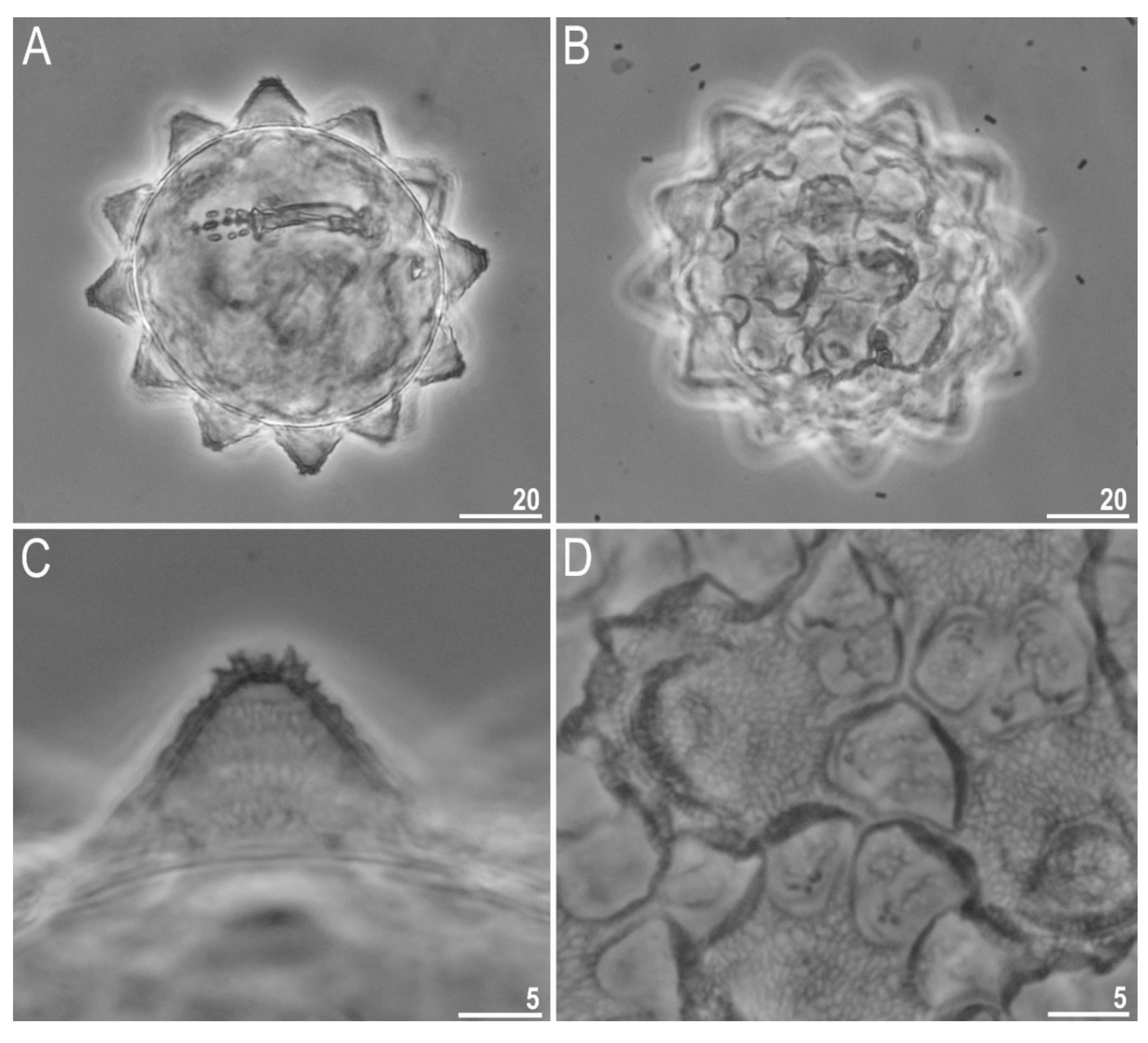
3.1. Species Found
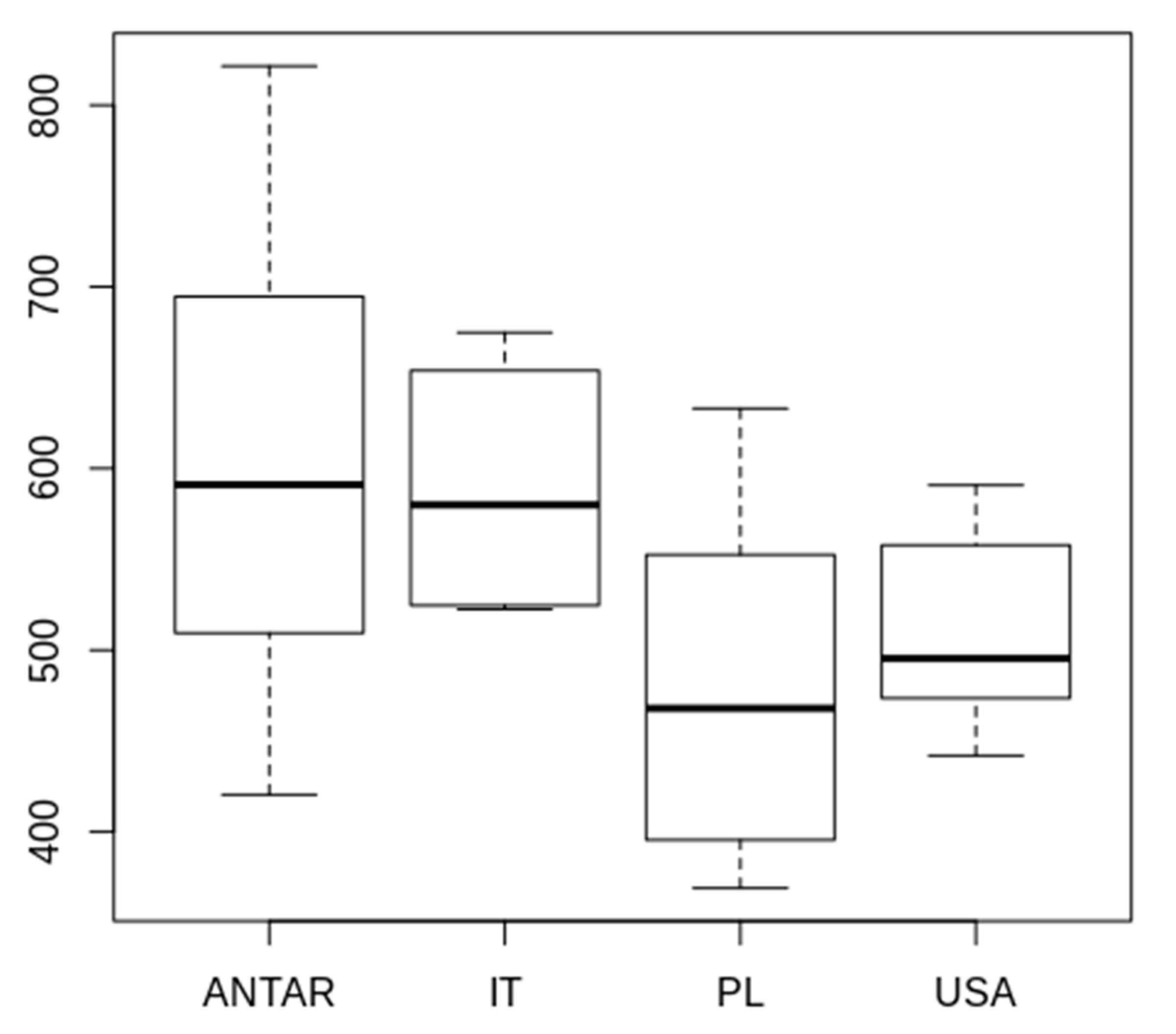
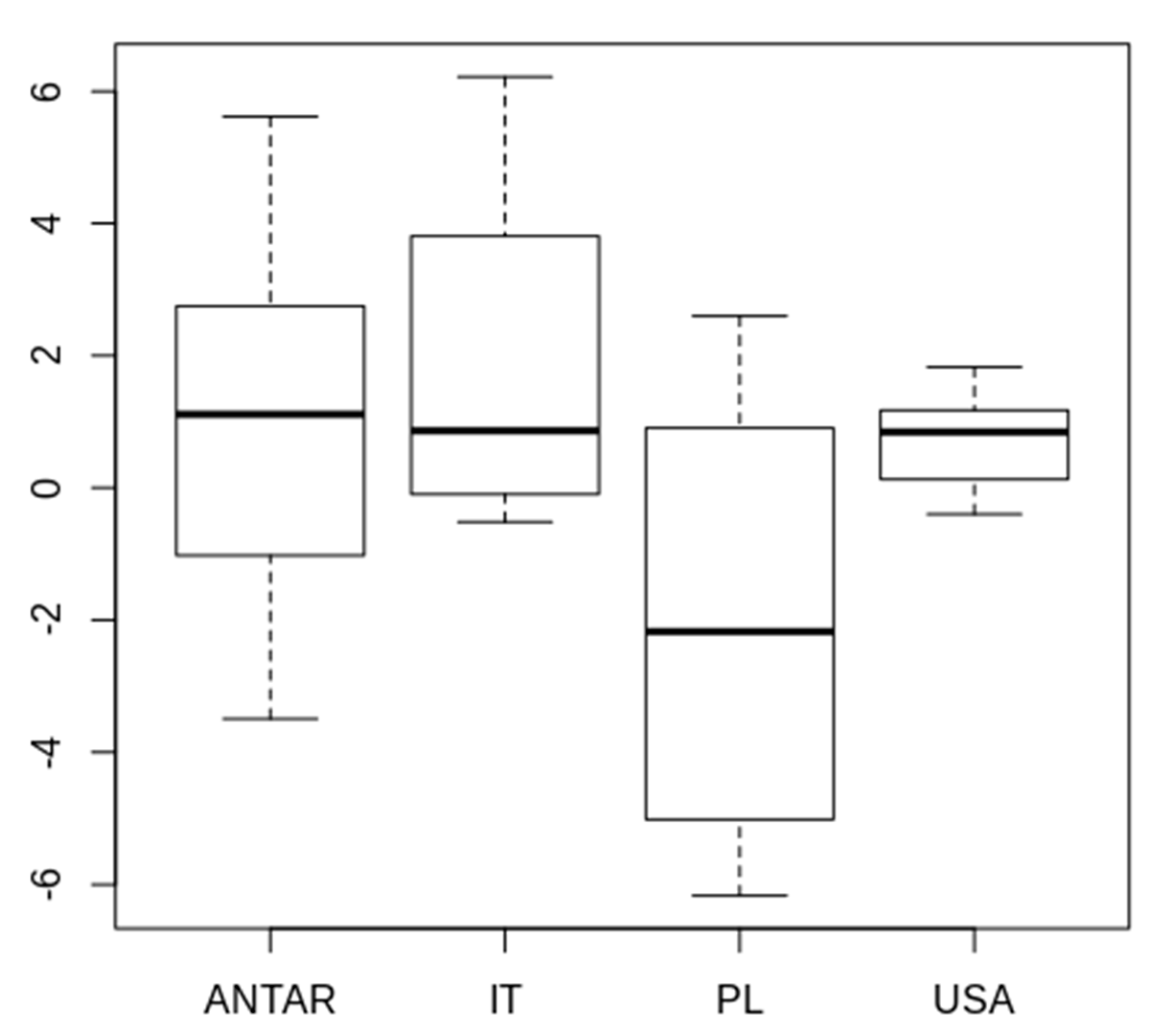
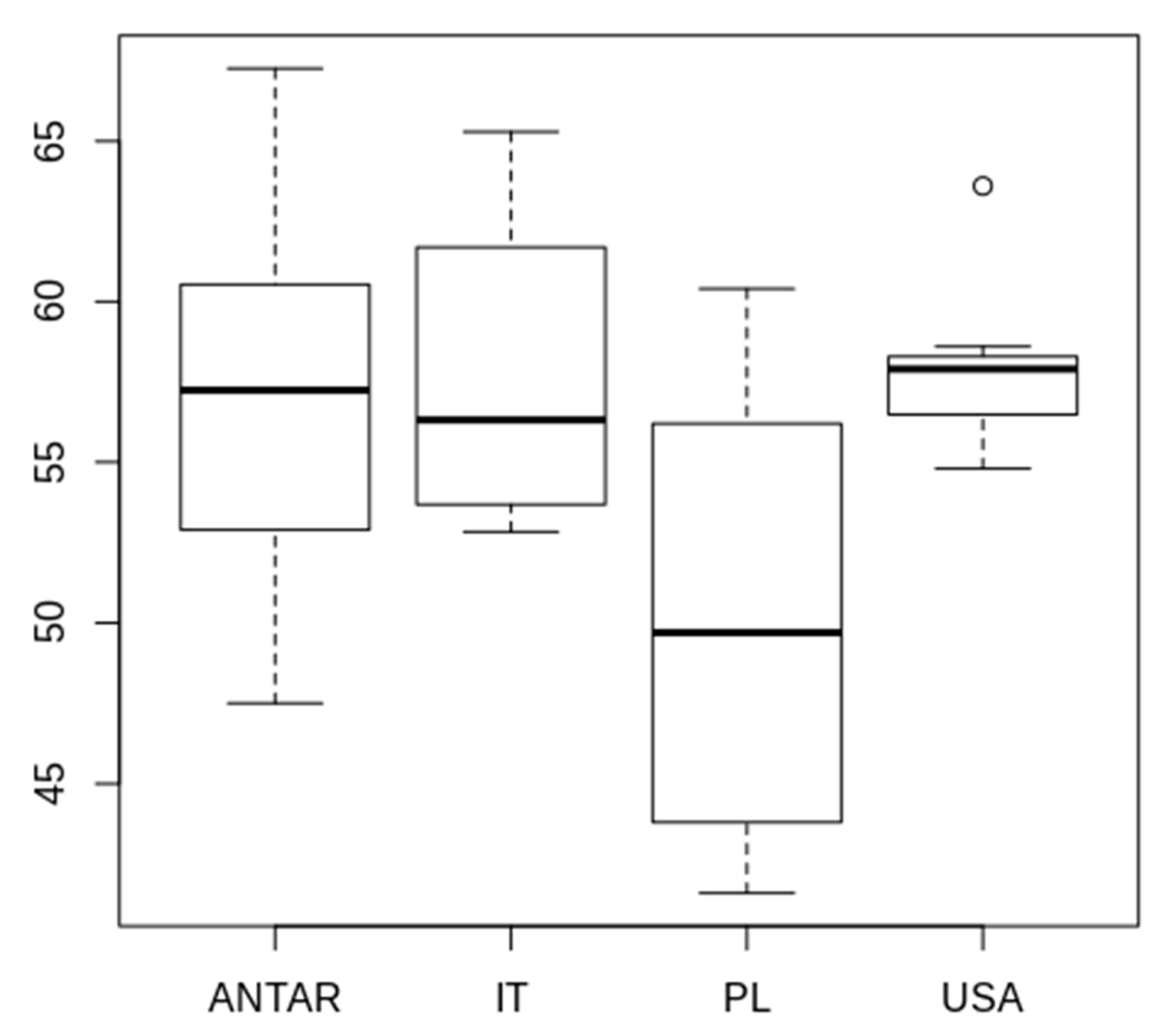
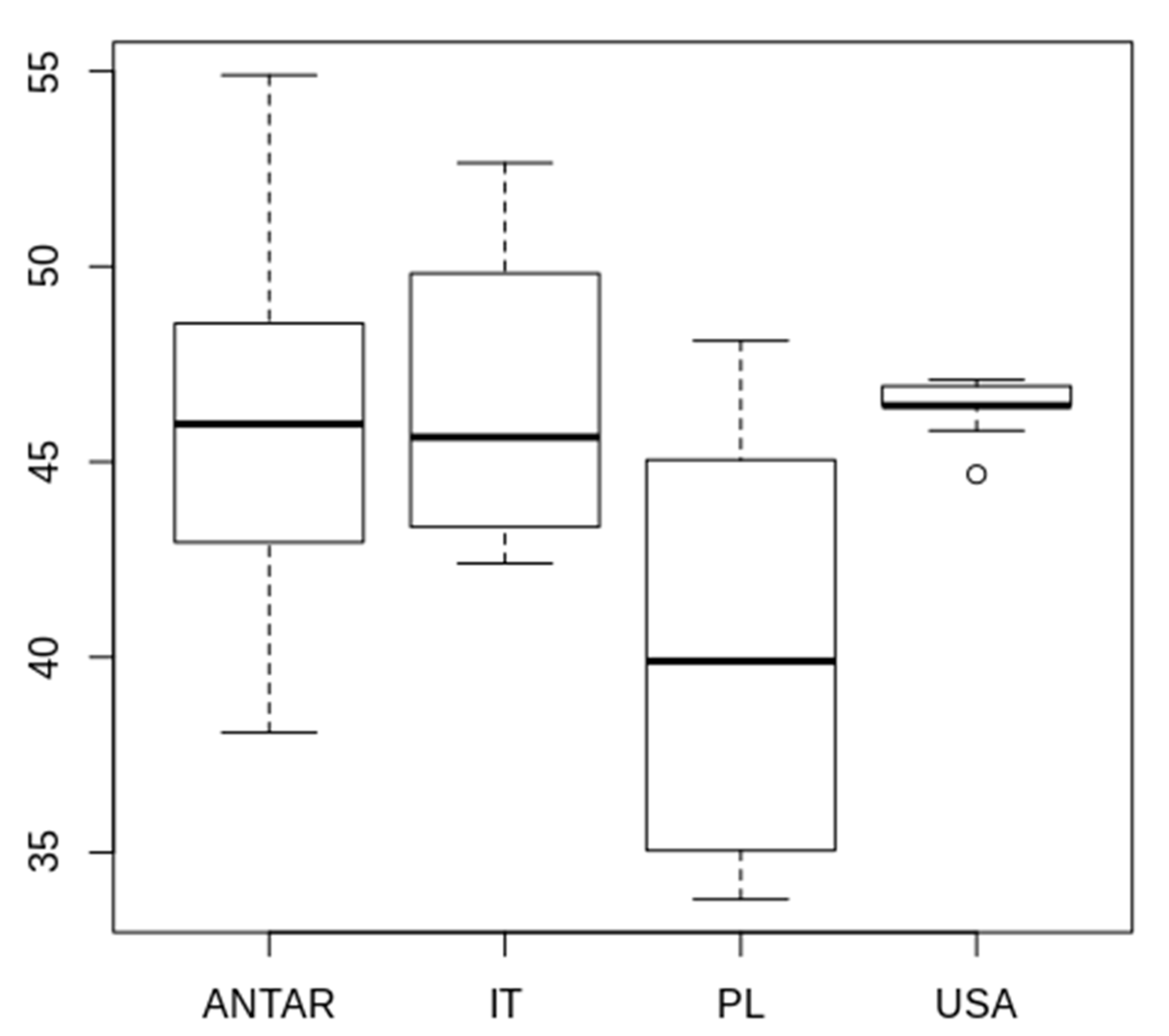
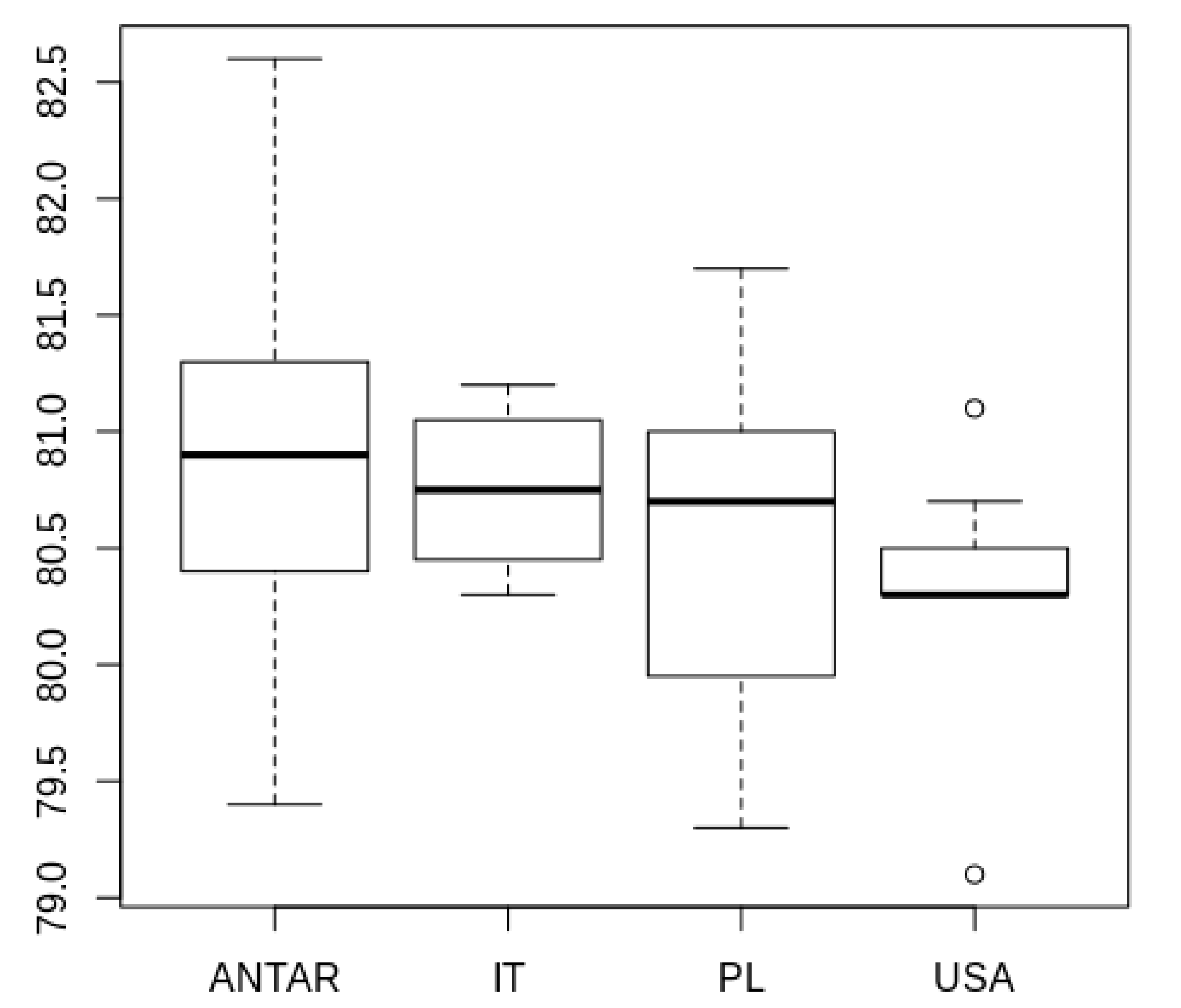
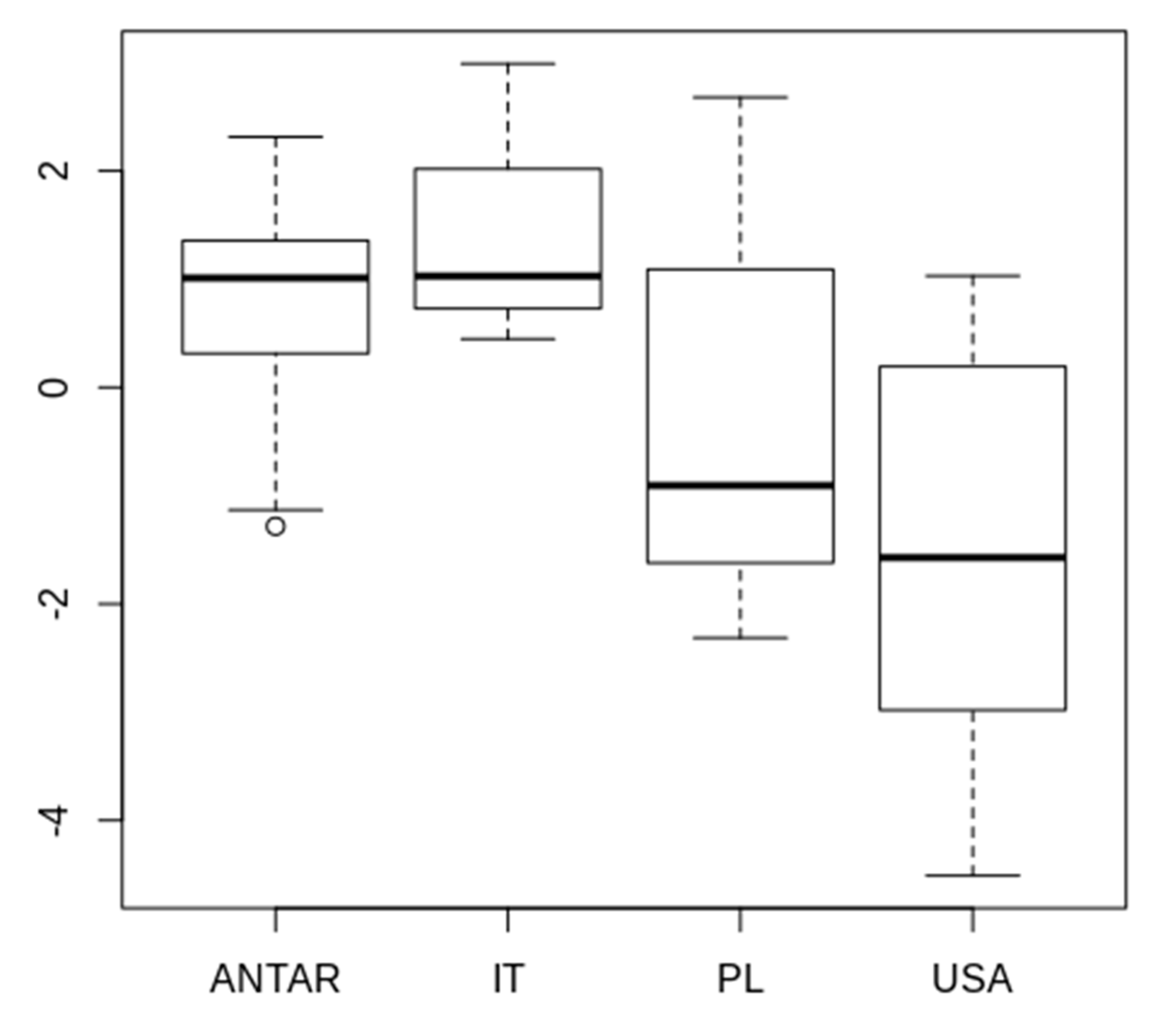
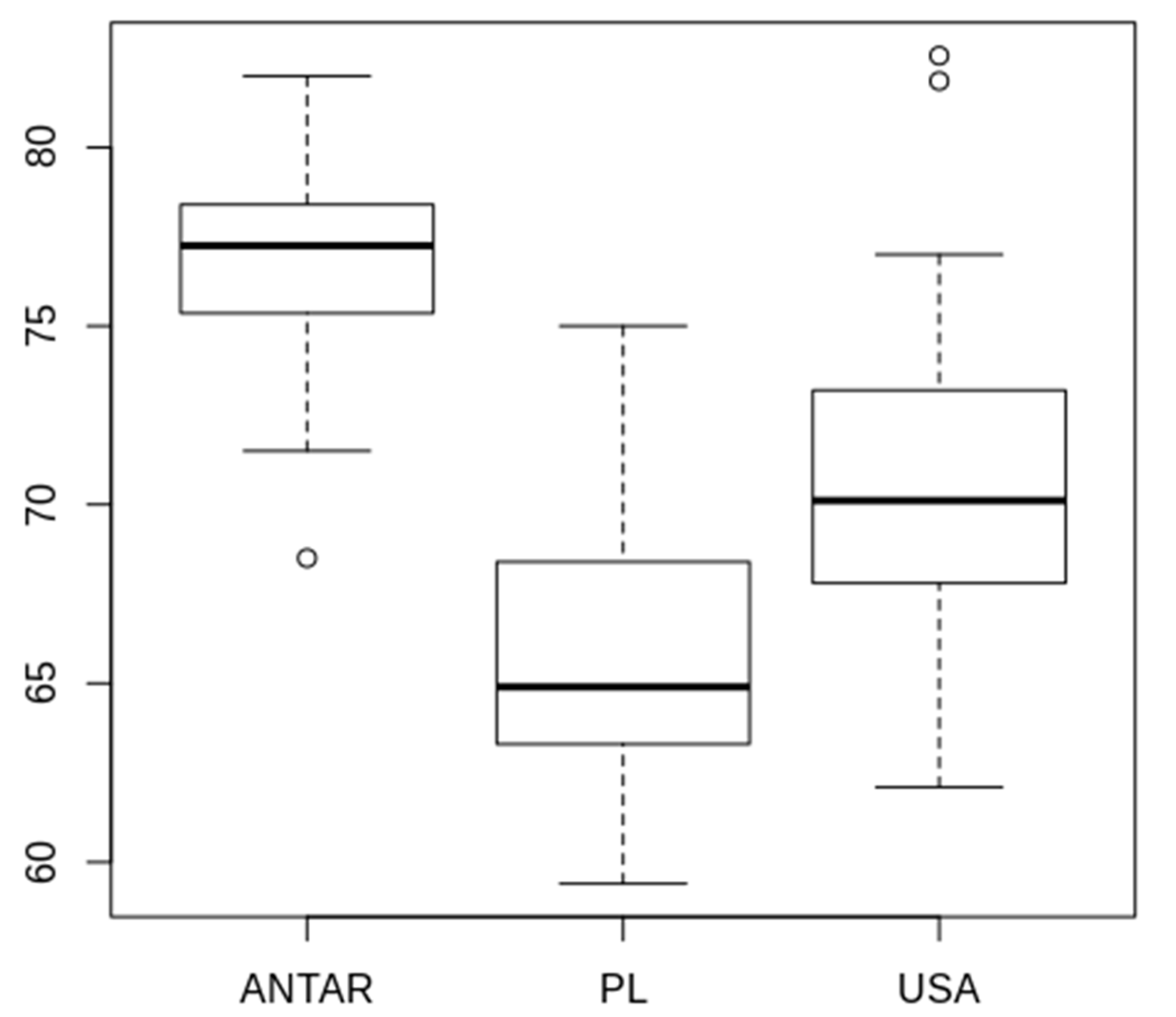
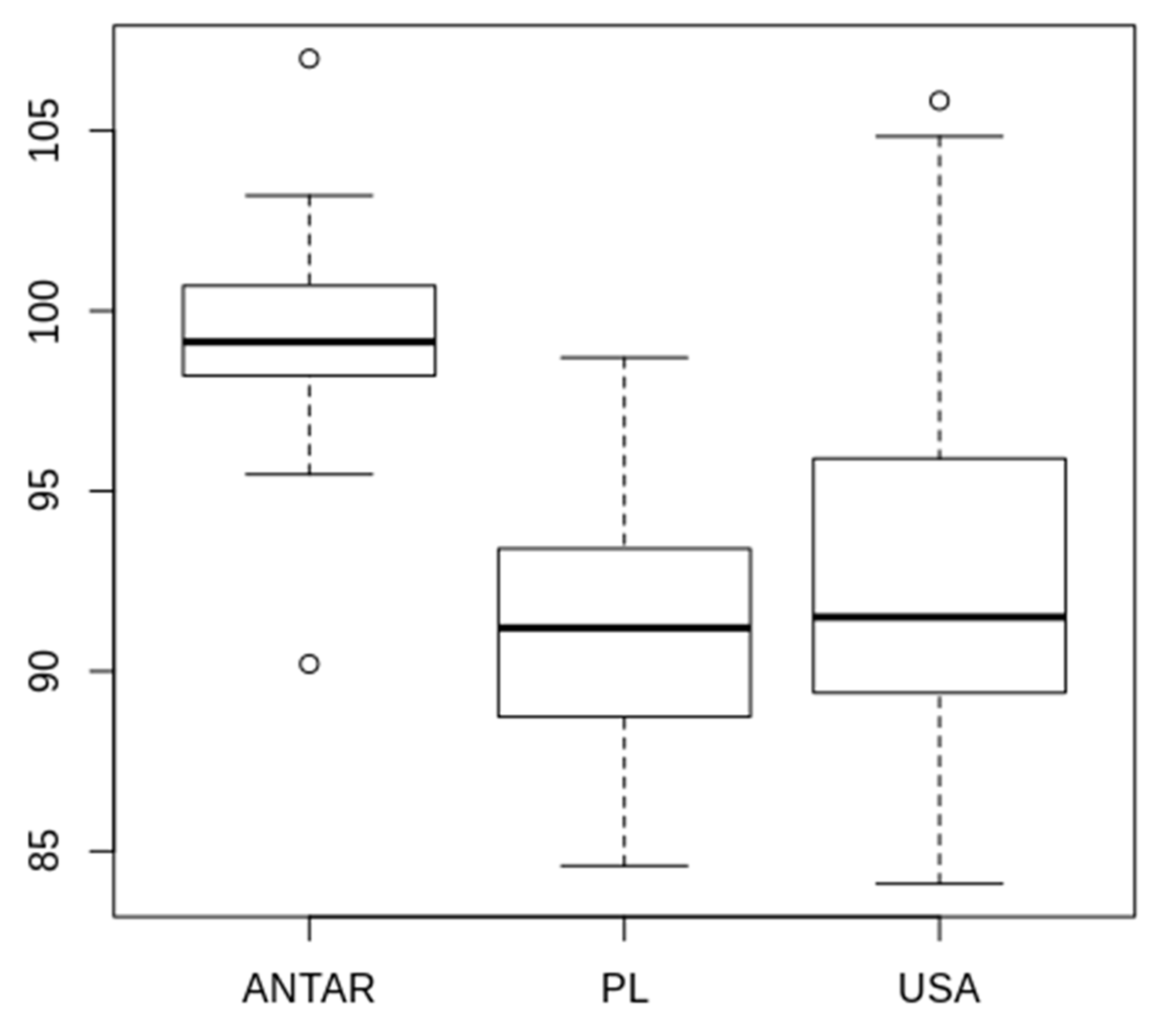
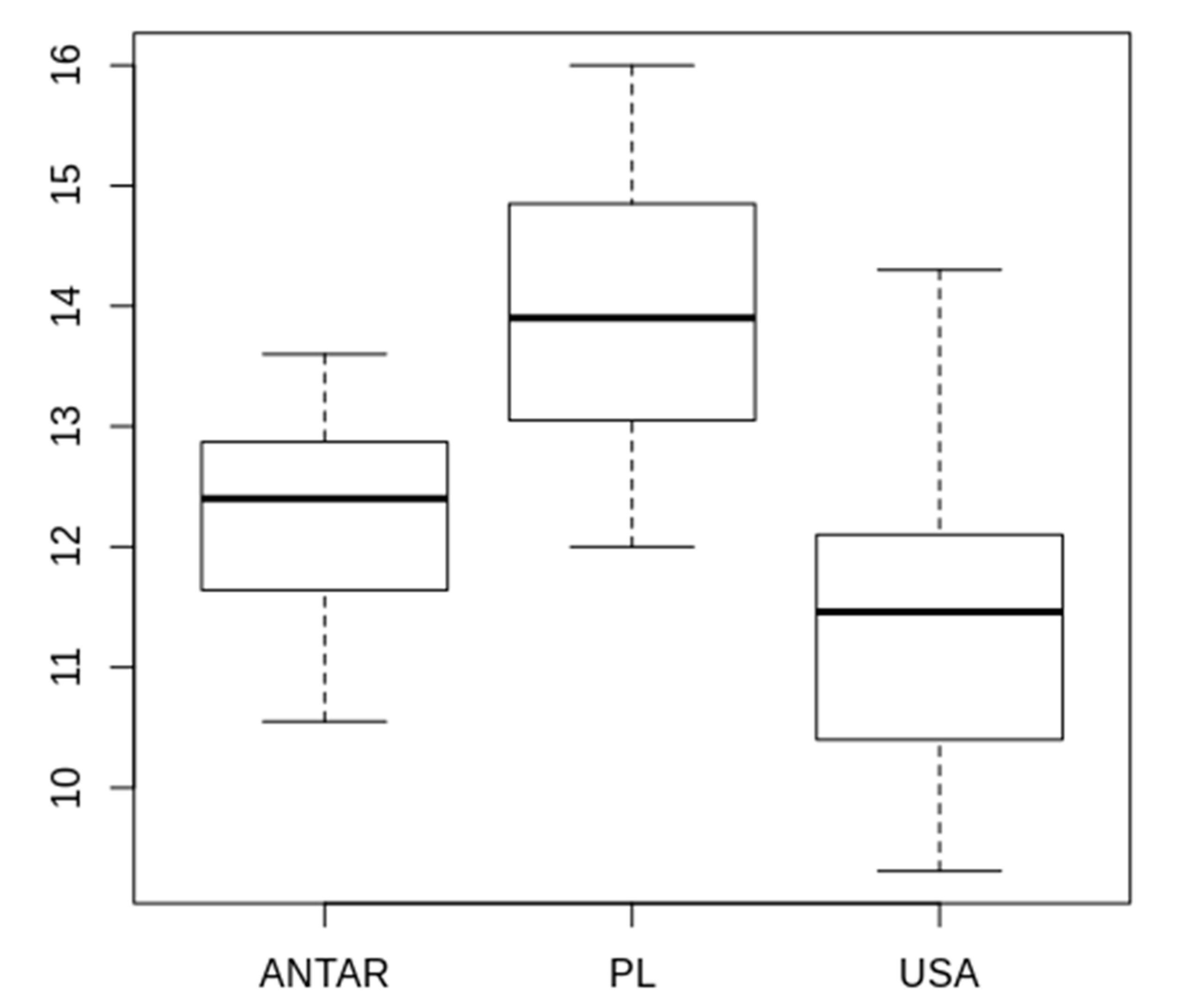
3.2. Morphometric Comparison of Different Populations of the Pam. fairbanksi
3.3. Genetic Comparison of Different Populations of the Pam. fairbanksi
4. Discussion
4.1. Taxa
4.2. Morphometric Comparison of Different Populations of the Pam. fairbanksi
4.3. Genetic Comparison of Different Populations of the Pam. fairbanksi
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kimble, J.M. Cryosols. Permafrost-Affected Soils; Springer: Berlin/Heidelberg, Germany, 2004; pp. 1–94. [Google Scholar]
- Velasco-Castrillón, A.; Gibson, J.A.E.; Stevens, M.I. A review of current Antarctic limno-terrestrial microfauna. Polar Biol. 2014, 37, 1517–1531. [Google Scholar]
- Richters, F. Vorläufiger Bericht über die antarktische Moosfauna. Verh. Dtsch. Zool. Ges. 1904, 14, 236–239. [Google Scholar]
- Pugh, P.J.A.; Convey, P. Surviving out in the cold: Antarctic endemic invertebrates and their refugia. J. Biogeogr. 2008, 35, 2176–2186. [Google Scholar] [CrossRef]
- Cesari, M.; McInnes, S.; Bertolani, R.; Rebecchi, L.; Guidetti, R. Genetic diversity and biogeography of the south polar water bear Acutuncus antarcticus (Eutardigrada: Hypsibiidae)—Evidence that it is a truly pan-Antarctic species. Invertebr. Syst. 2016, 30, 635–649. [Google Scholar] [CrossRef]
- Guidetti, R.; Bertolani, R. Tardigrade taxonomy: An updated check list of the taxa and a list of characters for their identification. Zootaxa 2005, 845, 1–46. [Google Scholar] [CrossRef]
- Degma, P.; Guidetti, R. Notes to the current checklist of Tardigrada. Zootaxa 2007, 1579, 41–53. [Google Scholar] [CrossRef]
- Vicente, F.; Bertolani, R. Considerations on the taxonomy of the Phylum Tardigrada. Zootaxa 2013, 3626, 245–248. [Google Scholar] [CrossRef]
- Degma, P.; Bertolani, R.; Guidetti, R. Actual Checklist of Tardigrada Species; Version 35: Edition: 31-07-2019; Università di Modena e Reggio Emilia: Modena, Italy, 2019. [Google Scholar] [CrossRef]
- Ramazzotti, G.; Maucci, W. Il Phylum Tardigrada. III edizione riveduta e aggiornata. Mem. Ist. Ital. Idrobiol. 1983, 41, 1–1012. [Google Scholar]
- Nelson, D.R.; Guidetti, R.; Rebecchi, L. Phylum Tardigrada. In Ecology and General Biology. Vol. 1. Thorp and Covich’s Freshwater Invertebrates; Throp, J.H., Rogers, D.C., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 347–380. [Google Scholar] [CrossRef]
- Convey, P.; McInnes, S.J. Exceptional tardigrade-dominated ecosystems in Ellsworth Land, Antarctica. Ecology 2005, 86, 519–527. [Google Scholar] [CrossRef]
- Binda, M.G.; Pilato, G.; Lisi, O. Remarks on Macrobiotus furciger Murray, 1906 and description of three new species of the furciger group (Eutardigrada, Macrobiotidae). Zootaxa 2005, 1075, 55–68. [Google Scholar] [CrossRef]
- McInnes, S.J. Echiniscus corrugicaudatus (Heterotardigrada; Echiniscidae) a new species from Ellsworth Land, Antarctica. Polar Biol. 2010, 33, 59–70. [Google Scholar] [CrossRef]
- Pilato, G.; McInnes, S.J.; Lisi, O. Hebesuncus mollispinus (Eutardigrada, Hypsibiidae), a new species from maritime Antarctica. Zootaxa 2012, 3446, 60–68. [Google Scholar] [CrossRef]
- Guidetti, R.; Rebecchi, L.; Cesari, M.; McInnes, S.J. Mopsechiniscus franciscae, a new species of a rare genus of Tardigrada from continental Antarctica. Polar Biol. 2014, 37, 1221–1233. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Janko, K.; Smykla, J.; Michalczyk, Ł. Soil tardigrades from the Antarctic Peninsula with a description of a new species and some remarks on the genus Ramajendas (Eutardigrada, Isohypsibiidae). Polar Rec. 2014, 50, 176–182. [Google Scholar] [CrossRef]
- Tsujimoto, M.; McInnes, S.J.; Convey, P.; Imura, S. Preliminary description of tardigrade species diversity and distribution pattern around coastal Syowa Station and inland Sør Rondane Mountains, Dronning Maud Land, East Antarctica. Polar Biol. 2014, 37, 1361–1367. [Google Scholar] [CrossRef] [Green Version]
- Vecchi, M.; Cesari, M.; Bertolani, R.; Jönsson, K.I.; Rebecchi, L.; Guidetti, R. Integrative systematic studies on tardigrades from Antarctica identify new genera and new species within Macrobiotoidea and Echiniscoidea. Invertebr. Syst. 2016, 30, 303–322. [Google Scholar] [CrossRef]
- Guidetti, R.; McInnes, S.J.; Cesari, M.; Rebecchi, L.; Rota-Stabelli, O. Evolutionary scenarios for the origin of an Antarctic tardigrade species based on molecular clock analyses and biogeographic data. Contrib. Zool. 2017, 86, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Pilato, G.; Sabella, G.; D’Urso, V.; Lisi, O. Two new species of Eutardigrada from Victoria Land, Antarctica. Zootaxa 2017, 4317, 541–558. [Google Scholar] [CrossRef]
- Dastych, H. Redescription and revalidation of the sub-Antarctic tardigrade Hypsibius murrayi (Richters, 1907) based on the rediscovered type material (Tardigrada, Panarthropoda). Entomol. Heute 2018, 30, 95–115. [Google Scholar]
- Kaczmarek, Ł.; Parnikoza, I.; Gawlak, M.; Esefeld, J.; Peter, H.-U.; Kozeretska, I.; Roszkowska, M. Tardigrades from Larus dominicanus Lichtenstein, 1823 nests on the Argentine Islands (maritime Antarctic). Polar Biol. 2018, 41, 283–301. [Google Scholar] [CrossRef] [Green Version]
- Guidetti, R.; Massa, E.; Bertolani, R.; Rebecchi, L.; Cesari, M. Increasing knowledge of Antarctic biodiversity: New endemic taxa of tardigrades (Eutardigrada; Ramazzottiidae) and their evolutionary relationships. Syst. Biodivers. 2019, 17, 573–593. [Google Scholar] [CrossRef]
- Schill, R.O.; Forster, F.; Dandekar, T.; Wolf, N. Using compensatory base change analysis of internal transcribed spacer 2 secondary structures to identify three new species in Paramacrobiotus (Tardigrada). Org. Divers. Evol. 2010, 10, 287–296. [Google Scholar] [CrossRef]
- Guil, N.; Giribet, G. A comprehensive molecular phylogeny of tardigrades—Adding genes and taxa to a poorly resolved phylum-level phylogeny. Cladistics 2012, 28, 21–49. [Google Scholar] [CrossRef]
- Kosztyła, P.; Stec, D.; Morek, W.; Gąsiorek, P.; Zawierucha, Z.; Michno, K.; Małek, D.; Hlebowicz, K.; Ufir, K.; Laska, A.; et al. Experimental taxonomy confirms the environmental stability of morphometric traits in a taxonomically challenging group of microinvertebrates. Zool. J. Linn. Soc. 2016, 178, 765–775. [Google Scholar]
- Guidetti, R.; Cesari, M.; Bertolani, R.; Altiero, T.; Rebecchi, L. High diversity in species, reproductive modes and distribution within the Paramacrobiotus richtersi complex (Eutardigrada, Macrobiotidae). Zool. Lett. 2019, 5, 1. [Google Scholar] [CrossRef]
- Guidetti, R.; Gandolfi, A.; Rossi, V.; Bertolani, R. Phylogenetic analysis in Macrobiotidae (Eutardigrada, Parachela): A combined morphological and molecular approach. Zool. Scr. 2005, 34, 235–244. [Google Scholar] [CrossRef]
- Murray, J. Arctiscoida. Proc. Roy. Irish Acad. 1911, 31, 1–16. [Google Scholar]
- Beijerinck, M.W. De infusies en de ontdekking der bakterien. Jaarb. Kon Akad. Wetensch. 1913, 1–28. [Google Scholar]
- Baas Becking, L.G.M. Geobiologie of Inleiding Tot de Milieukunde; W.P. Van Stockum & Zoon N.V: Den Haag, The Netherlands, 1934; pp. 1–263. [Google Scholar]
- Cardillo, M.; Bromhan, L. Body size and risk of extinction in Australian mammals. Conserv. Biol. 2001, 15, 1435–1440. [Google Scholar] [CrossRef]
- Finlay, B.J. Global dispersal of free living microbial eukaryote species. Science 2002, 296, 1061–1063. [Google Scholar] [CrossRef] [Green Version]
- Foissner, W. Biogeography and dispersal of micro-organisms: A review emphasizing protists. Acta Protozool. 2006, 45, 111–136. [Google Scholar]
- Schön, I.; Martens, K.; Dijk, P. Lost Sex. The Evolutionary Biology of Parthenogenesis; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–615. [Google Scholar]
- Pilato, G.; Binda, M.G. Biogeography and limno-terrestrial tardigrades: Are they truly incompatible binomials? Zool. Anz. 2001, 240, 511–516. [Google Scholar] [CrossRef]
- Guil, N.; Sánchez-Moreno, S.; Machordom, A. Local biodiversity patterns in micrometazoans: Are tardigrades everywhere? Syst. Biodivers. 2009, 7, 259–268. [Google Scholar] [CrossRef]
- Faurby, S.; Jorgensen, A.; Kristensen, R.M.; Funch, P. Distribution and speciation in marine intertidal tardigrades: Testing the roles of climatic and geographical isolation. J. Biogeogr. 2012, 39, 1596–1607. [Google Scholar] [CrossRef]
- Gąsiorek, P.; Vončina, K.; Michalczyk, Ł. Echiniscus testudo (Doyère, 1840) in New Zealand: Anthropogenic dispersal or evidence for the ‘Everything is Everywhere’ hypothesis? N. Z. J. Zool. 2019, 46, 174–181. [Google Scholar]
- Doyère, P.L.N. Memoire sur les Tardigrades. Ann. Sci. Nat. Zool. Paris Ser. 2 1840, 14, 269–362. [Google Scholar]
- Stec, D.; Smolak, R.; Kaczmarek, Ł.; Michalczyk, Ł. An integrative description of Macrobiotus paulinae sp. nov. (Tardigrada: Eutardigrada: Macrobiotidae: Hufelandi group) from Kenya. Zootaxa 2015, 4052, 501–526. [Google Scholar] [CrossRef] [Green Version]
- Beasley, C.W. The Phylum Tardigrada, 3rd ed.; Ramazzotti, G., Maucci, W., Eds.; English Translation; Clark Beasley: Abilene, TX, USA, 1995; pp. 1–1014. [Google Scholar]
- Pilato, G.; Binda, M.G. Definition of families, subfamilies, genera and subgenera of the Eutardigrada, and keys to their identification. Zootaxa 2010, 2404, 1–52. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, Ł.; Michalczyk, Ł. The Macrobiotus hufelandi group (Tardigrada) revisited. Zootaxa 2017, 4363, 101–123. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Cytan, J.; Zawierucha, K.; Diduszko, D.; Michalczyk, Ł. Tardigrades from Peru (South America), with descriptions of three new species of Parachela. Zootaxa 2014, 3790, 357–379. [Google Scholar] [CrossRef] [Green Version]
- Pilato, G. Analisi di nuovi caratteri nello studio degli Eutardigradi. Animalia 1981, 8, 51–57. [Google Scholar]
- Michalczyk, Ł.; Kaczmarek, Ł. The Tardigrada Register: A comprehensive online data repository for tardigrade taxonomy. J. Limnol. 2013, 72, 175–181. [Google Scholar] [CrossRef]
- Bertolani, R.; Guidetti, R.; Marchioro, T.; Altiero, T.; Rebechci, L.; Cesari, M. Phylogeny of Eutardigrada: New molecular data and their morphological support lead to the identification of new evolutionary lineages. Mol. Phylogenetics Evol. 2014, 76, 110–126. [Google Scholar] [CrossRef] [PubMed]
- Perry, E.; Miller, W.R.; Kaczmarek, Ł. Recommended abbreviations for the names of genera of the phylum Tardigrada. Zootaxa 2019, 4608, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Nederström, P. Die bis jetzt aus Finland bekannten Tardigraden. Acta Soc. Fauna Flora Fenn. 1919, 46, 1–25. [Google Scholar]
- Dastych, H. The Tardigrada from Antarctica with description of several new species. Acta Zool. Cracov. 1984, 27, 377–436. [Google Scholar]
- Pilato, G. The taxonomic value of the structures for the insertion of the stylet muscles in the Eutardigrada, and description of a new genus. Zootaxa 2013, 3721, 365–378. [Google Scholar] [CrossRef] [Green Version]
- Pilato, G.; Binda, M.G. Acutuncus, a new genus of Hypsibiidae (Eutardigrada). Entomol. Mitt. Zool. Mus. Hamburg 1997, 12, 159–162. [Google Scholar]
- Casquet, J.; Thebaud, C.; Gillespie, R.G. Chelex without boiling, a rapid and easy technique to obtain stable amplifiable DNA from small amounts of ethanol-stored spiders. Mol. Ecol. Resour. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- Sands, C.J.; Mcinnes, S.J.; Marley, N.J.; Goodall–Copestake, W.P.; Convey, P.; Linse, K. Phylum Tardigrada: An “individual” approach. Cladistics 2008, 24, 861–871. [Google Scholar] [CrossRef]
- Mironov, S.V.; Dabert, J.; Dabert, M. A new feather mite species of the genus Proctophyllodes Robin, 1877 (Astigmata: Proctophyllodidae) from the long–tailed tit Aegithalos caudatus (Passeriformes: Aegithalidae): Morphological description with DNA barcode data. Zootaxa 2012, 3253, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. 2012. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positionspecific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Goujon, M.; Mcwilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; Lopez, R. A new bioinformatics analysis tools framework at EMBL–EBI. Nucleic Acids Res. 2010, 38, 695–699. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Roszkowska, M.; Stec, D.; Ciobanu, D.A.; Kaczmarek, Ł. (2016) Tardigrades from Nahuel Huapi National Park (Argentina, South America) with descriptions of two new Macrobiotidae species. Zootaxa 2016, 4105, 243–260. [Google Scholar] [CrossRef]
- Morek, W.; Gąsiorek, P.; Stec, D.; Blagden, B.; Michalczyk, Ł. Experimental taxonomy exposes ontogenetic variability and elucidates the taxonomic value of claw configuration in Milnesium Doyère, 1840 (Tardigrada: Eutardigrada: Apochela). Contrib. Zool. 2016, 85, 173–200. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, Ł.; Grobys, D.; Kulpa, A.; Bartylak, T.; Kmita, H.; Kepel, M.; Kepel, A.; Roszkowska, M. Two new species of the genus Milnesium Doyère, 1840 (Tardigrada, Apochela, Milnesiidae) from Madagascar. ZooKeys 2019, 884, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Morek, W.; Stec, D.; Gąsiorek, P.; Surmacz, B.; Michalczyk, Ł. Milnesium tardigradum Doyère, 1840: The first integrative study of inter-population variability in a tardigrade species. J. Zool. Syst. Evol. Res. 2019, 57, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Surmacz, B.; Morek, W.; Michalczyk, Ł. What if multiple claw configurations are present in a sample: A case study with the description of Milnesium pseudotardigradum sp. nov. with unique developmental variability. Zool. Stud. 2019, 58, 32. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Coordinates | Sample Description | Expedition |
|---|---|---|---|
| 1 | 67°39′S, 46°09′E | Vechernia Mt Base, lichen Usnea sphacelata. | 9 BAE (2016–2017) |
| 2 | 67°39′S, 46°09′E | Vechernia Mt Base, lichen Umbilicaria aprina. | 9 BAE (2016–2017) |
| 3 | 69°24′S, 76°24′E | Progress Base, lichen Umbilicaria decussata. | 9 BAE (2016–2017) |
| 4 | 67°39′S, 46°09′E | Vechernia Mt Base, lichen Xanthoria elegans. | 9 BAE (2016–2017) |
| 5 | 67°39′S, 46°09′E | Vechernia Mt Base, algae-bacterial mat from Nizhne Lake. | 7 BAE (2014–2015) |
| 6 | 70°46′S, 11°49′E | Vechernia Mt Base, algae-bacterial mat from Nizhne Lake. | 5 BAE (2012–2013) |
| 7 | 67°39′S, 46°09′E | Vechernia Mt Base, moss Ceratodon purpureus. | 7 BAE (2014–2015) |
| 8 | 69°24′S, 76°24′E | Progress Base, malanized soil surface algae. | 9 BAE (2016–2017) |
| 9 | 69°23′S, 76°23′E | Progress Base, algae Parsiola crispa. | 9 BAE (2016–2017) |
| 10 | 69°22′S, 76°23′E | Progress Base, sediment from bottom of the snow collector for fresh-water maker. | 9 BAE (2016–2017) |
| 11 | 67°39′S, 46°09′E | Progress Base, soil with vegetation. | 9 BAE (2016–2017) |
| Taxon | s/s + e | Remarks |
|---|---|---|
| Acu. antarcticus (Richters [3]) | 6/1, 8/59, 9/205 | Pan-Antarctic species [5] |
| Bar. pseudowendti (Dastych [52]) | 1/53, 2/3, 4/1 | Endemic species known only from Antarctica [52] |
| Das. improvisus (Dastych [52]) | 2/9 | Species of the monotypic genus endemic for Antarctica This is a second report of this species [52] |
| Heb. mollispinus Pilato et al. [15] | 2/50, 4/4, 7/19 | Species known only from Antarctica and South America [15,66] |
| Mes. blocki (Dastych [52]) | 7/25 + 68 | Endemic species known only from Antarctica [52]. This is second report of this species |
| Mesobiotus sp. | 3/4, 2/2 | Undefined species due to lack of eggs in the examined samples |
| Mil. quadrifidum Nederström [51] | 2/1 | Species with unclear taxonomic position [27] |
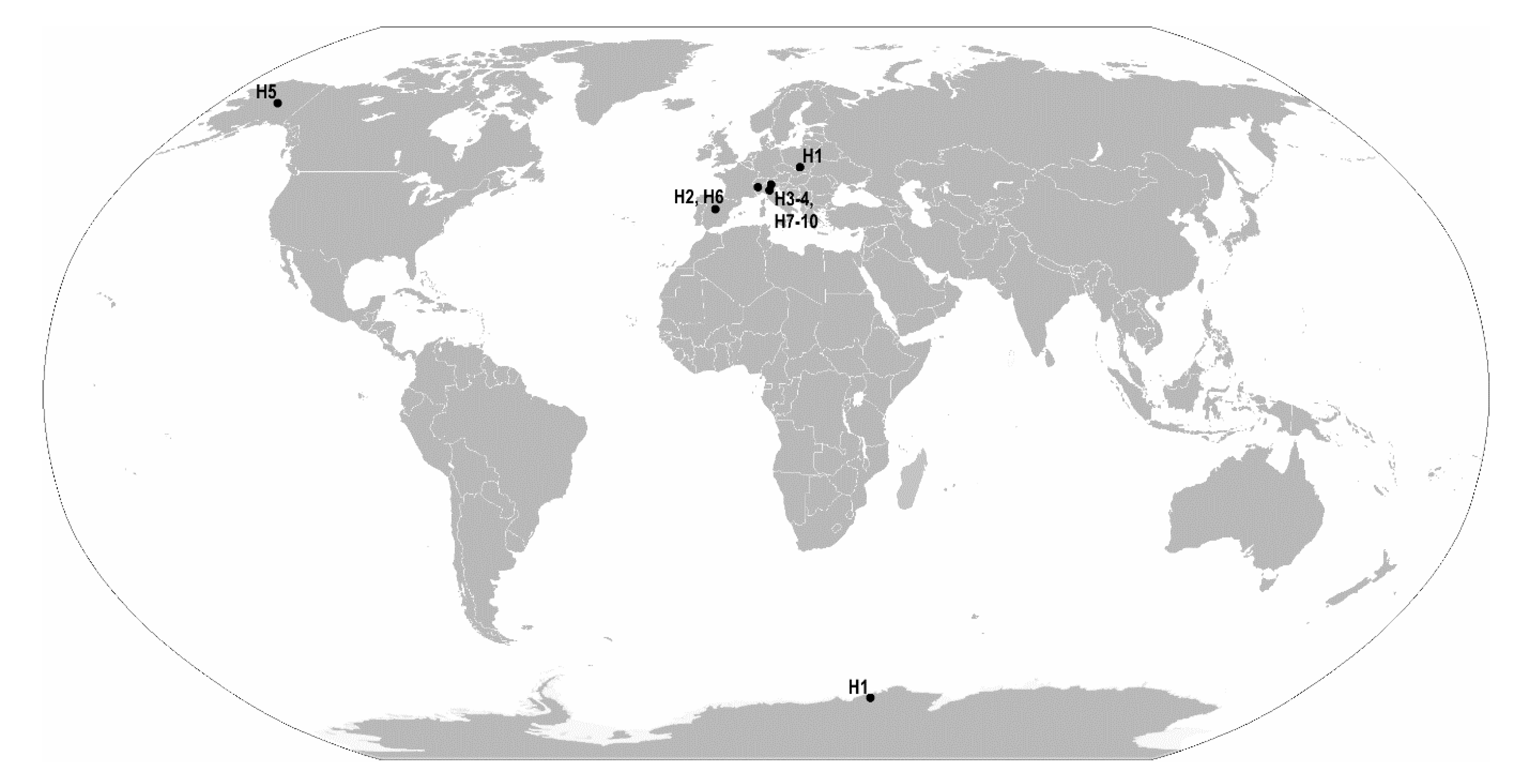
| Pam. fairbanksi Schill et al. [25] | 7/25 + 59 | Probably cosmopolitan species up to now reported from Italy, Poland, Spain, USA (Alaska; type locality) [25,26,27,28,29] |
| CHARACTER | N | RANGE | MEAN | SD | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | ||||||
| Body length | 16 | 420 | – | 821 | – | 595 | – | 112 | – | ||
| Buccopharyngeal tube | |||||||||||
| Buccal tube length | 16 | 47.5 | – | 67.3 | – | 57.1 | – | 5.4 | – | ||
| Stylet support insertion point | 16 | 38.1 | – | 54.9 | 79.4 | – | 82.6 | 46.1 | 80.9 | 4.4 | 0.8 |
| Buccal tube external width | 16 | 10.5 | – | 14.8 | 21.8 | – | 22.5 | 12.7 | 22.2 | 1.2 | 0.2 |
| Buccal tube internal width | 16 | 7.7 | – | 11.3 | 16.0 | – | 18.1 | 9.7 | 16.9 | 1.0 | 0.5 |
| Ventral lamina length | 16 | 29.6 | – | 42.6 | 60.4 | – | 64.7 | 35.9 | 62.9 | 3.4 | 1.2 |
| Placoid lengths | |||||||||||
| Macroplacoid 1 | 16 | 7.8 | – | 12.1 | 16.4 | – | 18.4 | 10.0 | 17.4 | 1.3 | 0.7 |
| Macroplacoid 2 | 16 | 6.6 | – | 10.1 | 13.9 | – | 15.9 | 8.6 | 15.0 | 1.0 | 0.6 |
| Macroplacoid 3 | 16 | 9.2 | – | 14.0 | 19.4 | – | 21.1 | 11.7 | 20.4 | 1.3 | 0.5 |
| Microplacoid | 16 | 4.1 | – | 6.9 | 8.2 | – | 10.9 | 5.1 | 8.9 | 0.7 | 0.6 |
| Macroplacoid row | 16 | 26.8 | – | 38.7 | 54.1 | – | 59.0 | 32.8 | 57.4 | 3.5 | 1.1 |
| Placoid row | 16 | 36.1 | – | 52.6 | 71.7 | – | 81.0 | 44.5 | 77.9 | 4.9 | 2.0 |
| Claw 1 heights | |||||||||||
| External primary branch | 15 | 12.3 | – | 17.6 | 25.7 | – | 26.9 | 15.0 | 26.3 | 1.6 | 0.5 |
| External secondary branch | 15 | 9.8 | – | 13.9 | 20.0 | – | 21.5 | 11.8 | 20.6 | 1.2 | 0.4 |
| Internal primary branch | 15 | 11.9 | – | 16.4 | 24.1 | – | 25.5 | 14.2 | 24.9 | 1.4 | 0.5 |
| Internal secondary branch | 15 | 9.8 | – | 13.3 | 19.3 | – | 21.0 | 11.6 | 20.3 | 1.1 | 0.4 |
| Claw 2 heights | |||||||||||
| External primary branch | 15 | 13.1 | – | 18.9 | 27.0 | – | 29.1 | 16.2 | 28.3 | 1.7 | 0.6 |
| External secondary branch | 15 | 9.9 | – | 15.8 | 20.6 | – | 23.5 | 12.7 | 22.3 | 1.6 | 1.2 |
| Internal primary branch | 15 | 11.4 | – | 15.8 | 22.7 | – | 25.8 | 14.0 | 24.6 | 1.3 | 0.9 |
| Internal secondary branch | 15 | 9.8 | – | 13.7 | 19.8 | – | 22.1 | 11.8 | 20.7 | 1.1 | 0.7 |
| Claw 3 heights | |||||||||||
| External primary branch | 15 | 13.3 | – | 18.6 | 27.4 | – | 29.1 | 16.1 | 28.4 | 1.6 | 0.5 |
| External secondary branch | 15 | 10.0 | – | 15.2 | 21.1 | – | 22.7 | 12.4 | 21.8 | 1.4 | 0.4 |
| Internal primary branch | 15 | 11.7 | – | 16.4 | 23.9 | – | 24.7 | 13.9 | 24.3 | 1.4 | 0.3 |
| Internal secondary branch | 15 | 9.6 | – | 13.8 | 19.1 | – | 21.7 | 11.4 | 20.0 | 1.3 | 0.8 |
| Claw 4 heights | |||||||||||
| Anterior primary branch | 13 | 14.0 | – | 19.7 | 29.0 | – | 30.1 | 17.0 | 29.5 | 1.7 | 0.4 |
| Anterior secondary branch | 13 | 10.4 | – | 14.8 | 21.4 | – | 22.7 | 12.7 | 22.0 | 1.2 | 0.4 |
| Posterior primary branch | 12 | 15.2 | – | 20.4 | 29.7 | – | 31.0 | 17.7 | 30.3 | 1.5 | 0.3 |
| Posterior secondary branch | 12 | 11.1 | – | 15.4 | 21.6 | – | 22.9 | 13.0 | 22.2 | 1.2 | 0.3 |
| CHARACTER | N | RANGE | MEAN | SD | ||
|---|---|---|---|---|---|---|
| Egg bare diameter | 16 | 68.5 | – | 82.0 | 76.5 | 3.5 |
| Egg full diameter | 16 | 90.2 | – | 107.0 | 99.1 | 3.6 |
| Process height | 48 | 10.1 | – | 14.5 | 12.3 | 0.9 |
| Process base width | 48 | 14.6 | – | 19.8 | 16.8 | 1.4 |
| * Process base/height ratio | 48 | 130% | – | 146% | 137% | 5% |
| Inter-process distance | 48 | 4.8 | – | 8.1 | 6.1 | 0.7 |
| * Number of processes on the egg circumference | 16 | 12 | – | 12 | 12.0 | 0.0 |
| CHARACTER | N | RANGE | MEAN | SD | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | ||||||
| Body length | 15 | 369 | – | 633 | – | 483 | – | 96 | – | ||
| Buccopharyngeal tube | |||||||||||
| Buccal tube length | 15 | 41.6 | – | 60.4 | – | 50.2 | – | 6.7 | – | ||
| Stylet support insertion point | 15 | 33.8 | – | 48.1 | 79.3 | – | 81.7 | 40.4 | 80.5 | 5.3 | 0.7 |
| Buccal tube external width | 15 | 8.3 | – | 13.1 | 18.4 | – | 22.1 | 10.4 | 20.6 | 1.5 | 0.8 |
| Buccal tube internal width | 15 | 6.3 | – | 10.0 | 14.0 | – | 16.9 | 7.9 | 15.7 | 1.3 | 0.8 |
| Ventral lamina length | 15 | 26.4 | – | 39.0 | 61.6 | – | 65.4 | 31.9 | 63.6 | 4.3 | 1.3 |
| Placoid lengths | |||||||||||
| Macroplacoid 1 | 15 | 6.3 | – | 11.3 | 15.1 | – | 19.9 | 8.6 | 17.1 | 1.8 | 1.4 |
| Macroplacoid 2 | 15 | 5.4 | – | 9.4 | 12.9 | – | 16.5 | 7.5 | 14.8 | 1.4 | 1.1 |
| Macroplacoid 3 | 15 | 7.9 | – | 12.5 | 18.9 | – | 22.0 | 10.2 | 20.2 | 1.7 | 1.0 |
| Microplacoid | 15 | 3.7 | – | 6.0 | 8.0 | – | 10.1 | 4.6 | 9.1 | 0.8 | 0.6 |
| Macroplacoid row | 15 | 21.4 | – | 35.3 | 51.4 | – | 62.3 | 28.3 | 56.2 | 4.9 | 2.8 |
| Placoid row | 15 | 28.9 | – | 48.0 | 69.5 | – | 81.4 | 37.9 | 75.3 | 6.6 | 3.8 |
| Claw 1 heights | |||||||||||
| External primary branch | 13 | 10.5 | – | 16.8 | 24.3 | – | 29.6 | 12.9 | 26.2 | 2.0 | 1.5 |
| External secondary branch | 11 | 7.8 | – | 13.1 | 18.4 | – | 23.1 | 9.7 | 20.1 | 1.8 | 1.5 |
| Internal primary branch | 13 | 10.3 | – | 16.0 | 22.8 | – | 28.2 | 12.6 | 24.6 | 1.8 | 1.3 |
| Internal secondary branch | 11 | 8.5 | – | 12.4 | 18.3 | – | 21.3 | 10.1 | 19.9 | 1.7 | 1.0 |
| Claw 2 heights | |||||||||||
| External primary branch | 12 | 11.5 | – | 17.8 | 25.1 | – | 31.4 | 13.9 | 27.5 | 2.2 | 1.7 |
| External secondary branch | 11 | 8.8 | – | 13.6 | 20.8 | – | 23.8 | 11.0 | 22.1 | 1.8 | 1.0 |
| Internal primary branch | 12 | 9.8 | – | 15.5 | 22.5 | – | 26.5 | 12.4 | 24.6 | 1.8 | 1.2 |
| Internal secondary branch | 10 | 8.4 | – | 12.5 | 19.5 | – | 21.9 | 10.6 | 21.0 | 1.4 | 0.8 |
| Claw 3 heights | |||||||||||
| External primary branch | 13 | 11.4 | – | 17.6 | 25.3 | – | 31.0 | 13.8 | 27.6 | 1.8 | 1.7 |
| External secondary branch | 12 | 8.5 | – | 14.3 | 19.0 | – | 25.2 | 10.9 | 21.8 | 1.9 | 1.5 |
| Internal primary branch | 12 | 9.8 | – | 15.7 | 22.3 | – | 26.5 | 12.6 | 24.5 | 2.0 | 1.5 |
| Internal secondary branch | 11 | 8.2 | – | 13.4 | 19.5 | – | 23.3 | 10.7 | 20.8 | 1.8 | 1.3 |
| Claw 4 heights | |||||||||||
| Anterior primary branch | 11 | 11.7 | – | 17.6 | 26.8 | – | 30.5 | 14.4 | 28.7 | 1.9 | 1.3 |
| Anterior secondary branch | 10 | 9.0 | – | 13.4 | 20.5 | – | 23.5 | 10.9 | 21.8 | 1.8 | 1.0 |
| Posterior primary branch | 13 | 12.2 | – | 18.7 | 28.7 | – | 32.6 | 15.2 | 30.2 | 2.2 | 1.2 |
| Posterior secondary branch | 12 | 9.3 | – | 15.2 | 20.7 | – | 26.8 | 11.6 | 23.2 | 1.9 | 1.5 |
| CHARACTER | N | RANGE | MEAN | SD | ||
|---|---|---|---|---|---|---|
| Egg bare diameter | 15 | 59.4 | – | 75.0 | 66.2 | 4.9 |
| Egg full diameter | 15 | 84.6 | – | 98.7 | 91.4 | 4.3 |
| Process height | 45 | 11.5 | – | 16.5 | 13.9 | 1.3 |
| Process base width | 45 | 14.3 | – | 19.2 | 16.4 | 1.1 |
| * Process base/height ratio | 45 | 100% | – | 136% | 118% | 11% |
| Inter-process distance | 45 | 3.8 | – | 8.1 | 5.5 | 0.8 |
| * Number of processes on the egg circumference | 15 | 11 | – | 12 | 11.5 | 0.5 |
| CHARACTER | N | RANGE | MEAN | SD | Holotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | µm | pt | ||||||
| *Body length | 9 | 442 | – | 591 | – | 513 | – | 51 | – | 474 | – | ||
| Buccopharyngeal tube | |||||||||||||
| *Buccal tube length | 9 | 54.8 | – | 63.6 | – | 57.9 | – | 2.5 | – | 56.4 | – | ||
| Stylet support insertion point | 5 | 44.7 | – | 47.1 | 79.1 | – | 81.1 | 46.3 | 80.4 | 1.1 | 0.7 | ? | ? |
| *Buccal tube external width | 9 | 12.8 | – | 18.5 | 22.1 | – | 32.1 | 15.0 | 25.9 | 2.1 | 3.3 | 12.8 | 22.7 |
| Buccal tube internal width | 5 | 10.8 | – | 12.2 | 18.7 | – | 21.5 | 11.2 | 19.4 | 0.6 | 1.2 | ? | ? |
| Ventral lamina length | 5 | 33.3 | – | 36.2 | 57.5 | – | 62.8 | 34.6 | 60.1 | 1.3 | 2.1 | ? | ? |
| Placoid lengths | |||||||||||||
| *Macroplacoid 1 | 9 | 7.1 | – | 12.5 | 11.2 | – | 22.0 | 10.3 | 17.8 | 1.7 | 3.3 | 9.8 | 17.4 |
| *Macroplacoid 2 | 9 | 7.6 | – | 10.4 | 13.1 | – | 17.7 | 9.0 | 15.6 | 1.0 | 1.6 | 8.2 | 14.5 |
| Macroplacoid 3 | 5 | 10.6 | – | 11.3 | 18.4 | – | 19.5 | 10.9 | 19.0 | 0.3 | 0.5 | ? | ? |
| Microplacoid | 5 | 4.2 | – | 4.7 | 7.3 | – | 8.1 | 4.4 | 7.6 | 0.2 | 0.4 | ? | ? |
| *Macroplacoid row | 9 | 27.2 | – | 35.2 | 48.2 | – | 60.1 | 31.9 | 55.2 | 2.7 | 5.1 | 27.2 | 48.2 |
| Placoid row | 5 | 40.4 | – | 45.0 | 71.5 | – | 76.7 | 42.7 | 74.2 | 1.8 | 2.3 | ? | ? |
| Claw 1 heights | |||||||||||||
| *External primary branch | 7 | 8.4 | – | 16.8 | 14.5 | – | 29.0 | 12.4 | 21.5 | 3.1 | 5.7 | 10.1 | 17.9 |
| External secondary branch | 3 | 11.3 | – | 12.9 | 19.9 | – | 22.3 | 12.1 | 20.9 | 0.8 | 1.2 | ? | ? |
| Internal primary branch | 3 | 13.6 | – | 14.4 | 23.5 | – | 24.6 | 13.9 | 24.2 | 0.4 | 0.6 | ? | ? |
| Internal secondary branch | 3 | 11.0 | – | 12.3 | 19.5 | – | 21.0 | 11.6 | 20.1 | 0.7 | 0.8 | ? | ? |
| Claw 2 heights | |||||||||||||
| *External primary branch | 8 | 8.6 | – | 17.0 | 15.7 | – | 29.7 | 13.3 | 22.9 | 3.9 | 6.7 | 10.3 | 18.3 |
| External secondary branch | 4 | 12.5 | – | 14.2 | 21.9 | – | 24.6 | 13.4 | 23.2 | 0.8 | 1.3 | ? | ? |
| Internal primary branch | 3 | 15.1 | – | 15.3 | 25.9 | – | 26.1 | 15.2 | 26.0 | 0.1 | 0.1 | ? | ? |
| Internal secondary branch | 2 | 12.9 | – | 13.6 | 22.0 | – | 23.5 | 13.2 | 22.7 | 0.5 | 1.1 | ? | ? |
| Claw 3 heights | |||||||||||||
| *External primary branch | 5 | 15.8 | – | 18.3 | 27.1 | – | 31.1 | 17.2 | 29.9 | 1.0 | 1.7 | ? | ? |
| External secondary branch | 5 | 12.4 | – | 14.2 | 21.4 | – | 24.5 | 13.0 | 22.6 | 0.7 | 1.2 | ? | ? |
| Internal primary branch | 3 | 13.2 | – | 15.3 | 23.3 | – | 26.1 | 14.0 | 24.3 | 1.2 | 1.6 | ? | ? |
| Internal secondary branch | 3 | 10.2 | – | 11.5 | 17.6 | – | 19.8 | 10.9 | 18.7 | 0.7 | 1.1 | ? | ? |
| Claw 4 heights | |||||||||||||
| *Anterior primary branch | 9 | 9.6 | – | 19.5 | 16.6 | – | 33.4 | 15.0 | 26.0 | 3.5 | 6.3 | 10.5 | 18.6 |
| Anterior secondary branch | 5 | 11.4 | – | 14.2 | 19.7 | – | 24.2 | 12.5 | 21.7 | 1.4 | 2.1 | ? | ? |
| Posterior primary branch | 5 | 15.3 | – | 17.9 | 26.9 | – | 30.7 | 16.5 | 28.7 | 1.1 | 1.7 | ? | ? |
| Posterior secondary branch | 5 | 12.1 | – | 13.1 | 21.5 | – | 22.6 | 12.7 | 22.1 | 0.4 | 0.5 | ? | ? |
| CHARACTER | N | RANGE | MEAN | SD | ||
|---|---|---|---|---|---|---|
| ** Egg bare diameter | 18 | 62.1 | – | 82.6 | 70.9 | 5.5 |
| ** Egg full diameter | 18 | 84.1 | – | 105.8 | 92.5 | 6.1 |
| * Process height | 24 | 6.3 | – | 14.3 | 11.3 | 1.8 |
| Process base width | 24 | 14.1 | – | 22.6 | 18.0 | 2.4 |
| *,** Process base/height ratio | 24 | 120% | – | 325% | 164% | 43% |
| Inter-process distance | 9 | 3.4 | – | 11.5 | 6.1 | 2.9 |
| *,** Number of processes on the egg circumference | 18 | 10 | – | 20 | 13.9 | 2.5 |
| CHARACTER | N | RANGE | MEAN | SD | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | ||||||
| Body length | 4 | 522 | – | 675 | – | 589 | – | 77 | – | ||
| Buccopharyngeal tube | |||||||||||
| Buccal tube length | 4 | 52.8 | – | 65.3 | – | 57.7 | – | 5.5 | – | ||
| Stylet support insertion point | 4 | 42.4 | – | 52.7 | 80.3 | – | 81.2 | 46.6 | 80.7 | 4.5 | 0.4 |
| Buccal tube external width | 4 | 12.3 | – | 18.2 | 22.5 | – | 27.9 | 14.3 | 24.6 | 2.8 | 2.4 |
| Buccal tube internal width | 4 | 9.6 | – | 15.5 | 17.6 | – | 23.8 | 11.8 | 20.2 | 2.7 | 2.6 |
| Ventral lamina length | 4 | 33.7 | – | 43.0 | 61.7 | – | 65.9 | 37.2 | 64.3 | 4.4 | 1.9 |
| Placoid lengths | |||||||||||
| Macroplacoid 1 | 4 | 9.1 | – | 14.2 | 17.3 | – | 21.8 | 11.1 | 19.2 | 2.2 | 1.9 |
| Macroplacoid 2 | 4 | 8.6 | – | 12.7 | 14.8 | – | 19.4 | 9.8 | 16.9 | 1.9 | 2.1 |
| Macroplacoid 3 | 4 | 11.2 | – | 13.4 | 20.0 | – | 21.5 | 11.9 | 20.7 | 1.0 | 0.7 |
| Microplacoid | 4 | 4.4 | – | 5.8 | 7.9 | – | 9.2 | 4.9 | 8.6 | 0.6 | 0.6 |
| Macroplacoid row | 4 | 30.3 | – | 41.2 | 57.3 | – | 63.1 | 34.3 | 59.3 | 4.8 | 2.7 |
| Placoid row | 4 | 38.5 | – | 53.8 | 72.6 | – | 82.4 | 44.1 | 76.2 | 6.7 | 4.5 |
| Claw 1 heights | |||||||||||
| External primary branch | 3 | 14.9 | – | 16.4 | 25.1 | – | 27.3 | 15.6 | 26.3 | 0.8 | 1.1 |
| External secondary branch | 3 | 10.4 | – | 11.8 | 18.1 | – | 21.8 | 11.2 | 19.6 | 0.8 | 1.9 |
| Internal primary branch | 4 | 12.7 | – | 14.9 | 22.4 | – | 25.6 | 13.9 | 24.2 | 1.0 | 1.6 |
| Internal secondary branch | 2 | 11.9 | – | 13.7 | 21.0 | – | 22.6 | 12.8 | 21.8 | 1.3 | 1.1 |
| Claw 2 heights | |||||||||||
| External primary branch | 4 | 15.2 | – | 19.3 | 28.1 | – | 29.6 | 16.6 | 28.7 | 1.9 | 0.6 |
| External secondary branch | 3 | 13.1 | – | 17.1 | 22.6 | – | 26.1 | 14.4 | 24.3 | 2.3 | 1.8 |
| Internal primary branch | 4 | 13.0 | – | 16.7 | 24.7 | – | 26.4 | 14.6 | 25.4 | 1.7 | 0.8 |
| Internal secondary branch | 1 | 11.8 | – | 11.8 | 21.7 | – | 21.7 | 11.8 | 21.7 | ? | ? |
| Claw 3 heights | |||||||||||
| External primary branch | 4 | 15.7 | – | 18.8 | 28.8 | – | 30.9 | 17.1 | 29.6 | 1.6 | 1.0 |
| External secondary branch | 3 | 13.4 | – | 16.6 | 25.3 | – | 26.1 | 15.1 | 25.6 | 1.6 | 0.4 |
| Internal primary branch | 4 | 13.1 | – | 15.9 | 24.2 | – | 25.8 | 14.3 | 24.8 | 1.4 | 0.7 |
| Internal secondary branch | 0 | ? | ? | ? | ? | ? | ? | ||||
| Claw 4 heights | |||||||||||
| Anterior primary branch | 2 | 16.3 | – | 16.9 | 28.0 | – | 31.0 | 16.6 | 29.5 | 0.4 | 2.1 |
| Anterior secondary branch | 1 | 13.0 | – | 13.0 | 22.4 | – | 22.4 | 13.0 | 22.4 | ? | ? |
| Posterior primary branch | 3 | 15.3 | – | 17.4 | 26.7 | – | 29.5 | 16.6 | 28.1 | 1.2 | 1.4 |
| Posterior secondary branch | 1 | 12.7 | – | 12.7 | 23.2 | – | 23.2 | 12.7 | 23.2 | ? | ? |
| CHARACTER | N | RANGE | MEAN | SD | ||
|---|---|---|---|---|---|---|
| Egg bare diameter | 1 | 67.0 | – | 67.0 | 67.0 | – |
| Egg full diameter | 1 | 90.2 | – | 90.2 | 90.2 | – |
| Process height | 3 | 11.0 | – | 17.3 | 13.3 | 3.5 |
| Process base width | 3 | 15.8 | – | 20.8 | 18.7 | 2.6 |
| * Process base/height ratio | 3 | 112% | – | 180% | 145% | 34% |
| Inter-process distance | 3 | 4.0 | – | 5.0 | 4.4 | 0.5 |
| * Number of processes on the egg circumference | 1 | 11 | – | 11 | 11.0 | – |
| H1 | H2 | H3 | H4 | H5 | H6 | H7 | H8 | H9 | H10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| H1 | 0.003 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | |
| H2 | 0.005 | 0.003 | 0.002 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | |
| H3 | 0.003 | 0.005 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | |
| H4 | 0.002 | 0.003 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | |
| H5 | 0.003 | 0.005 | 0.003 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | 0.002 | |
| H6 | 0.002 | 0.005 | 0.003 | 0.002 | 0.003 | 0.002 | 0.002 | 0.002 | 0.002 | |
| H7 | 0.003 | 0.005 | 0.003 | 0.002 | 0.003 | 0.003 | 0.002 | 0.002 | 0.002 | |
| H8 | 0.003 | 0.005 | 0.003 | 0.002 | 0.003 | 0.003 | 0.003 | 0.002 | 0.002 | |
| H9 | 0.003 | 0.005 | 0.003 | 0.002 | 0.003 | 0.003 | 0.003 | 0.003 | 0.002 | |
| H10 | 0.003 | 0.005 | 0.003 | 0.002 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaczmarek, Ł.; Mioduchowska, M.; Kačarević, U.; Kubska, K.; Parnikoza, I.; Gołdyn, B.; Roszkowska, M. New Records of Antarctic Tardigrada with Comments on Interpopulation Variability of the Paramacrobiotus fairbanksi Schill, Förster, Dandekar and Wolf, 2010. Diversity 2020, 12, 108. https://doi.org/10.3390/d12030108
Kaczmarek Ł, Mioduchowska M, Kačarević U, Kubska K, Parnikoza I, Gołdyn B, Roszkowska M. New Records of Antarctic Tardigrada with Comments on Interpopulation Variability of the Paramacrobiotus fairbanksi Schill, Förster, Dandekar and Wolf, 2010. Diversity. 2020; 12(3):108. https://doi.org/10.3390/d12030108
Chicago/Turabian StyleKaczmarek, Łukasz, Monika Mioduchowska, Uroš Kačarević, Katarzyna Kubska, Ivan Parnikoza, Bartłomiej Gołdyn, and Milena Roszkowska. 2020. "New Records of Antarctic Tardigrada with Comments on Interpopulation Variability of the Paramacrobiotus fairbanksi Schill, Förster, Dandekar and Wolf, 2010" Diversity 12, no. 3: 108. https://doi.org/10.3390/d12030108
APA StyleKaczmarek, Ł., Mioduchowska, M., Kačarević, U., Kubska, K., Parnikoza, I., Gołdyn, B., & Roszkowska, M. (2020). New Records of Antarctic Tardigrada with Comments on Interpopulation Variability of the Paramacrobiotus fairbanksi Schill, Förster, Dandekar and Wolf, 2010. Diversity, 12(3), 108. https://doi.org/10.3390/d12030108