Functional Diversity Changes after Selective Thinning in a Tropical Mountain Forest in Southern Ecuador

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
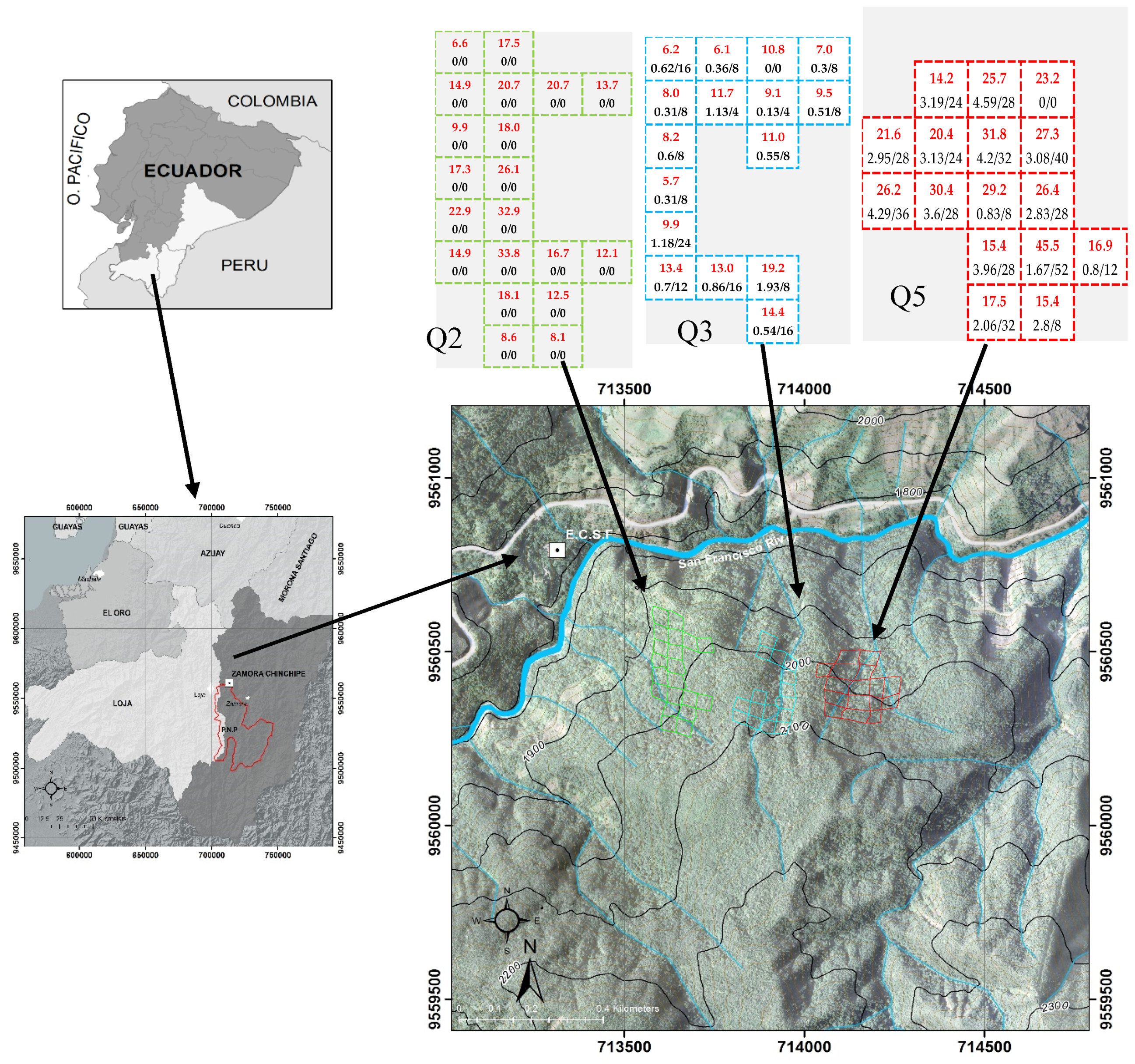
2.1. Study Area
2.2. Plot Installation
2.3. Selective Thinning and Intensity
2.4. Assessing Impact of Selective Thinning on Functional Diversity
2.5. Data Acquisition
2.6. Statistical Analysis
- -
- Functional Richness (FRic), which represents the amount of space occupied by a community, independent from the relative abundances of the species [70].
- -
- Functional Divergence (FDiv) measures how species of a community diverge in their distance from the center of gravity in the functional space, and determines the relative abundance of a species within its functional range, [70,71]. FDiv provides additional information for FRic, because this index indicates which species has the most extreme feature combinations (i.e., specialized species), and which ones are generalist [72].
- -
- Functional Evenness (FEve) quantifies the regularity of how species abundances are distributed within their functional range, by correlating abundance with average distance between different species [57]. FEve can be related to the uniformity of species (Pielou’s J) and the FDiv because it determines if the extent of functional distances between the species is regular [41].
- -
- Functional Dispersion (FDis) gives the mean distance of an individual species relative to the centroid of the community, which depicts its mean distance relative to all species in the community, and therefore, accounts for relative abundances of a species.
- -
- Rao Quadratic Entropy (RaoQ) correlates the abundances of different species [73] to improve the FD index. An adverse property of RaoQ is that its value may decrease if species richness increases, because abundances and dissimilarity between the species are considered. Therefore, the introduction or loss of species in a community increases or decreases the species abundance, but, at the same time, may reduce the dissimilarity between the species.
- -
- Community weighted mean (CWM) represents the sum of each species trait value weighted by its relative abundance in the community [74]. To characterize the community structure from a functional point of view, we used the community weighted mean, which is calculated combining the species abundance with the trait values of the given species [75]. This describes the trait averages over a community [76] and reflects the dominant trait in a given community [33,77,78].
3. Results
3.1. Selective Thinning and Structure
3.2. Floristic Composition
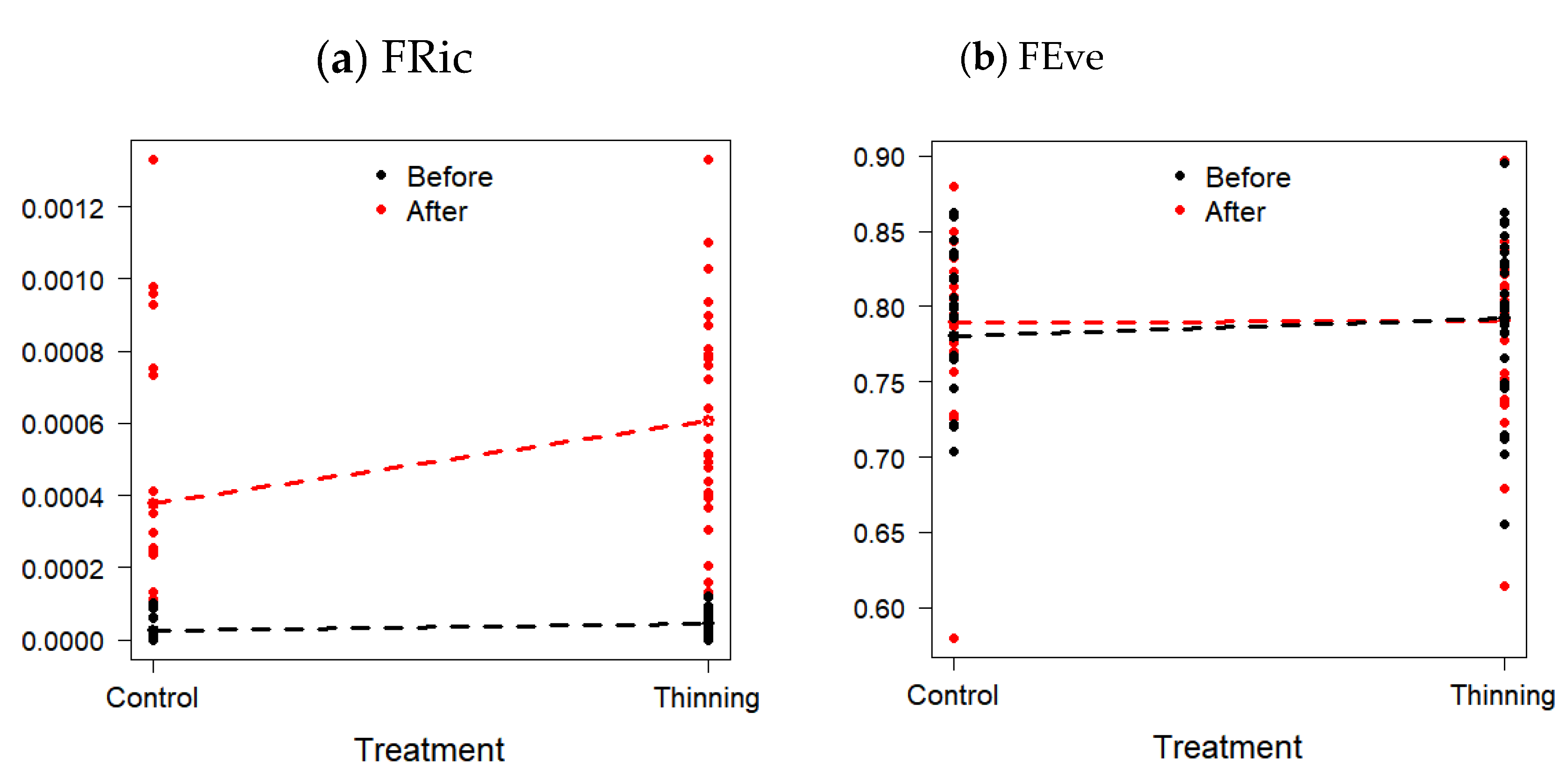
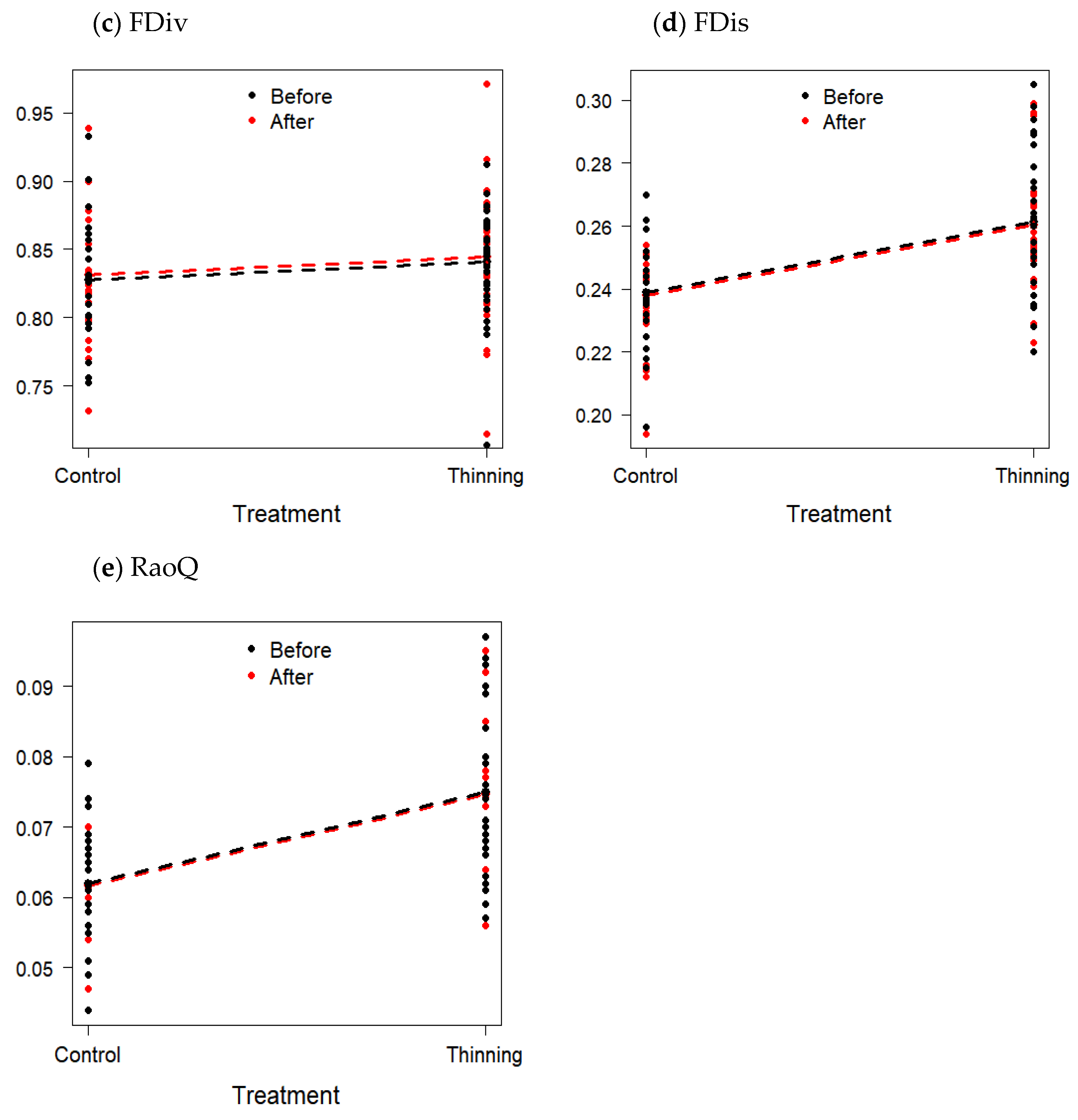
3.3. Changes in Functional Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mosandl, R.; Günter, S.; Stimm, B.; Weber, M. Ecuador suffers the highest deforestation rate in South America. In Gradients in a Tropical Mountain Ecosystem of Ecuador; Beck, E., Bendix, J., Kottke, I., Makeschin, F., Mosandl, R., Eds.; Springer: Berlin, Germany, 2008; pp. 347–359. ISBN 978-3-540-73526-7. [Google Scholar]
- Benítez, A.; Aragón, G.; González, Y.; Prieto, M. Functional traits of epiphytic lichens in response to forest disturbance and as predictors of total richness and diversity. Ecol. Indic. 2018, 86, 18–26. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Gardner, T.A.; Barlow, J.; Chazdon, R.; Ewers, R.M.; Harvey, C.A.; Peres, C.A.; Sodhi, N.S. Prospects for tropical forest biodiversity in a human-modified world. Ecol. Lett. 2009, 12, 561–582. [Google Scholar] [CrossRef] [Green Version]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Peres, C.A.; Bradshaw, C.J.A.; Laurance, W.F.; Lovejoy, T.E.; et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2011, 478, 378–381. [Google Scholar] [CrossRef]
- Dirzo, R.; Raven, P.H. Global state of biodiversity and loss. Annu. Rev. Environ. Resour. 2003, 28, 137–167. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.L. Beyond deforestation: Restoring forests and ecosystem services on degraded lands. Science 2008, 320, 1458–1460. [Google Scholar] [CrossRef] [Green Version]
- Carreño-Rocabado, G.; Peña-Claros, M.; Bongers, F.; Alarcón, A.; Licona, J.C.; Poorter, L. Effects of disturbance intensity on species and functional diversity in a tropical forest. J. Ecol. 2012, 100, 1453–1463. [Google Scholar] [CrossRef]
- Pessoa, M.S.; Hambuckers, A.; Benchimol, M.; Rocha-Santos, L.; Bomfim, J.A.; Faria, D.; Cazetta, E. Deforestation drives functional diversity and fruit quality changes in a tropical tree assemblage. Perspect. Plant Ecol. Evol. Syst. 2017, 28, 78–86. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global Consequences of Land Use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Putz, F.E.; Blate, G.M.; Redford, K.H.; Fimbel, R.; Robinson, J. Tropical forest management and conservation of biodiversity: An overview. Conserv. Biol. 2001, 15, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Kusumoto, B.; Shiono, T.; Miyoshi, M.; Maeshiro, R.; Fujii, S.; Kuuluvainen, T.; Kubota, Y. Functional response of plant communities to clearcutting: Management impacts differ between forest vegetation zones. J. Appl. Ecol. 2014, 52, 171–180. [Google Scholar] [CrossRef]
- Günter, S.; Weber, M.; Erreis, R.; Aguirre, N. Influence of distance to forest edges on natural regeneration of abandoned pastures—A case study in the tropical mountain rain forest of Southern Ecuador. Eur. J. For. Res. 2006, 126, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Edwards, D.P.; Tobias, J.A.; Sheil, D.; Meijaard, E.; Laurance, W.F. Maintaining ecosystem function and services in logged tropical forests. Trends Ecol. Evol. 2014, 29, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Darrigo, M.R.; Venticinque, E.M.; dos Santos, F.A.M. Effects of reduced impact logging on the forest regeneration in the central Amazonia. Forest Ecol. Manag. 2016, 360, 52–59. [Google Scholar] [CrossRef]
- Wadsworth, F.H. Forest Production for Tropical America (No. 710); US Department of Agriculture, Forest Service: Washington, WA, USA, 1997.
- Dawkins, H.C. The refining of mixed forest: A new objective for tropical silviculture. Emp. For. Rev. 1955, 34, 188–191. [Google Scholar]
- Louman, B.; Valerio, J.; Jiménez, W. Bases ecológicas. Silvicultura de bosques latifoliados húmedos con énfasis en América Central. Silvicultura de Bosques Latifoliados Húmedos con Énfasis en América Central. Serie Técnica. Manual Técnico. 2001, 46, 19–78. [Google Scholar]
- De Avila, A.L.; Van Der Sande, M.T.; Dormann, C.F.; Peña-Claros, M.; Poorter, L.; Mazzei, L.; Ruschel, A.R.; Silva, J.N.M.; De Carvalho, J.O.P.; Bauhus, J. Disturbance intensity is a stronger driver of biomass recovery than remaining tree-community attributes in a managed Amazonian forest. J. Appl. Ecol. 2018, 55, 1647–1657. [Google Scholar] [CrossRef] [Green Version]
- Yguel, B.; Piponiot, C.; Mirabel, A.; Dourdain, A.; Hérault, B.; Gourlet-Fleury, S.; Forget, P.-M.; Fontaine, C. Beyond species richness and biomass: Impact of selective logging and silvicultural treatments on the functional composition of a neotropical forest. For. Ecol. Manag. 2019, 433, 528–534. [Google Scholar] [CrossRef]
- Bonilla-Bedoya, S.; Estrella-Bastidas, A.; Ordoñez, M.; Sánchez, A.; Herrera, M.A. Patterns of timber harvesting and its relationship with sustainable forest management in the Western Amazon, Ecuador case. J. Sustain. For. 2017, 36, 433–453. [Google Scholar] [CrossRef]
- Jadán, O.; Cifuentes Jara, M.; Torres, B.; Selesi, D.; Veintimilla Ramos, D.A.; Günter, S. Influence of tree cover on diversity, carbon sequestration and productivity of cocoa systems in the Ecuadorian Amazon. Bois. For. Trop. 2015, 325, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Middendorp, R.S.; Pérez, A.J.; Molina, A.; Lambin, E.F. The potential to restore native woody plant richness and composition in a reforesting landscape: A modeling approach in the Ecuadorian Andes. Landsc. Ecol. 2016, 31, 1581–1599. [Google Scholar] [CrossRef]
- Chirino, E.; Ruiz-Yanetti, S.; Vilagrosa, A.; Mera, X.; Espinoza, M.; Lozano, P. Morpho-functional traits and plant response to drought conditions in seedlings of six native species of Ecuadorian Ecosystems. Flora 2017, 233, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Aguilar, M.J.; Konold, W.; Schmitt, C.B. Community mapping of ecosystem services in tropical rainforest of Ecuador. Ecol. Indic. 2017, 73, 460–471. [Google Scholar] [CrossRef]
- Raes, L.; D’Haese, M.; Aguirre, N.; Knoke, T. A portfolio analysis of incentive programmes for conservation, restoration and timber plantations in Southern Ecuador. Land Use Policy 2016, 51, 244–259. [Google Scholar] [CrossRef]
- Hayes, T.; Murtinho, F.; Camacho, L.M.C.; Crespo, P.; McHugh, S.; Salmerón, D. Can conservation contracts co-exist with change? Payment for ecosystem services in the context of adaptive decision-making and sustainability. Environ. Manag. 2015, 55, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Wunder, S. Are Direct Payments for Environmental Services Spelling Doom for Sustainable Forest Management in the Tropics? Ecol. Soc. 2006, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- MAE 2000. Estrategia Para el Desarrollo Forestal Sustentable del Ecuador; MAE 2000: Quito, Ecuador, 2000. [Google Scholar]
- Bricca, A.; Chelli, S.; Canullo, R.; Cutini, M. The Legacy of the Past Logging: How Forest Structure Affects Different Facets of Understory Plant Diversity in Abandoned Coppice Forests. Diversity 2020, 12, 109. [Google Scholar] [CrossRef] [Green Version]
- Tonteri, T.; Salemaa, M.; Rautio, P.; Hallikainen, V.; Korpela, L.; Merilä, P. Forest management regulates temporal change in the cover of boreal plant species. For. Ecol. Manag. 2016, 381, 115–124. [Google Scholar] [CrossRef]
- Schleuter, D.; Daufresne, M.; Massol, F.; Argillier, C. A user’s guide to functional diversity indices. Ecol. Monogr. 2010, 80, 469–484. [Google Scholar] [CrossRef] [Green Version]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional. Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity: Back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Báez, S.; Homeier, J. Functional traits determine tree growth and ecosystem productivity of a tropical montane forest: Insights from a long-term nutrient manipulation experiment. Glob. Chang. Biol. 2018, 24, 399–409. [Google Scholar] [CrossRef]
- Maire, E.; Grenouillet, G.; Brosse, S.; Villéger, S. How many dimensions are needed to accurately assess functional diversity? A pragmatic approach for assessing the quality of functional spaces. Glob. Ecol. Biogeogr. 2015, 24, 728–740. [Google Scholar] [CrossRef]
- Bendix, J.; Trachte, K.; Cermak, J.; Rollenbeck, R.; Nauß, T. Formation of convective clouds at the foothills of the tropical eastern Andes (South Ecuador). J. Appl. Meteorol. Clim. 2009, 48, 1682–1695. [Google Scholar] [CrossRef]
- Gonzalez-Jaramillo, V.; Fries, A.; Rollenbeck, R.; Paladines, J.; Oñate-Valdivieso, F.; Bendix, J. Assessment of deforestation during the last decades in Ecuador using NOAA-AVHRR satellite data. Erdkunde 2016, 217–235. [Google Scholar] [CrossRef]
- Berry, N.J.; Phillips, O.L.; Ong, R.C.; Hamer, K.C. Impacts of selective logging on tree diversity across a rainforest landscape: The importance of spatial scale. Landsc. Ecol. 2008, 23, 915–929. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U. Disturbance effects on species diversity and functional diversity in riparian and upland plant communities. Ecology 2010, 91, 28–35. [Google Scholar] [CrossRef]
- Baraloto, C.; Herault, B.; Paine, C.E.; Massot, H.; Blanc, L.; Bonal, D.; Sabatier, D. Contrasting taxonomic and functional responses of a tropical tree community to selective logging. J. Appl. Ecol. 2012, 49, 861–870. [Google Scholar] [CrossRef]
- Clark, J.A.; Covey, K.R. Tree species richness and the logging of natural forests: A meta-analysis. For. Ecol. Manag. 2012, 276, 146–153. [Google Scholar] [CrossRef]
- Chaudhary, A.; Burivalova, Z.; Koh, L.P.; Hellweg, S. Impact of forest management on species richness: Global meta-analysis and economic trade-offs. Sci. Rep. 2016, 6, 23954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohl, C.; Bussmann, R. Recolonisation of natural landslides in tropical mountain forests of Southern Ecuador. Feddes Repert. 2004, 115, 248–264. [Google Scholar] [CrossRef]
- Brehm, G.; Homeier, J.; Fiedler, K. Beta diversity of geometrid moths (Lepidoptera: Geometridae) in an Andean montane rainforest. Divers. Distrib. 2003, 9, 351–366. [Google Scholar] [CrossRef]
- Fernández, G.F.C.; Obermeier, W.; Gerique, A.; López-Sandoval, M.; Lehnert, L.W.; Thies, B.; Bendix, J. Land Cover Change in the Andes of Southern Ecuador—Patterns and Drivers. Remote Sens. 2015, 7, 2509–2542. [Google Scholar] [CrossRef] [Green Version]
- González-Jaramillo, V.; Fries, A.; Zeilinger, J.; Homeier, J.; Paladines-Benitez, J.; Bendix, J. Estimation of above ground biomass in a tropical mountain forest in southern Ecuador using airborne LIDAR data. Rem. Sens. 2018, 10, 660. [Google Scholar] [CrossRef] [Green Version]
- Moser, G.; Leuschner, C.; Hertel, D.; Graefe, S.; Soethe, N.; Iost, S. Elevation effects on the carbon budget of tropical mountain forests (S Ecuador): The role of the belowground compartment. Glob. Chang. Biol. 2011, 17, 2211–2226. [Google Scholar] [CrossRef] [Green Version]
- Dislich, C.; Huth, A. Modelling the impact of shallow landslides on forest structure in tropical montane forests. Ecol. Model. 2012, 239, 40–53. [Google Scholar] [CrossRef]
- Cabrera, O.; Fries, A.; Hildebrandt, P.; Günter, S.; Mosandl, R. Early Growth Response of Nine Timber Species to Release in a Tropical Mountain Forest of Southern Ecuador. Forests 2019, 10, 254. [Google Scholar] [CrossRef] [Green Version]
- Paulsch, A. Development and Application of a Classification System for Undisturbed and Disturbed Tropical Montane Forests Based on Vegetation Structure. Ph.D. Thesis, University of Bayreuth, Bayreuth, Germany, 2002. [Google Scholar]
- Homeier, J.; Breckle, S.W.; Günter, S.; Rollenbeck, R.T.; Leuschner, C. Tree Diversity, Forest Structure and Productivity along Altitudinal and Topographical Gradients in a Species-Rich Ecuadorian Montane Rain Forest. Biotropica 2010, 42, 140–148. [Google Scholar] [CrossRef]
- Andresen, E.; Pedroza-Espino, L.; Allen, E.B.; Pérez-Salicrup, D.R. Effects of Selective Vegetation Thinning on Seed Removal in Secondary Forest Succession. Biotropica 2005, 37, 145–148. [Google Scholar] [CrossRef]
- De Graaf, N.R.; Poels, R.L.H.; Van Rompaey, R.S.A.R. Effect of silvicultural treatment on growth and mortality of rainforest in Surinam over long periods. For. Ecol. Manag. 1999, 124, 123–135. [Google Scholar] [CrossRef]
- Peña-Claros, M.; Peters, E.M.; Justiniano, M.J.; Bongers, F.; Blate, G.M.; Fredericksen, T.S.; Putz, F.E. Regeneration of commercial tree species following silvicultural treatments in a moist tropical forest. For. Ecol. Manag. 2008, 255, 1283–1293. [Google Scholar] [CrossRef]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Chave, J. Global Wood Density Database. 2009. Available online: https://dryad.figshare.com/articles/Global_Wood_Density_Database/4172847 (accessed on 2 April 2009).
- Chave, J.; Coomes, D.A.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- MAE (Ministerio del Ambiente del Ecuador); FAO (Organización de las Naciones Unidas para la Alimentación y la Agricultura, IT). Propiedades Anatómicas, Físicas y Mecánicas de 93 Especies Forestales; MAE: Quito, Ecuador, 2014; p. 105. [Google Scholar]
- Palacios, W.; Jaramillo, N. Gremios ecológicos forestales del noroccidente del Ecuador: Implicaciones en el manejo del bosque nativo. Lyonia 2004, 6, 55–75. [Google Scholar]
- Osazuwa-Peters, O.L.; Jiménez, I.; Oberle, B.; Chapman, C.A.; Zanne, A.E. Selective logging: Do rates of forest turnover in stems, species composition and functional traits decrease with time since disturbance?–A 45 year perspective. For. Ecol. Manag. 2015, 357, 10–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, V.A.H.F.; Ferreira, M.J. Are photosynthetic leaf traits related to the first-year growth of tropical tree seedlings? A light-induced plasticity test in a secondary forest enrichment planting. For. Ecol. Manag. 2020, 460, 117900. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, J.; Xiao, H.; Liu, J.; Wang, L. Variations in leaf functional traits among plant species grouped by growth and leaf types in Zhenjiang, China. J. For. Res. 2017, 28, 241–248. [Google Scholar] [CrossRef]
- Xu, F.; Guo, W.; Xu, W.; Wei, Y.; Wang, R. Leaf morphology correlates with water and light availability: What consequences for simple and compound leaves? Prog. Nat. Sci. 2009, 19, 1789–1798. [Google Scholar] [CrossRef]
- Jara-Guerrero, A.; De la Cruz, M.; Méndez, M. Seed dispersal spectrum of woody species in south Ecuadorian dry forests: Environmental correlates and the effect of considering species abundance. Biotropica 2011, 43, 722–730. [Google Scholar] [CrossRef] [Green Version]
- Seidler, T.G.; Plotkin, J.B. Seed dispersal and spatial pattern in tropical trees. PLoS Biol. 2006, 4, e344. [Google Scholar] [CrossRef]
- Cortés-Flores, J.; Cornejo-Tenorio, G.; Urrea-Galeano, L.A.; Andresen, E.; González-Rodríguez, A.; Ibarra-Manríquez, G. Phylogeny, fruit traits, and ecological correlates of fruiting phenology in a Neotropical dry forest. Oecologia 2019, 189, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Günter, S.; Cabrera, O.; Weber, M.; Stimm, B.; Zimmermann, M.; Fiedler, K.; Knuth, J.; Boy, J.; Wilcke, W.; Iost, S.; et al. Natural Forest Management in Neotropical Mountain Rain Forests—An Ecological Experiment. In Gradients in a Tropical Mountain Ecosystem of Ecuador; Beck, E., Bendix, J., Kottke, I., Makeschin, F., Mosandl, R., Eds.; Springer: Berlin, Germany, 2008; pp. 347–359. ISBN 978-3-540-73526-7. [Google Scholar]
- Laliberte, A.E.; Legendre, P.; Shipley, B.; Laliberté, M.E. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version 1.0-12. 2010. Available online: https://www.semanticscholar.org/paper/Package-%27fd%27-Title-Measuring-Functional-Diversity-Lalibert%C3%A9-Shipley/99009d911cb83b2bd2bc1ea6e7399b0d05e5cb63 (accessed on 16 April 2010).
- Mason, N.W.; Bello, F.; Mouillot, D.; Pavoine, S.; Dray, S. A guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. J. Veget. Sci. 2013, 24, 794–806. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Mouchet, M.A.; Villeger, S.; Mason, N.W.H.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Botta-Dukat, Z. Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.; Garden, D.; Dorrough, J.; Bonis, A. Assessing functional diversity in the field–methodology matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Ricotta, C.; Moretti, M. CWM and Rao’s quadratic diversity: A unified framework for functional ecology. Oecologia 2011, 167, 181–188. [Google Scholar] [CrossRef]
- Bello, F.D.; Lepš, J.; Lavorel, S.; Moretti, M. Importance of species abundance for assessment of trait composition: An example based on pollinator communities. Community Ecol. 2007, 8, 163–170. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billes, G.; Navas, M.L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Lepsˇ, J.; de Bello, F.; Lavorel, S.; Berman, S. Quantifying and interpreting functional diversity of natural communities: Practical considerations matter. Preslia 2006, 78, 481–501. [Google Scholar]
- Bates, D.; Sarkar, D.; Bates, M.D.; Matrix, L. The lme4 package. R Package Version 2007, 2, 74. [Google Scholar]
- Hothorn, T.; Zeileis, A.; Farebrother, R.W.; Cummins, C.; Millo, G.; Mitchell, D.; Zeileis, M.A. Package ‘Lmtest’; Project for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Costa, F.R.; Magnusson, W.E. Effects of Selective Logging on the Diversity and Abundance of Flowering and Fruiting Understory Plants in a Central Amazonian Forest. Biotropica 2003, 35, 103–114. [Google Scholar] [CrossRef]
- Bawa, K.S.; Seidler, R. Natural forest management and conservation of biodiversity in tropical forests. Conserv. Biol. 1998, 12, 46–55. [Google Scholar] [CrossRef]
- Lohbeck, M.; Poorter, L.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Paz, H.; Bongers, F. Successional changes in functional composition contrast for dry and wet tropical forest. Ecology 2013, 94, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- De Avila, A.L.; Ruschel, A.R.; de Carvalho, J.O.P.; Mazzei, L.; Silva, J.N.M.; do Carmo Lopes, J.; Bauhus, J. Medium-term dynamics of tree species composition in response to silvicultural intervention intensities in a tropical rain forest. Biol. Conserv. 2015, 191, 577–586. [Google Scholar] [CrossRef]
- Nöske, N.M.; Hilt, N.; Werner, F.A.; Brehm, G.; Fiedler, K.; Sipman, H.J.; Gradstein, S.R. Disturbance effects on diversity of epiphytes and moths in a montane forest in Ecuador. Basic Appl. Ecol. 2008, 9, 4–12. [Google Scholar] [CrossRef]
- Werner, F.A. Reduced growth and survival of vascular epiphytes on isolated remnant trees in a recent tropical montane forest clear-cut. Basic. Appl. Ecol. 2011, 12, 172–181. [Google Scholar] [CrossRef]
- Lopez-Iglesias, B.; Villar, R.; Poorter, L. Functional traits predict drought performance and distribution of Mediterranean woody species. Acta Oecologica 2014, 56, 10–18. [Google Scholar] [CrossRef]
- Lawton, J.H.; Bignell, D.E.; Bolton, B.; Bloemers, G.F.; Eggleton, P.; Hammond, P.M.; Stork, N.E. Biodiversity inventories, indicator taxa and effects of habitat modification in tropical forest. Nature 1998, 391, 72. [Google Scholar] [CrossRef]
- Huang, W.; Pohjonen, V.; Johansson, S.; Nashanda, M.; Katigula, M.I.L.; Luukkanen, O. Species diversity, forest structure and species composition in Tanzanian tropical forests. For. Ecol. Manag. 2003, 173, 11–24. [Google Scholar] [CrossRef]
- Putz, F.E.; Zuidema, P.A.; Synnott, T.; Peña-Claros, M.; Pinard, M.A.; Sheil, D.; Vanclay, J.; Sist, P.; Gourlet-Fleury, S.; Griscom, B.; et al. Sustaining conservation values in selectively logged tropical forests: The attained and the attainable. Conserv. Lett. 2012, 5, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Curzon, M.T.; D’Amato, A.W.; Fraver, S.; Palik, B.J.; Bottero, A.; Foster, J.R.; Gleason, K.E. Harvesting influences functional identity and diversity over time in forests of the northeastern USA. For. Ecol. Manag. 2017, 400, 93–99. [Google Scholar] [CrossRef]
- Mori, A.S.; Lertzman, K.P.; Gustafsson, L. Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest ecology. J. App. Ecol. 2017, 54, 12–27. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Functional Trait | Type | Categories | Unit | Code |
|---|---|---|---|---|
| Wood density | Acquisitive | - | g cm3 | WD |
| Stem Density | Acquisitive | - | cm | DBH |
| Diametric growth | Acquisitive | - | cm year−1 | G |
| Leaf type | Acquisitive | Simple | LT | |
| Compound | ||||
| Ecological guild | Conservative | Shade tolerant | ST | |
| Light Tolerant | LT | |||
| Partial light Tolerant | PLT | |||
| Partial Shade Tolerant | PST | |||
| Dispersion syndrome | Conservative | Anemochory | AN | |
| Zoochory | ZOC | |||
| Barochory | BAR |
| Predictors | Functional Richness: FRic | Functional Diversity: FDiv | Functional Dispersion: FDis | Rao Quadratic Entropy: RaoQ | ||||
|---|---|---|---|---|---|---|---|---|
| LTR p Value <0.001 | LTR p Value <0.001 | LTR p Value <0.001 | LRT p Value 0.01 | |||||
| Coeff | p-Value | Coeff | p-Value | Coeff | p-Value | Coeff | p-Value | |
| (Intercept) | −1.08 | <0001 | 8.6 | <0001 | 4.3 | <0001 | 2.06 | <0001 |
| Stem density | 5.2 | 0.02 | ||||||
| Total Species | ||||||||
| Thinning Intensity | 2.47 | 0.0182 | 4.04 | 0.004 | ||||
| Altitude | −7.31 | 0.001 | 12.9 | 0.005 | 14.4 | 0.0002 | ||
| Treatment | ||||||||
| Period | −7.3 | <0001 | ||||||
| Treatment: Period | −1.89 | 0.0270 | ||||||
| INDEX | Before | After | Coeff | F p-Value |
|---|---|---|---|---|
| Functional Richness (FRic) | 0.000025 ± 0.000003 | 0.0005 ± 0.00003 | −9.03 | <0001 |
| Functional Evenness (FEve) | 0.786 ± 0.061 | 0.791 ± 0.058 | −0.26 | 0.7902 |
| Functional Divergence (FDiv) | 0.835 ± 0.041 | 0.838 ± 0.047 | −0.41 | 0.6793 |
| Functional Dispersion (FDis) | 0.251 ± 0.022 | 0.263 ± 0.023 | 0.23 | 0.3246 |
| Quadratic Entropy (RaoQ) | 0.0696 ± 0.011 | 0.0691 ± 0.012 | 0.11 | 0.8919 |
| Predictors | FREQUENCY | GROWTH | DBH | |||
|---|---|---|---|---|---|---|
| LRT p Value 0.01 | LRT p Value <0.0001 | LRT p Value 0.01 | ||||
| Coeff | p-Value | Coeff | p-Value | Coeff | p-Value | |
| Stem density | 1.89 | 0.002 | 1.04 | 0.004 | ||
| Total Species | 1.8 | <0001 | ||||
| Thinning Intensity | 3.47 | <0001 | ||||
| Altitude | −0.98 | 0.003 | −2.43 | 0.002 | ||
| Treatment | 1.5 | <0001 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabrera, O.; Hildebrandt, P.; Stimm, B.; Günter, S.; Fries, A.; Mosandl, R. Functional Diversity Changes after Selective Thinning in a Tropical Mountain Forest in Southern Ecuador. Diversity 2020, 12, 256. https://doi.org/10.3390/d12060256
Cabrera O, Hildebrandt P, Stimm B, Günter S, Fries A, Mosandl R. Functional Diversity Changes after Selective Thinning in a Tropical Mountain Forest in Southern Ecuador. Diversity. 2020; 12(6):256. https://doi.org/10.3390/d12060256
Chicago/Turabian StyleCabrera, Omar, Patrick Hildebrandt, Bernd Stimm, Sven Günter, Andreas Fries, and Reinhard Mosandl. 2020. "Functional Diversity Changes after Selective Thinning in a Tropical Mountain Forest in Southern Ecuador" Diversity 12, no. 6: 256. https://doi.org/10.3390/d12060256
APA StyleCabrera, O., Hildebrandt, P., Stimm, B., Günter, S., Fries, A., & Mosandl, R. (2020). Functional Diversity Changes after Selective Thinning in a Tropical Mountain Forest in Southern Ecuador. Diversity, 12(6), 256. https://doi.org/10.3390/d12060256