An Overview of Subterranean Biodiversity Hotspots





1. Introduction
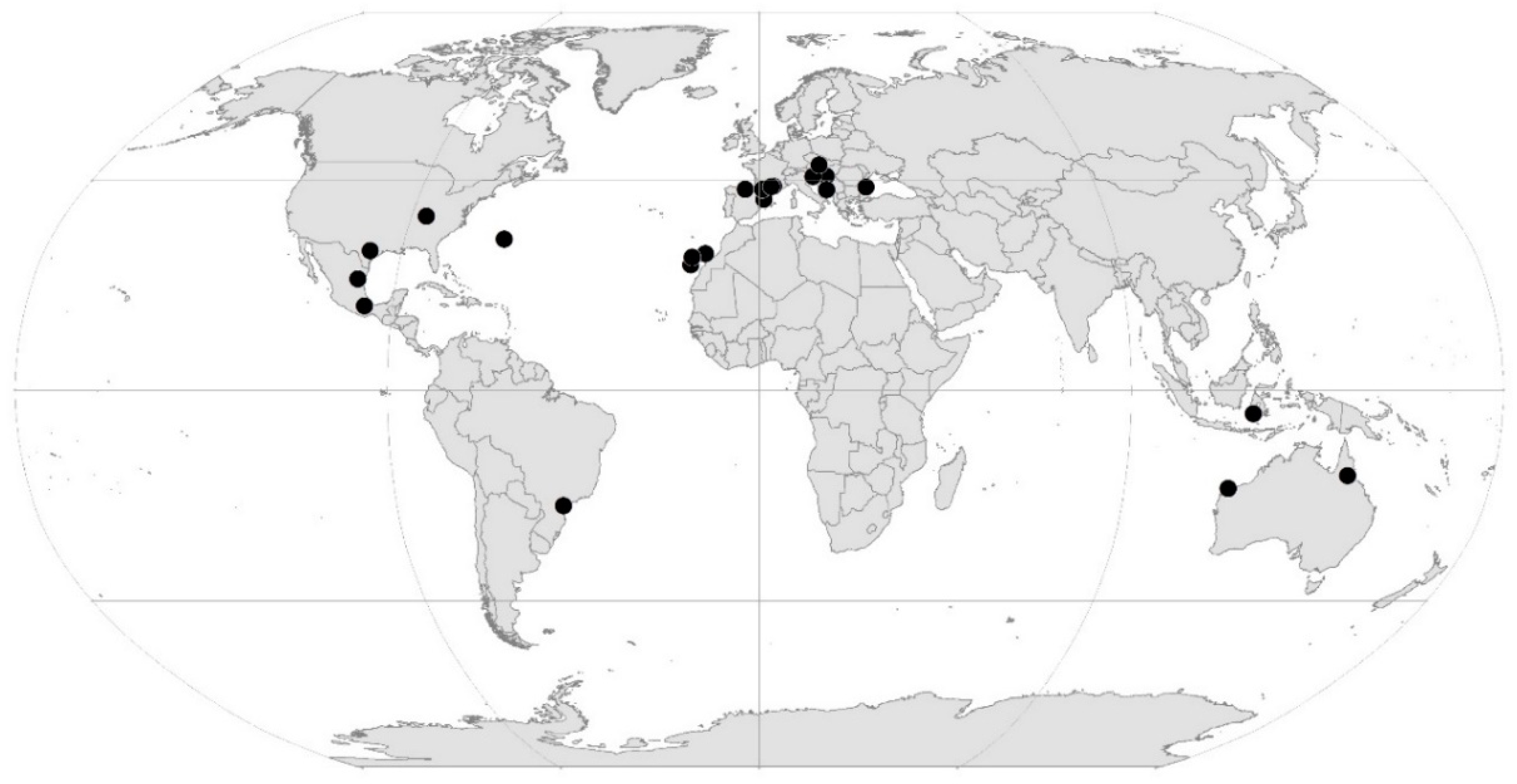
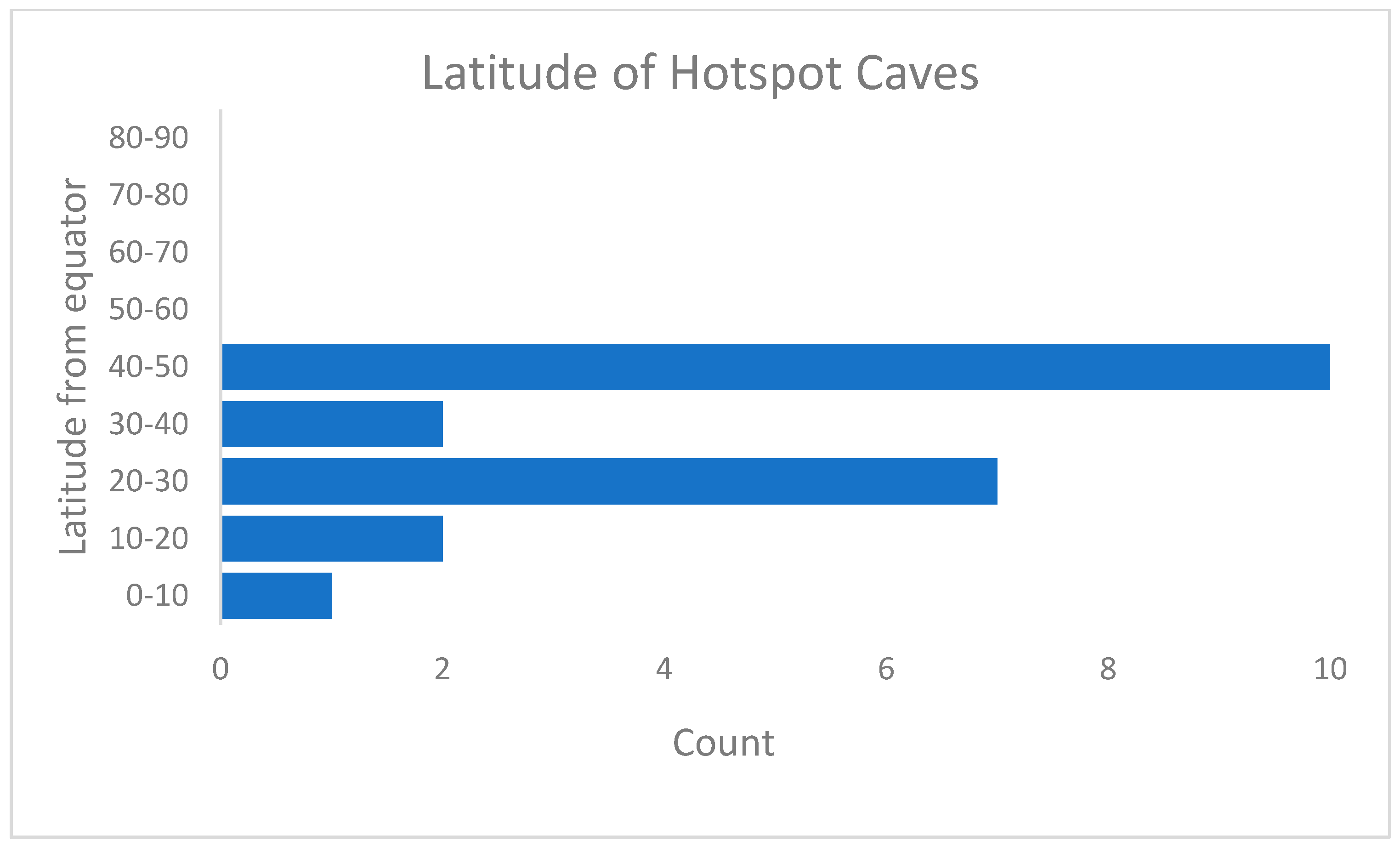
2. Overview of Hotspot Subterranean Sites
3. Caveats and Challenges
3.1. Differences in Working Definitions of Troglobionts and Stygobionts
3.2. Differences in Taxonomic Groups, Habitats Sampled, and Sampling Techniques
3.3. Described versus Undescribed Species
- 13 were described as new species;
- 6 were assigned to existing known stygobiotic and troglobiotic species;
- 8 remained unstudied.
4. Protection of Hotspot Caves
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Culver, D.C.; Sket, B. Hotspots of subterranean biodiversity in caves and wells. J. Cave Karst Stud. 2000, 62, 11–17. [Google Scholar]
- Gibert, J.; Deharveng, L. Subterranean ecosystems: A truncated functional diversity. Bioscience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Bregović, P.; Zagmajster, M. Understanding hotspots within a global hotspot—Identifying the drivers of regional species richness patterns in terrestrial subterranean habitats. Insect Conserv. Biodivers. 2016, 9, 268–281. [Google Scholar] [CrossRef]
- Zagmajster, M.; Eme, D.; Fišer, C.; Galassi, D.; Marmonier, P.; Stoch, F.; Cornu, J.; Malard, F. Geographic variation in range size and beta diversity of groundwater crustaceans: Insights from habitats with low thermal seasonality. Glob. Ecol. Biogeogr. 2014, 23, 1135–1145. [Google Scholar] [CrossRef]
- Malard, F.; Boutin, C.; Camacho, A.I.; Ferreira, D.; Michel, G.; Sket, B.; Stoch, F. Diversity patterns of stygobiotic crustaceans across multiple spatial scales in western Europe. Freshw. Biol. 2009, 54, 756–776. [Google Scholar] [CrossRef]
- Culver, D.C.; Deharveng, L.; Bedos, A.; Lewis, J.J.; Madden, M.; Reddell, J.R.; Sket, B.; Trontelj, P.; White, D. The mid-latitude biodiversity ridge in terrestrial cave fauna. Ecography 2006, 29, 120–128. [Google Scholar] [CrossRef]
- Christman, M.C.; Doctor, D.H.; Niemiller, M.L.; Weary, D.J.; Young, J.A.; Zigler, K.S.; Culver, D.C. Predicting the occurrence of cave-inhabiting fauna based on features of the Earth surface environment. PLoS ONE 2016, 11, e0160408. [Google Scholar] [CrossRef]
- Dole-Olivier, M.J.; Castellarini, F.; Coineau, N.; Galassi, D.M.P.; Martin, P.; Mori, N. Towards an optimal sampling strategy to assess groundwater biodiversity comparison across six European regions. Freshw. Biol. 2009, 54, 777–796. [Google Scholar] [CrossRef]
- Humphreys, W.H. Biodiversity patterns in Australia. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: New York, NY, USA, 2019; pp. 109–126. [Google Scholar]
- Guzick, M.T.; Austin, A.D.; Cooper, S.J.B.; Harvey, M.S.; Humphreys, W.F.; Bradford, T.; Eberhard, S.M.; King, R.A.; Leijs, R.; Muirhead, K.A.; et al. Is the Australian subterranean fauna uniquely diverse? Invert. Syst. 2010, 24, 407–418. [Google Scholar] [CrossRef]
- Souza-Silva, M.; Ferreira, R.L. The first two hotspots of subterranean biodiversity in South America. Subterr. Biol. 2016, 19, 1–21. [Google Scholar] [CrossRef]
- Trajano, E.; Gallão, J.E.; Bichuette, M.E. Spots of high diversity of troglobites in Brazil: The challenge of measuring subterranean diversity. Biodivers. Conserv. 2016, 25, 1805–1828. [Google Scholar] [CrossRef]
- Latella, L. Biodiversity: China. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 127–135. [Google Scholar]
- Deharveng, L.; Bedos, A. Biodiversity in the tropics. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 146–162. [Google Scholar]
- Palacios-Vargas, J.G.; Juberthie, C.; Reddell, J.R. México. Mundos Subterráneos 2014–2015, 25–26, 1–101. [Google Scholar]
- Culver, D.C.; Pipan, T. Biology of Caves and Other Subterranean Habitats, 1st ed.; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Culver, D.C.; Pipan, T. Subterranean ecosystems. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Waltham, MA, USA, 2013; Volume 7, pp. 49–62. [Google Scholar]
- Huang, S.; Wei, G.; Wang, H.; Liu, W.; Bedos, A.; Deharveng, L.; Tian, M. Ganxiao Dong: A hotspot of cave biodiversity in northern Guangxi, China. Diversity 2021, 13, 355. [Google Scholar] [CrossRef]
- Zagmajster, M.; Polak, S.; Fišer, C. Postojna Planina Cave System in Slovenia, a hotspot of subterranean biodiversity and a cradle of speleobiology. Diversity 2021, 13, 271. [Google Scholar] [CrossRef]
- Lučić, I.; Sket, B. Vjetrenica. Pogled U Dušu Zemlje; Savez Speleologa Bosne I Hercegovine and Hrvatsko Biospeleološko Društvo: Zagreb, Croatia, 2003. [Google Scholar]
- Ozimec, R.; Lučić, I. The Vjetrenica cave (Bosnia & Herzegovina)–one of the world’s most prominent biodiversity hotspots for cave-dwelling fauna. Subt. Biol. 2009, 7, 17–24. [Google Scholar]
- Polak, S.; Pipan, T. The subterranean fauna of Križna jama, Slovenia. Diversity 2021, 13, 210. [Google Scholar] [CrossRef]
- Camacho, A.I.; Puch, C. Ojo Guareña, a hotspot of subterranean biodiversity in Spain. Diversity 2021, 13, 199. [Google Scholar] [CrossRef]
- Niemiller, M.L.; Helf, K.; Toomey, R.S. Mammoth Cave: A hotspot of subterranean biodiversity in the United States. Diversity 2021, 13, 373. [Google Scholar] [CrossRef]
- Faille, A.; Deharveng, L. The Coume Ouarnède system, a hotspot of subterranean biodiversity in Pyrenees (France). Diversity 2021, 13, 419. [Google Scholar] [CrossRef]
- Brad, T.; Iepure, S.; Sarbu, S.M. The chemoautotrophically based Movile Cave groundwater ecosystem, a hotspot of subterranean biodiversity. Diversity 2021, 13, 128. [Google Scholar] [CrossRef]
- Olivier, M.J.; Martin, D.; Bou, C.; Prié, V. Interprétation du suivi hydrobiologique de la faune stygobie, réalisé sur le système karstique des Cents Fonts lors du pompage d’essai. In Système Karstique des Cent Fonts. Simulation de Scénarios d’Exploitation et de Gestion de la Resource; Ladouche, B., Maréchal, J.C., Dörfliger, N., Lachassagne, P., Eds.; Bureau de Recherches Géologiques et Minières: Montpelier, France, 2006; pp. 127–142. [Google Scholar]
- Iliffe, T.M.; Calderón Gutiérrez. Bermuda’s Walsingham Caves: A global hotspot for anchialine stygobionts. Diversity 2021, 13, 352. [Google Scholar] [CrossRef]
- Hutchins, B.T.; Gibson, J.R.; Diaz, P.H.; Schwartz, B.F. Stygobiont diversity in the San Marcos Artesian Well and Edwards Aquifer groundwater ecosystem, Texas, USA. Diversity 2021, 13, 234. [Google Scholar] [CrossRef]
- Martínez, A.; Gonzalez, B.C. Volcanic anchialine habitats of Lanzarote. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 399–414. [Google Scholar]
- Oromí, P.; Socorro, S. Biodiversity in the Cueva del Vientzo lava tube system (Tenerife, Canary Islands). Diversity 2021, 13, 226. [Google Scholar] [CrossRef]
- Francke, O.F.; Monjaraz-Ruedas, R.; Cruz-López, J. Biodiversity of the Huautla Cave System, Oaxaca, Mexico. Diversity 2021, 13, 429. [Google Scholar] [CrossRef]
- Eberhard, S.M.; Howarth, F.G. Undara basalt flow, a hotspot of subterranean biodiversity, Queensland, Australia. Diversity 2021, 13, 326. [Google Scholar] [CrossRef]
- Clark, H.L.; Buzatto, B.A.; Halse, S.A. A hotspot of arid zone subterranean biodiversity: The Robe Valley in Western Australia. Diversity 2021, 13, 482. [Google Scholar] [CrossRef]
- Deharveng, L.; Rahmadi, C.; Suhardjono, Y.R.; Bedos, A. The Towakkalak System, a hotspot of subterranean biodiversity in Sulawesi, Indonesia. Diversity 2021, 13, 392. [Google Scholar] [CrossRef]
- Jeannel, R. Les Fossiles Vivants des Cavernes; Gallimard: Paris, France, 1944. [Google Scholar]
- Trajano, E.; de Carvalho, M.R. Towards a biologically meaningful classification of subterranean organisms: A critical analysis of the Schiner-Racovitza system from a historical perspective, difficulties of its application and implications for conservation. Subterr. Biol. 2017, 22, 1–26. [Google Scholar] [CrossRef]
- Sket, B. Can we agree on an ecological classification of subterranean animals. J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. Ecological and evolutionary classifications of subterranean organisms. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 376–379. [Google Scholar]
- Christiansen, K.A. Proposition pour la classification des animaux cavernicoles. Spelunca 2019, 2, 75–78. [Google Scholar]
- Deharveng, L.; Bedos, A. Diversity of terrestrial invertebrates in subterranean environments. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 107–172. [Google Scholar]
- Peck, S.B. Eyeless arthropods of the Galapagos Islands, Ecuador: Composition and origin of the cryptozoic fauna of a young, tropical, oceanic archipelago. Biotropica 1990, 22, 366–381. [Google Scholar] [CrossRef]
- Schultz, G.A. Descriptions of new subspecies of Ligidium elrodii (Packard) comb. nov. with notes on other isopod crustaceans from caves in North America (Oniscoidea). Am. Midl. Nat. 1970, 84, 36–45. [Google Scholar] [CrossRef]
- Pipan, T.; Christman, M.C.; Culver, D.C. Dynamics of epikarst communities: Microgeographic pattern and environmental determinants of epikarst copepods in Organ Cave, West Virginia. Am. Midl. Nat. 2006, 156, 75–87. [Google Scholar] [CrossRef]
- Pipan, T. Epikarst–A Promising Habitat; Založba ZRC: Ljubljana, Slovenia, 2005. [Google Scholar]
- Pipan, T.; Culver, D.C.; Papi, F.; Kozel, P. Partitioning diversity in subterranean invertebrates: The epikarst fauna of Slovenia. PLoS ONE 2018, 13, e0185991. [Google Scholar] [CrossRef]
- Schneider, C.; Deharveng, L. First record of the genus Spinaethorax Papáč and Palacios-Vargas, 2016 (Collembola, Neelipleona, Neelidae) in Asia, with a new species from a Vietnamese cave. Eur. J. Taxon. 2017, 363, 1–20. [Google Scholar] [CrossRef][Green Version]
- Ficetola, C.F.; Canadoli, C.; Stoch, F. The Racovitzan impediment and the hidden biodiversity of unexplored environments. Conserv. Biol. 2019, 33, 214–216. [Google Scholar] [CrossRef]
- Holsinger, J.R.; Baroody, R.A.; Culver, D.C. The Invertebrate Cave Fauna of West Virginia; West Virginia Speleological Survey: Barrackville, WV, USA, 1976; No. 7. [Google Scholar]
- Fong, D.W.; Culver, D.C.; Hobbs, H.H., III; Pipan, T. The Invertebrate Cave Fauna of West Virginia, 2nd ed.; West Virginia Speleological Survey: Barrackville, WV, USA, 2007; No. 16. [Google Scholar]
- Mammola, S.; Cardoso, P.; Culver, D.C.; Deharveng, L.; Ferreira, R.L.; Fišer, C.; Galassi, D.M.P.; Griebler, C.; Halse, S.; Humphreys, W.F.; et al. Scientists’ warning on the conservation of subterranean ecosystems. BioScience 2019, 69, 641–650. [Google Scholar] [CrossRef]
- Mammola, S.; Piano, E.; Cardoso, P.; Vernon, P.; Domìnguez-Viller, D.; Culver, D.C.; Pipan, T.; Isaia, M. Climate change going deep: The effects of global climatic alterations on cave ecosystems. Anthr. Rev. 2019, 6, 98–116. [Google Scholar] [CrossRef]
- Niemiller, M.L.; Taylor, S.J.; Bichuette, M.E. Conservation of cave fauna, with emphasis on Europe and the Americas. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 451–478. [Google Scholar]
- Halse, S.J. Conservation and impact assessment of subterranean fauna in Australia. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 479–496. [Google Scholar]
- Olson, R.A. Environmental issues relevant to the Mammoth Cave area. In Mammoth Cave. A Human and Natural History; Hobbs, H.H., III, Olson, R.A., Winkler, E.G., Culver, D.C., Eds.; Springer Nature: Cham, Switzerland, 2017; pp. 265–275. [Google Scholar]
- Steele, C.W.; Shifflett, T.E. Huautla cave system (Sistema Huautla), Mexico. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: New York, NY, USA, 2019; pp. 527–536. [Google Scholar]



{kind=link}
{kind=link}
| Stygo- bionts | Rank | Troglo-bionts | Rank | Total | Rank | No. Unde-scribed | Terrain and Hydrogeology | Latitude | YearAssessed | Reference | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperate | |||||||||||
| Postojna Planina System (SLO) * | 72 | 1 | 45 | 1 | 117 | 1 | 8 | karst | 45.7 | 2021 | [19] |
| Vjetrenica (B&H) | 42 | 7 | 39 | 2 | 81 | 2 | 3 | karst | 42.9 | 2009 | [20,21] |
| Križna Jama (SLO) * | 32 | 10 | 28 | 6 | 60 | 4 | 5 | karst | 45.7 | 2021 | [22] |
| Ojo Guareña (ESP) * | 46 | 5 | 8 | 54 | 6 | 23 | karst | 43.0 | 2021 | [23] | |
| Logarček (SLO) | 28 | 15 | 43 | 9 | ? | karst | 43.5 | 2000 | [1] | ||
| Mammoth Cave (USA) * | 17 | 32 | 4 | 49 | 7 | 0 | karst | 45.9 | 2021 | [24] | |
| Coume Ouarnède (FRA) * | 21 | 17 | 38 | 3 | karst | 43.0 | 2021 | [25] | |||
| Pestera de la Movile (RO) * | 13 | 25 | 38 | 4 | karst, chemoautotrophic | 37.2 | 2021 | [26] | |||
| Cent fonts (FRA) | 43 | 6 | 0 | 43 | 9 | 7 | karst, phreatic | 43.6 | 2006 | [27] | |
| Triadou aquifer (FRA) | 34 | 9 | 0 | 34 | ? | karst, phreatic | 43.7 | 2000 | [1] | ||
| Baget system (FRA) 1 | 24 | 9 | 33 | ? | karst, phreatic | 43.0 | 2000 | [1] | |||
| Subtemperate | |||||||||||
| Walsingham Cave (BER) * | 65 | 2 | 0 | 65 | 3 | 0 | karst, anchialine | 32.3 | 2021 | [28] | |
| San Marcos Artesian Well (USA) * | 55 | 3 | 0 | 55 | 5 | 16 | karst, phreatic, chemoautotrophic | 29.9 | 2021 | [29] | |
| Jameos del Agua (ESP) | 40 | 8 | 0 | 40 | 4 | lava tube, anchialine | 28.4 | 2018 | [30] | ||
| Cueva de Felipe Reventon (ESP) * | 0 | 38 | 3 | 38 | 0 | lava tube | 28.4 | 2021 | [31] | ||
| Cueva del Viento-Sobrado (ESP) * | 0 | 28 | 6 | 28 | 0 | lava tube | 28.4 | 2021 | [31] | ||
| Sub-tropical | |||||||||||
| Sistema Huautla (MEX) * | 0 | 27 | 9 | 27 | 10 | deep karst | 18.1 | 2021 | [32] | ||
| Undara Basalt Flow (AUS) *2 | 1 | 30 | 5 | 31 | 27 | lava tube | −18.2 | 2021 | [33] | ||
| Robe Valley wells (AUS) *3 | 48 | 4 | 0 | 48 | 8 | 18 | karst, phreatic | −21.6 | 2021 | [34] | |
| Sistema Purificacion (MEX) | 3 | 28 | 6 | 31 | ? | karst | 23.8 | 2019 | [14] | ||
| Areias system (BRA) | 6 | 22 | 28 | 14 | karst | −24.6 | 2016 | [11] | |||
| Tropical | |||||||||||
| Towakkalak (SUL) * | 10 | 26 | 10 | 36 | 17 | karst | −5.0 | 2021 | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Culver, D.C.; Deharveng, L.; Pipan, T.; Bedos, A. An Overview of Subterranean Biodiversity Hotspots. Diversity 2021, 13, 487. https://doi.org/10.3390/d13100487
Culver DC, Deharveng L, Pipan T, Bedos A. An Overview of Subterranean Biodiversity Hotspots. Diversity. 2021; 13(10):487. https://doi.org/10.3390/d13100487
Chicago/Turabian StyleCulver, David C., Louis Deharveng, Tanja Pipan, and Anne Bedos. 2021. "An Overview of Subterranean Biodiversity Hotspots" Diversity 13, no. 10: 487. https://doi.org/10.3390/d13100487
APA StyleCulver, D. C., Deharveng, L., Pipan, T., & Bedos, A. (2021). An Overview of Subterranean Biodiversity Hotspots. Diversity, 13(10), 487. https://doi.org/10.3390/d13100487