
Issi saaneq gen. et sp. nov.—A New Sauropodomorph Dinosaur from the Late Triassic (Norian) of Jameson Land, Central East Greenland †

 ,
,  , , and
, , and
Abstract
:
1. Introduction
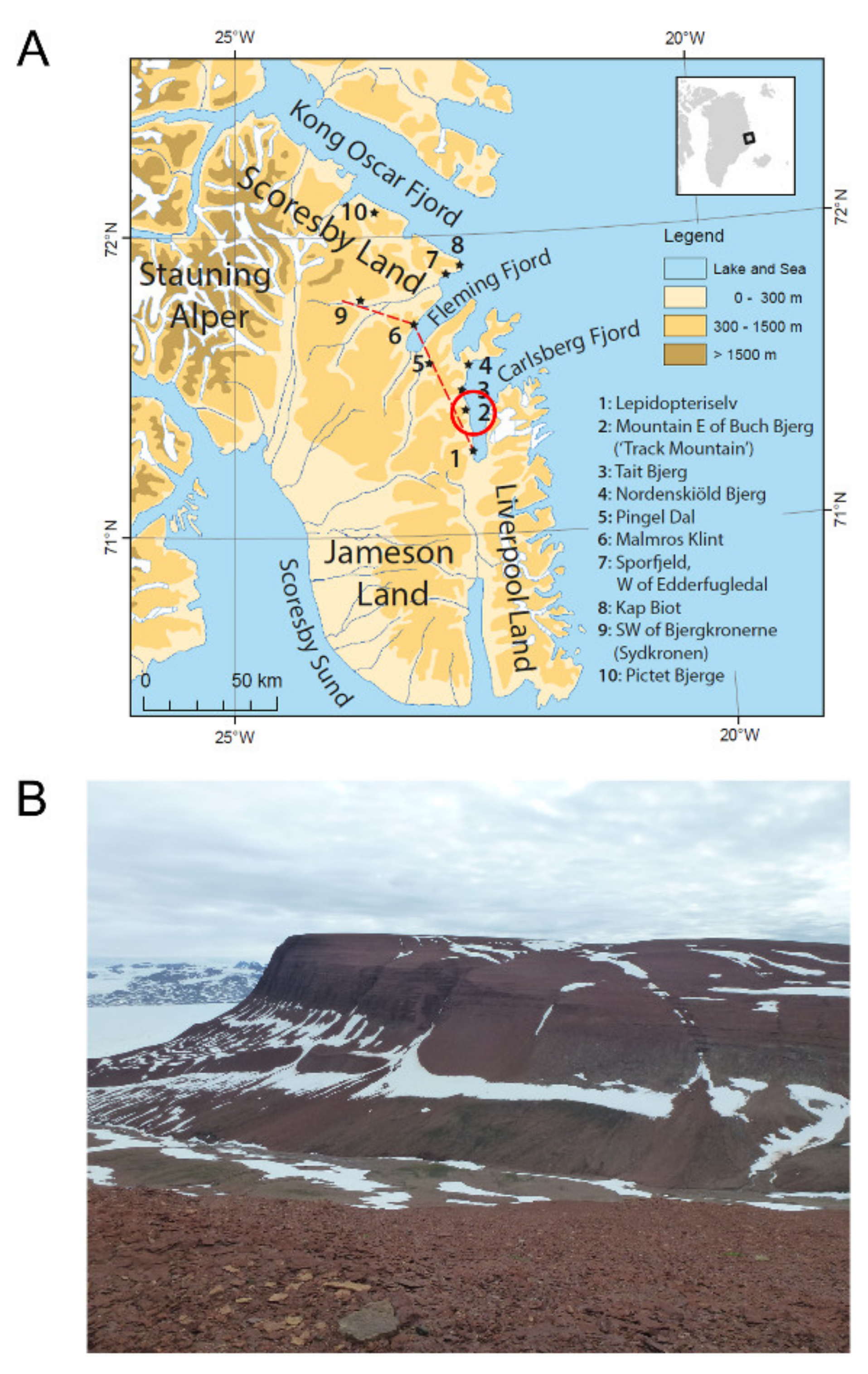
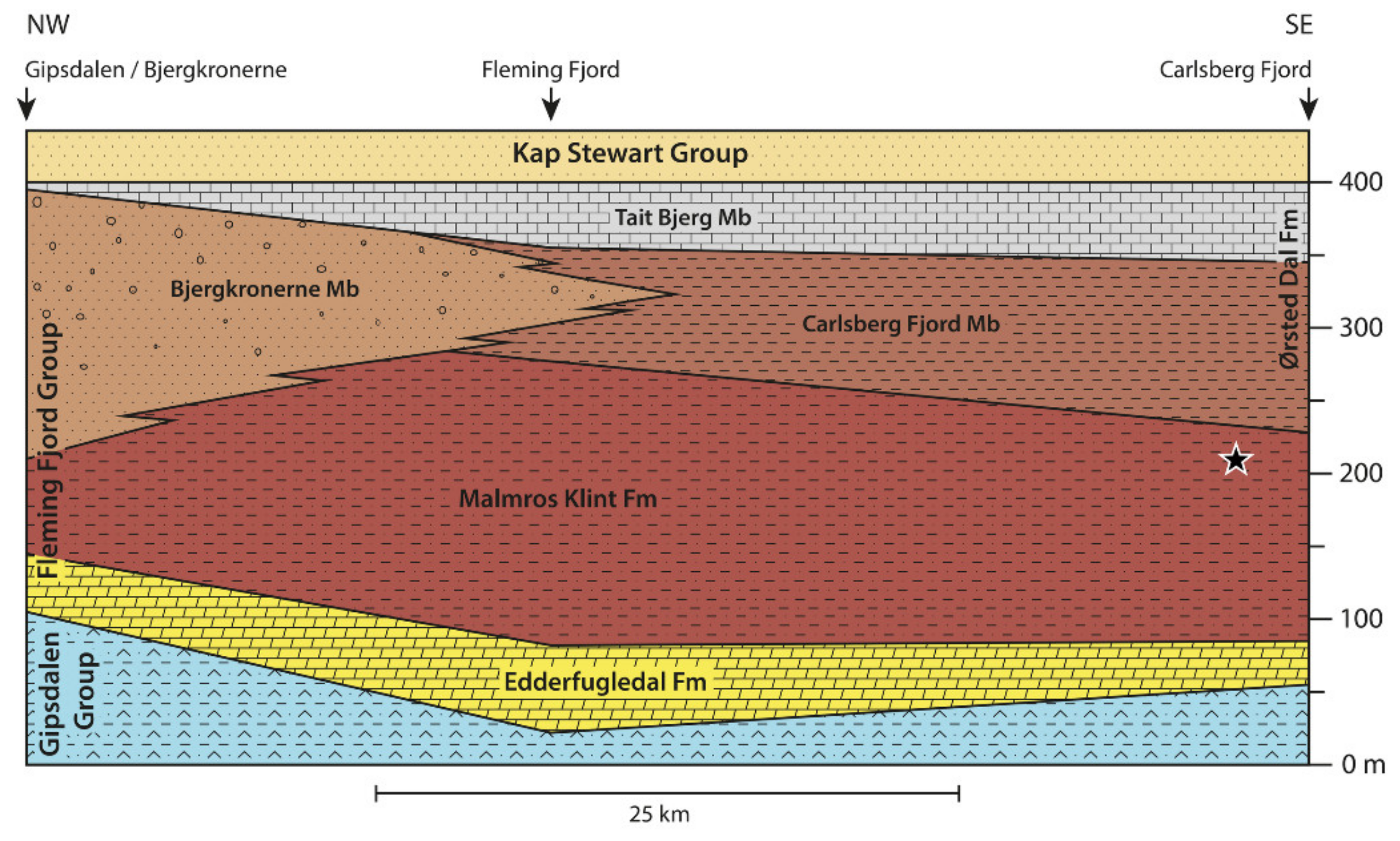
2. Geological Setting
3. Materials and Methods
3.1. Specimens in This Study
3.2. Digitization and Image Treatment
3.3. Skull Reconstruction
3.4. Phylogenetic Analysis
3.5. Institutional Abbreviations
3.6. Nomenclature Acts
4. Results
4.1. Systematic Palaeontology
4.2. Etymology
4.3. Holotype
4.4. Paratype
4.5. Locality
4.6. Horizon and Age
4.7. Diagnosis
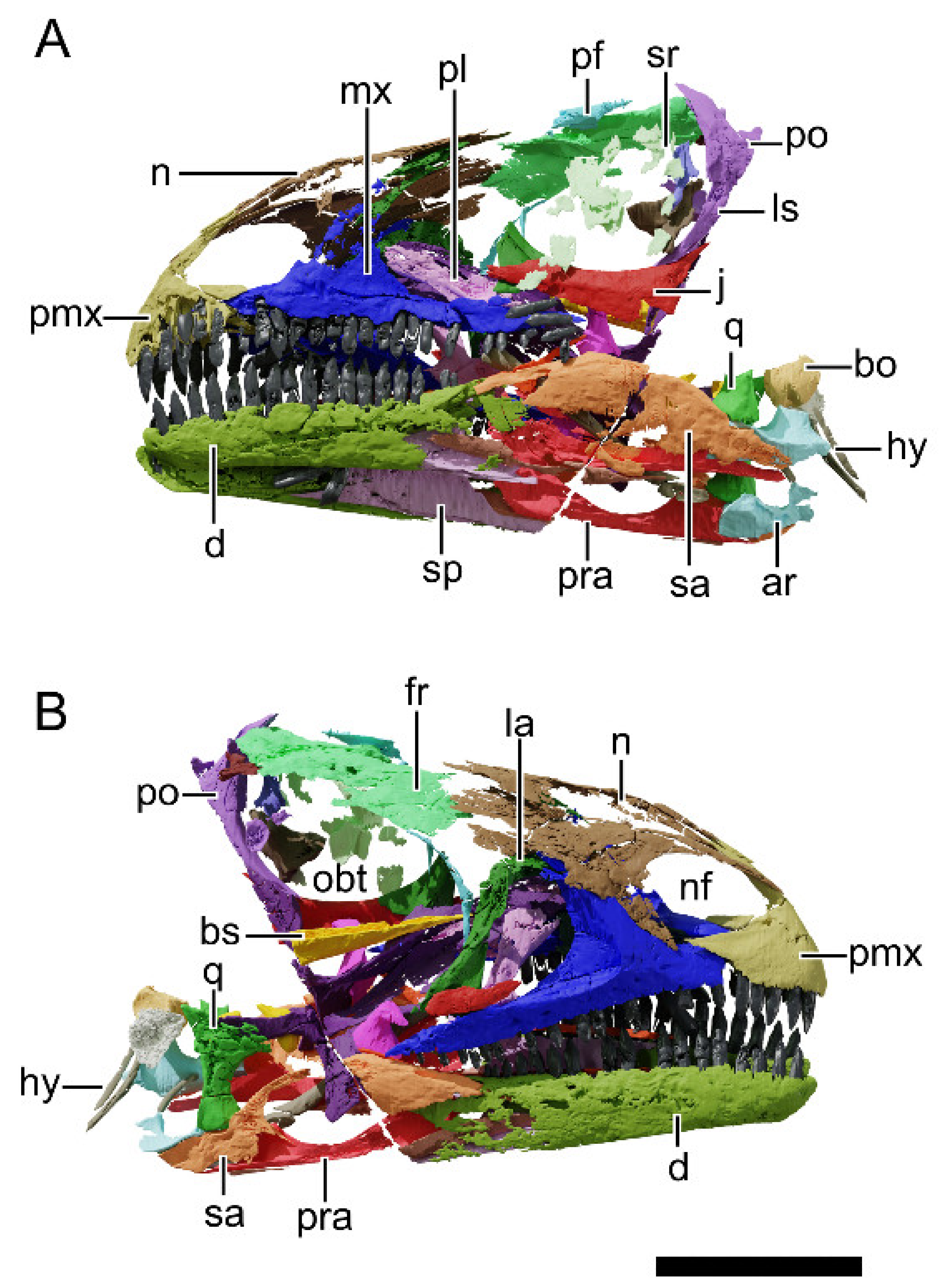
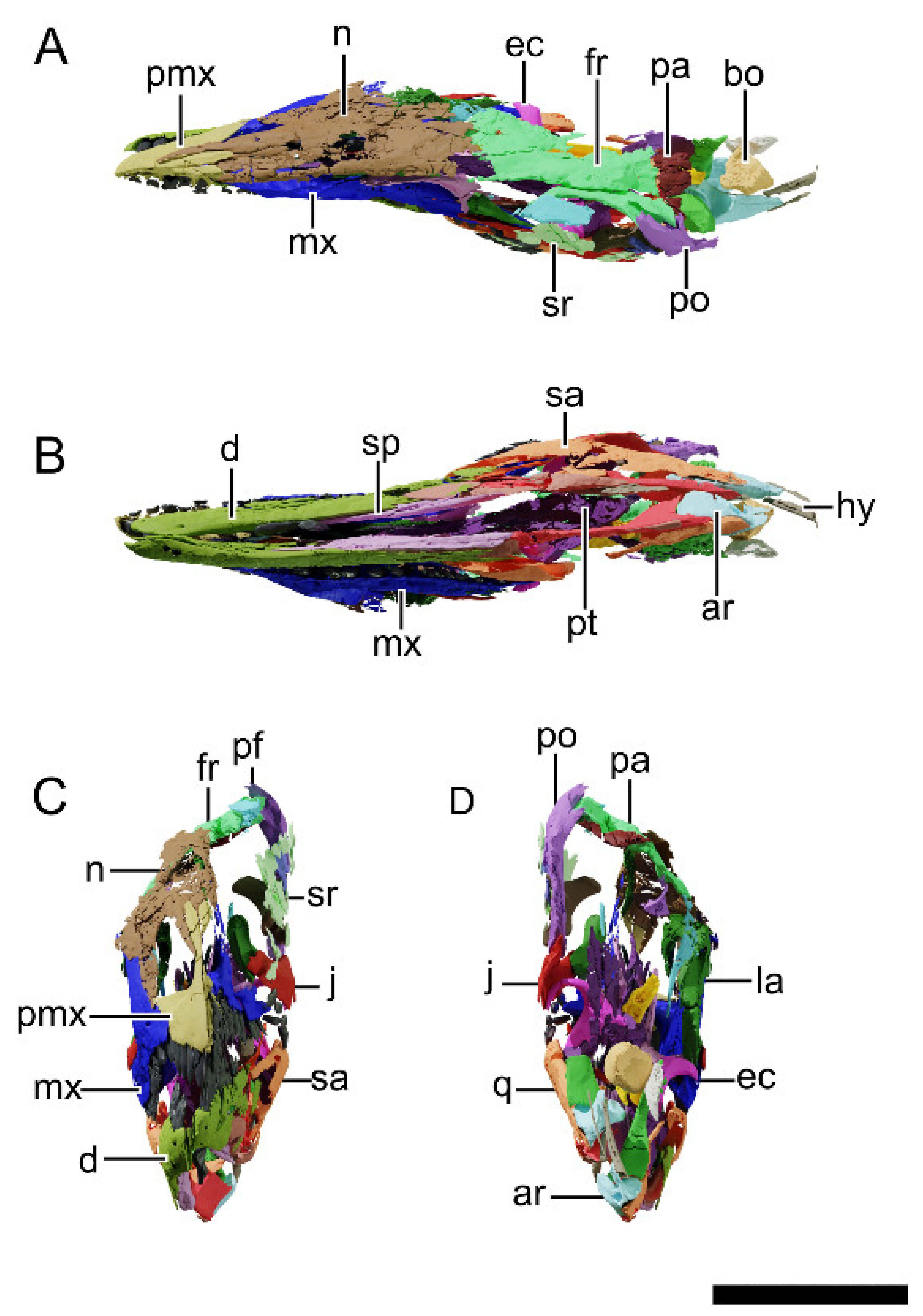
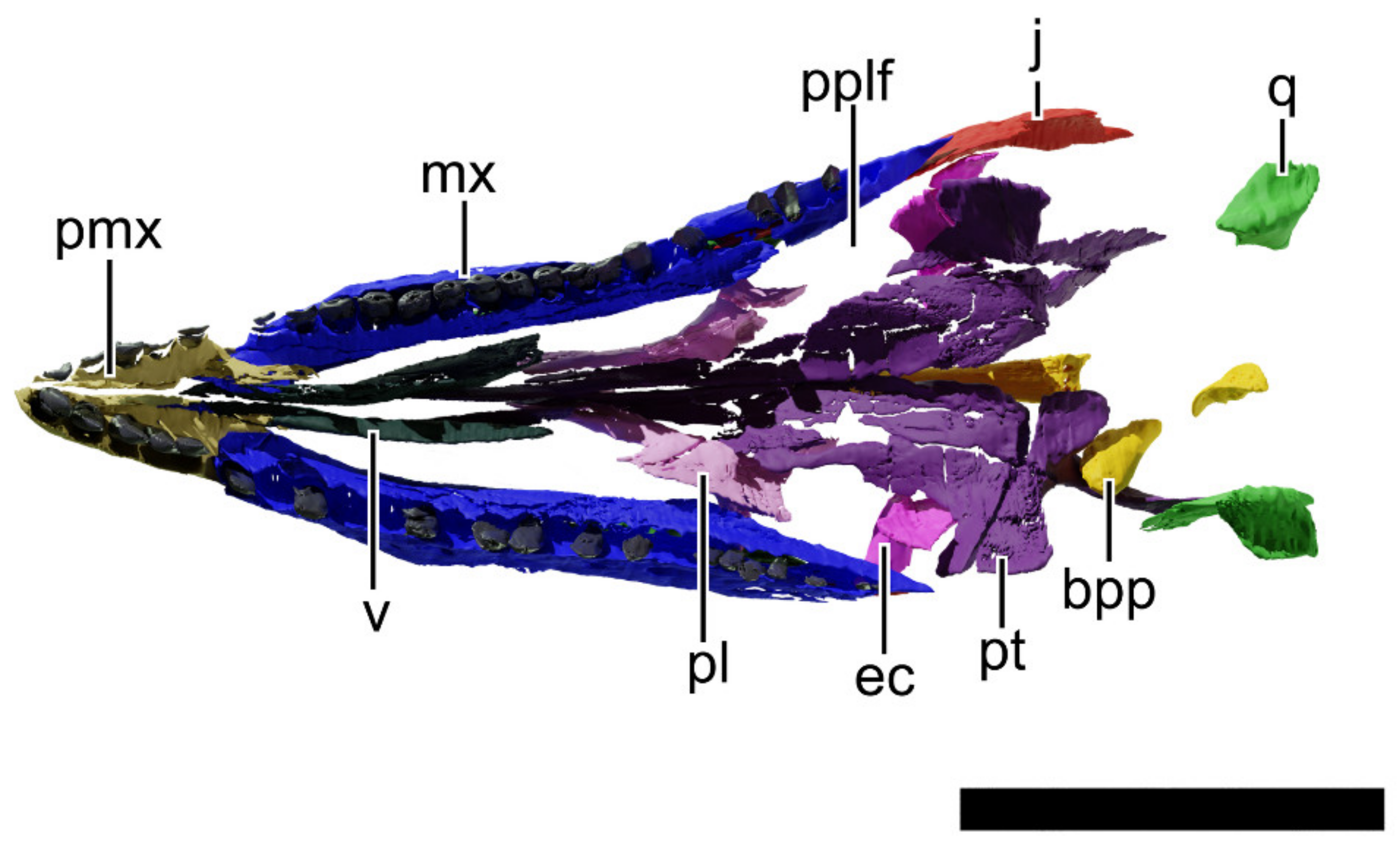
4.8. Description and Comparative Anatomy
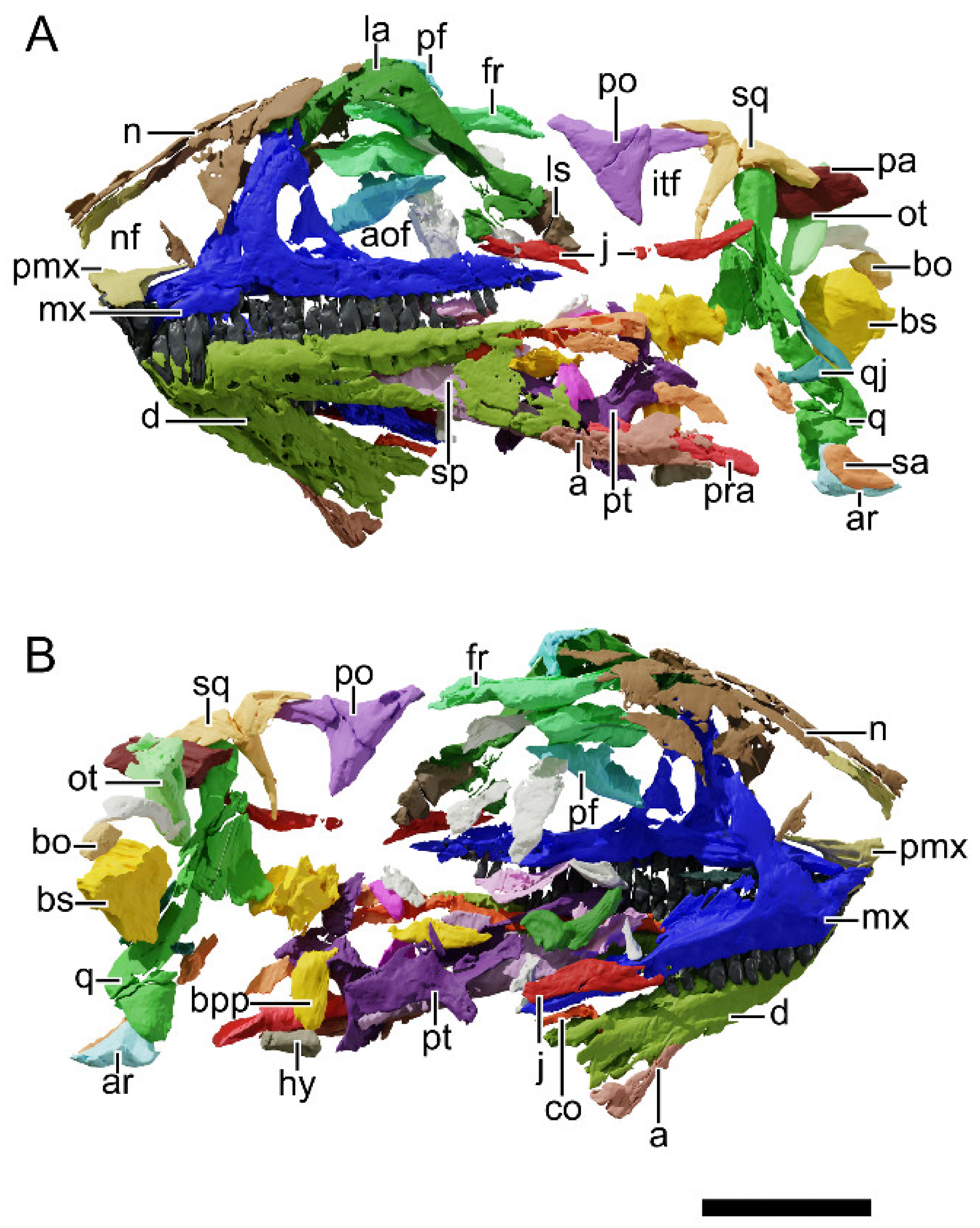
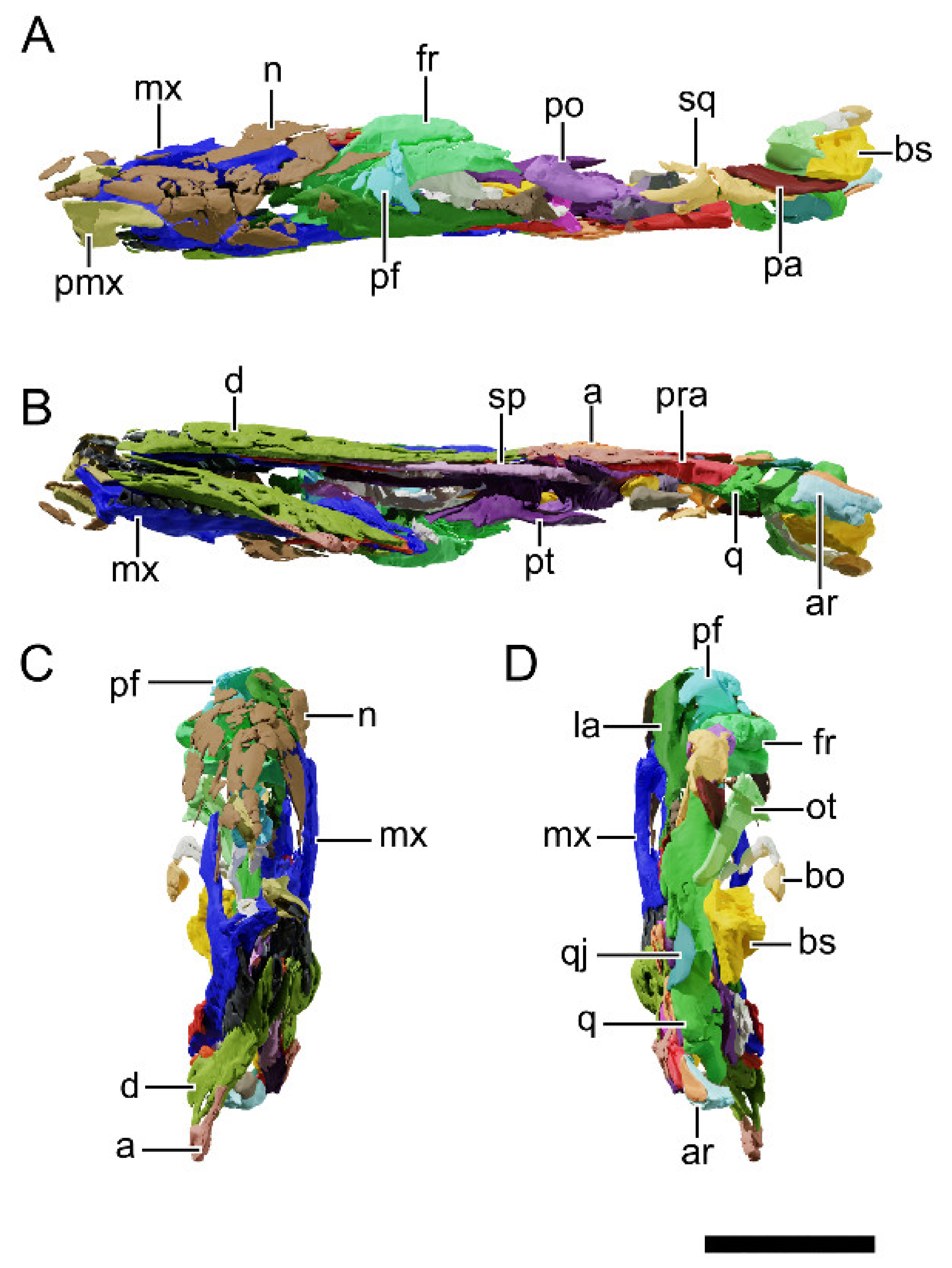
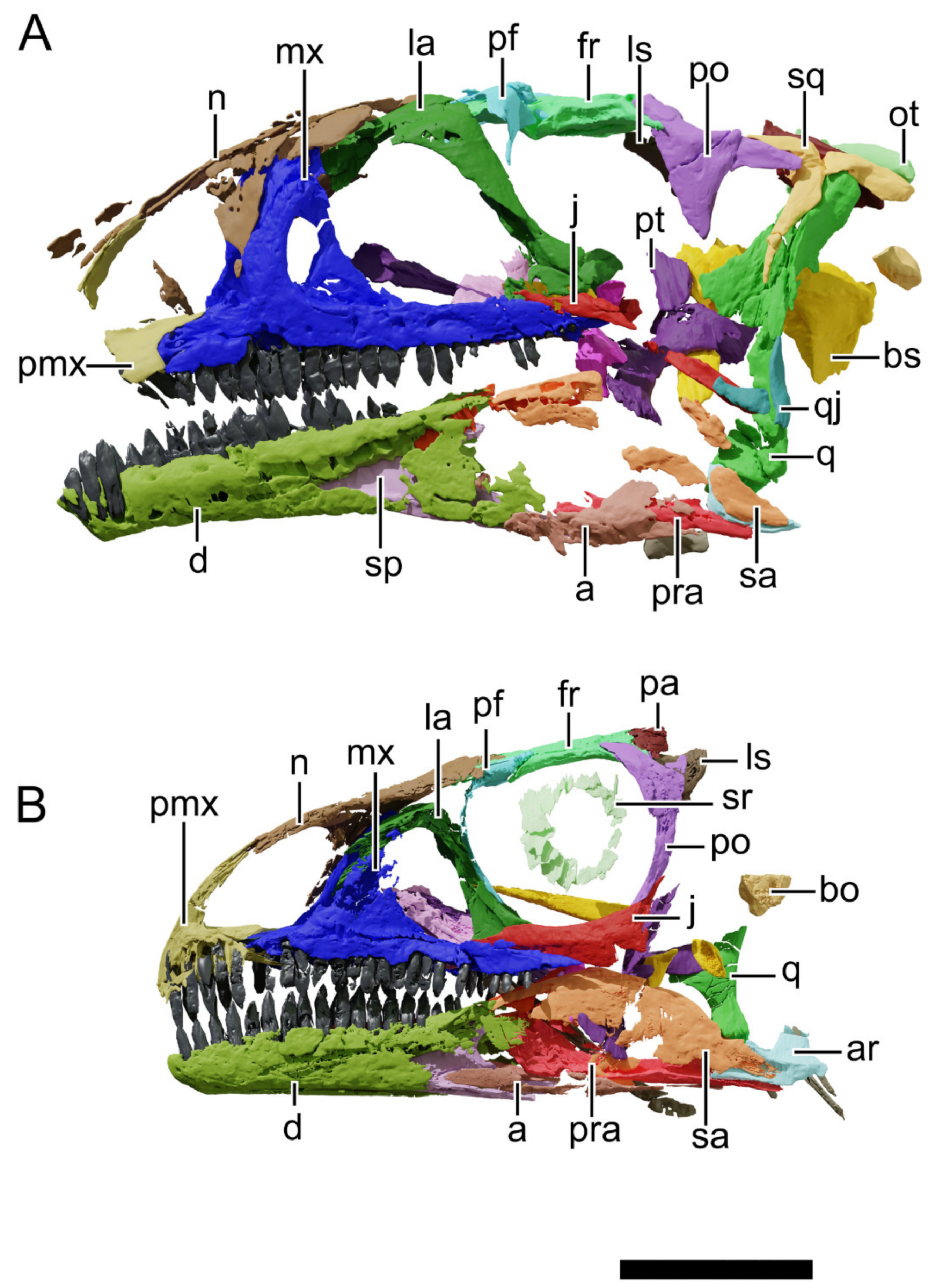
4.8.1. Generalities
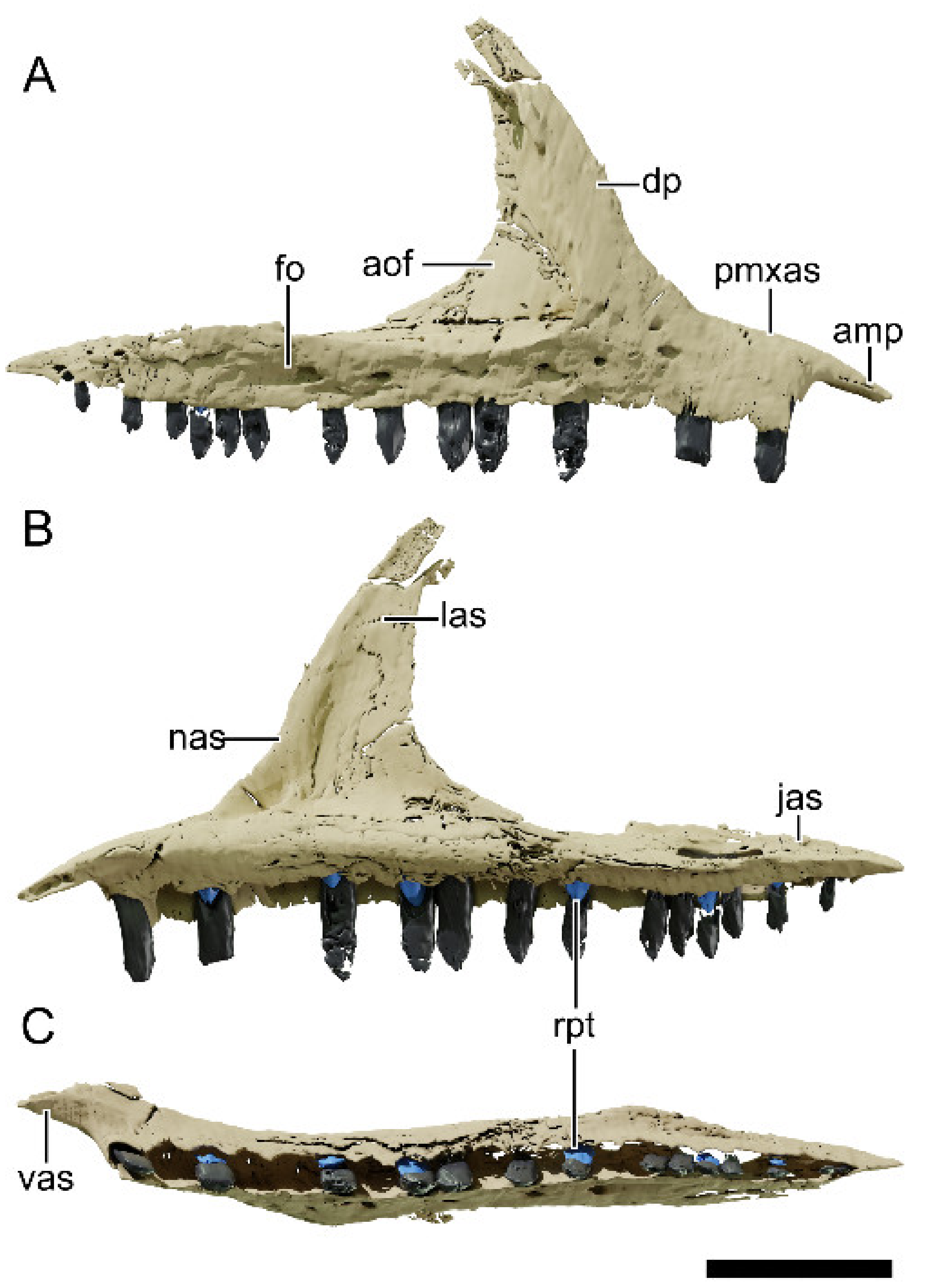
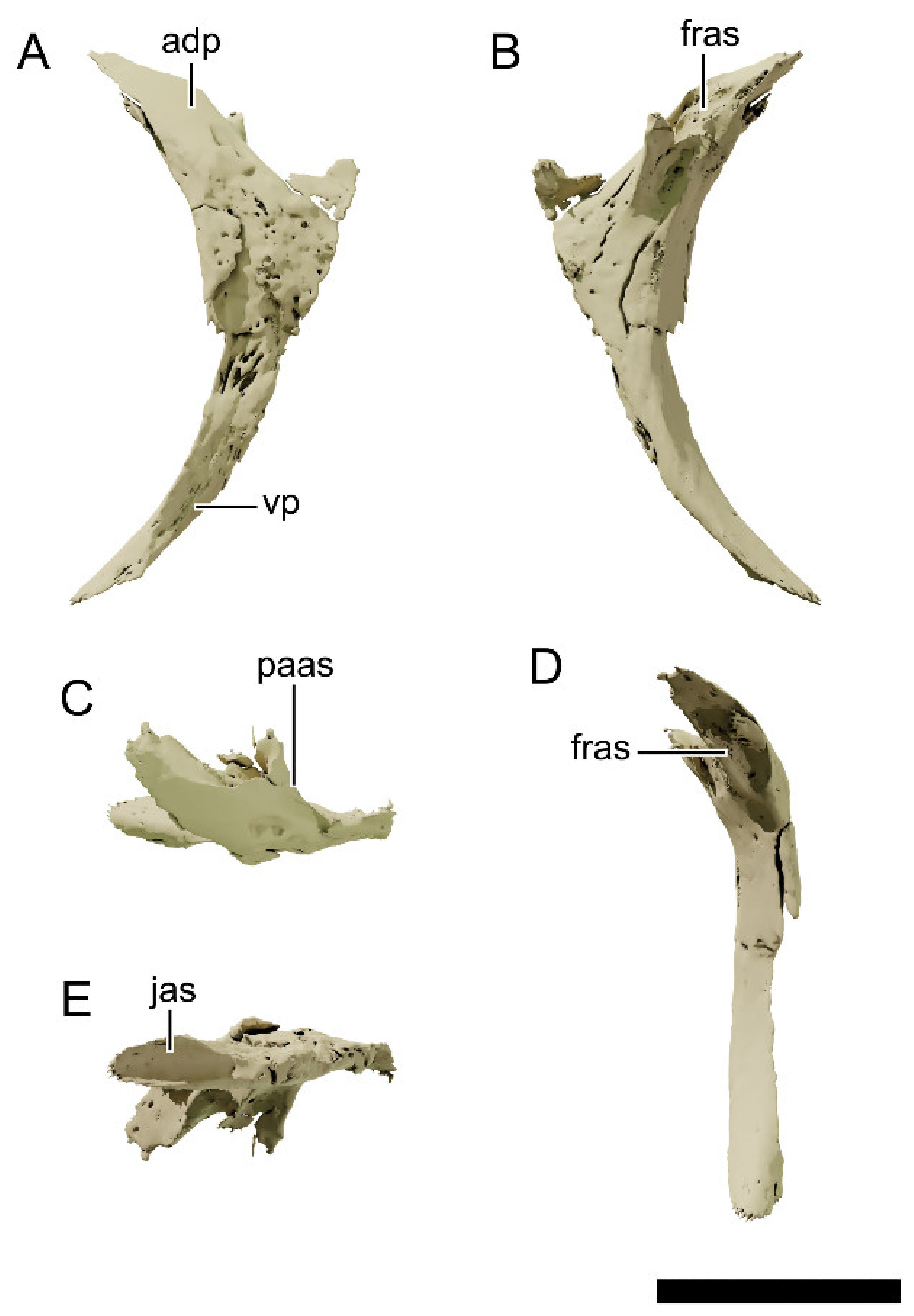
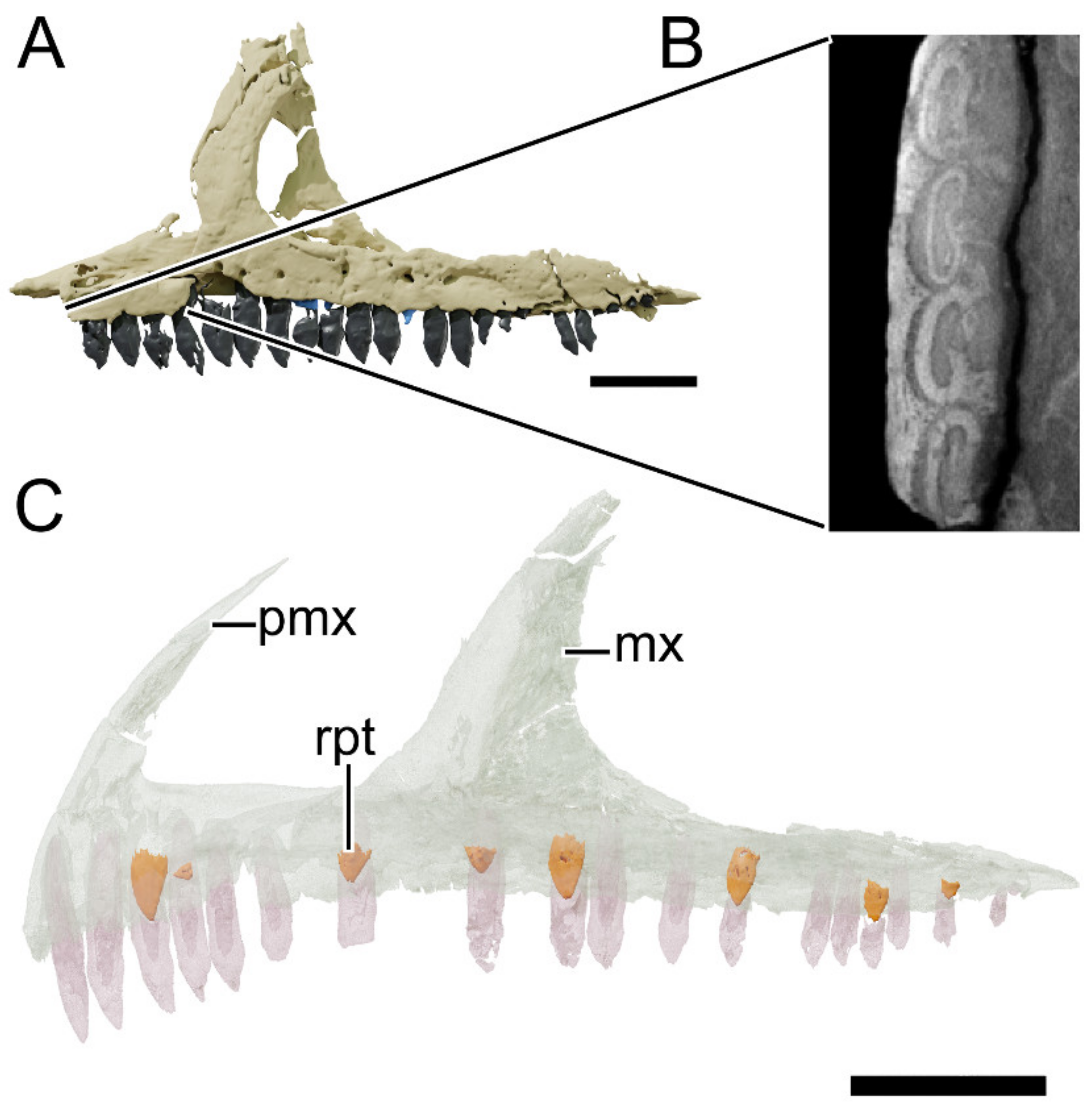
4.8.2. Premaxilla
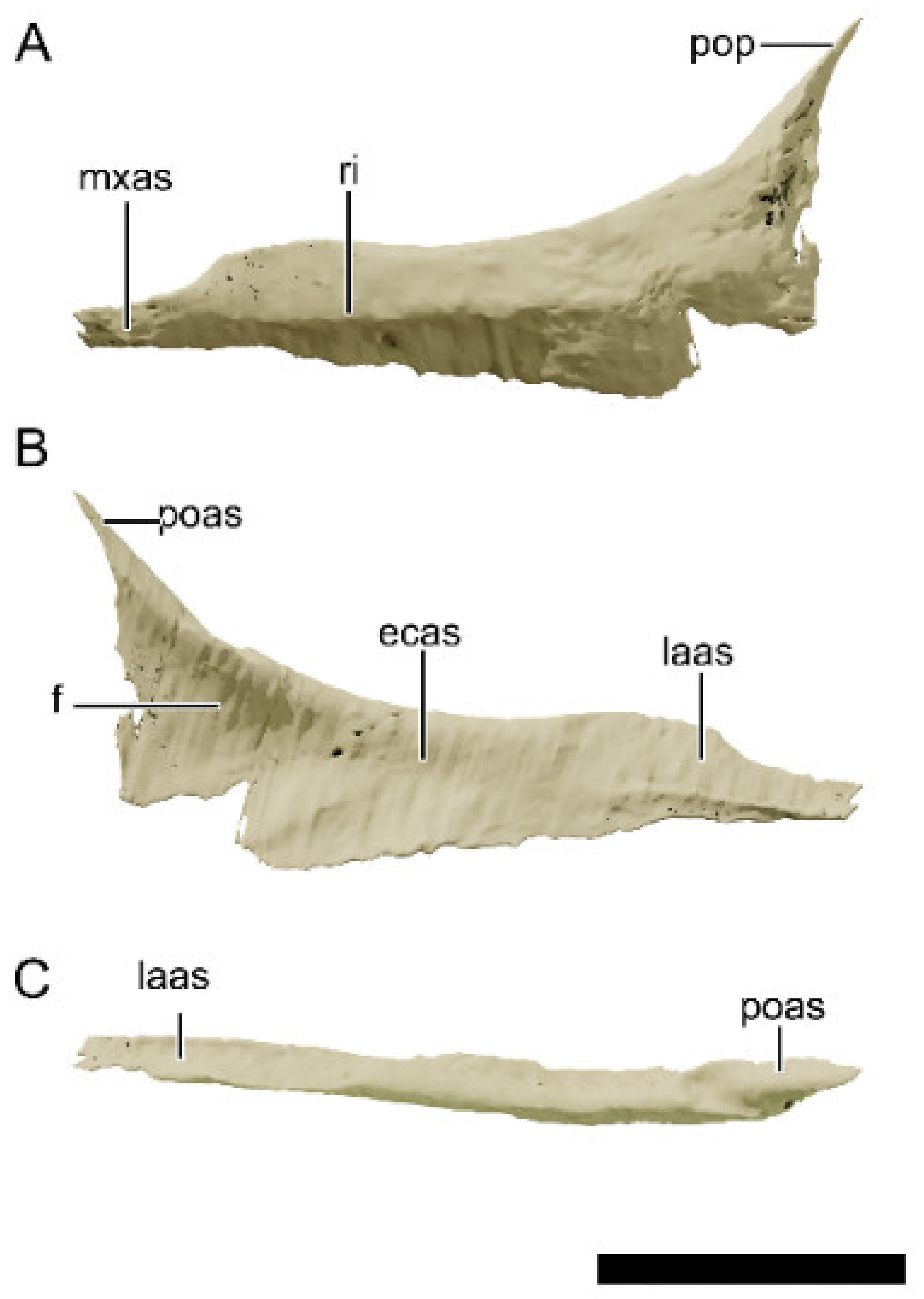
4.8.3. Maxilla
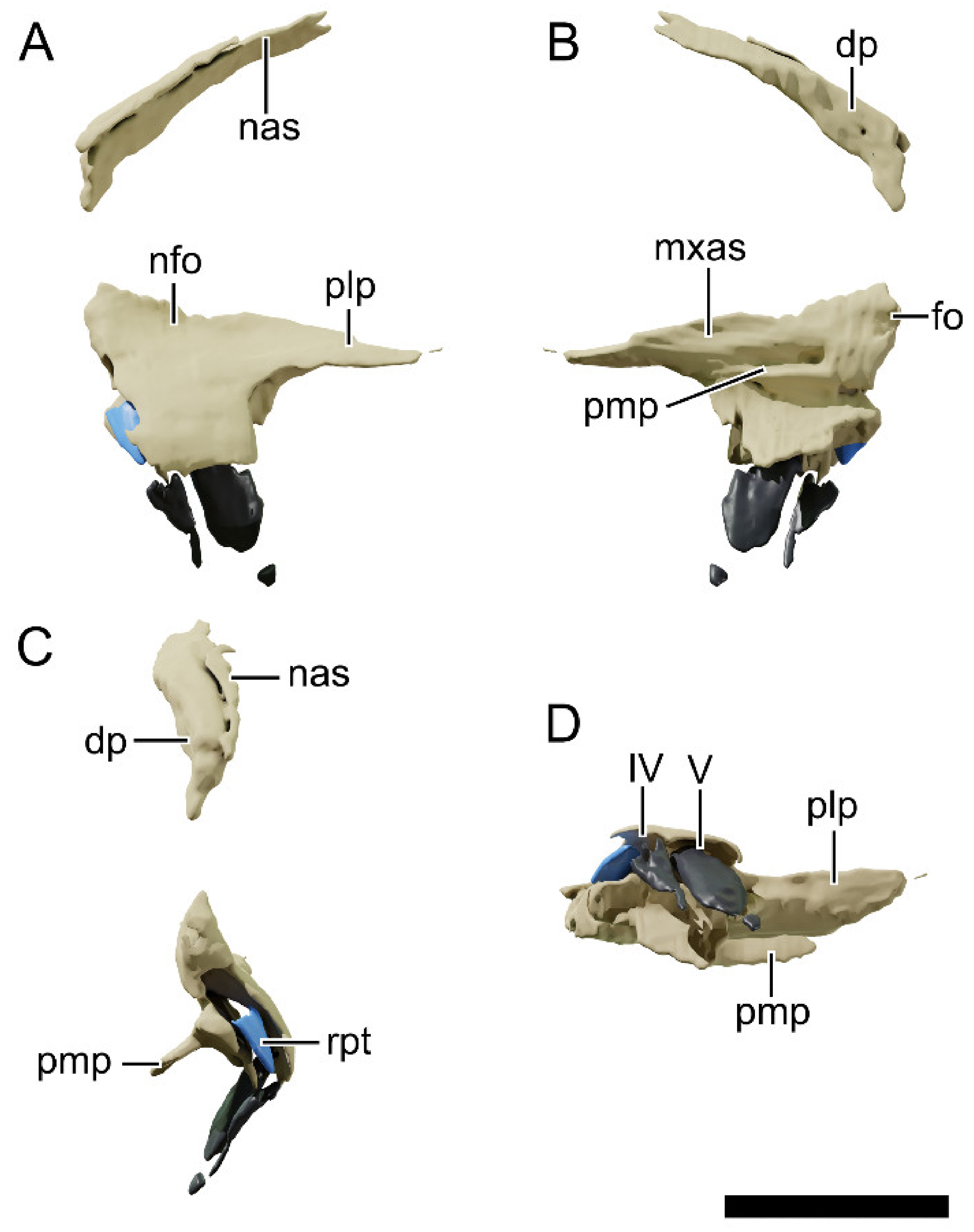
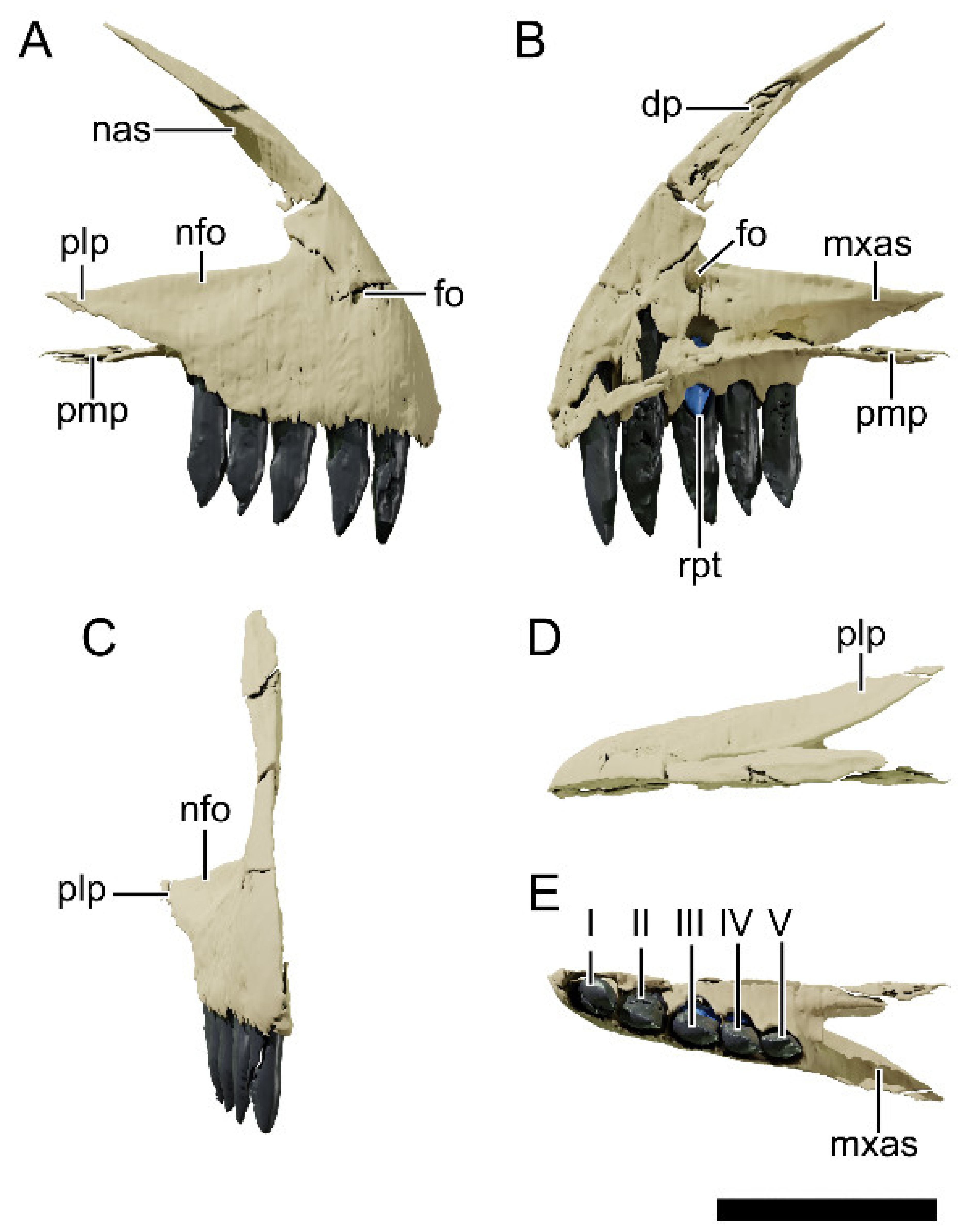
4.8.4. Nasal
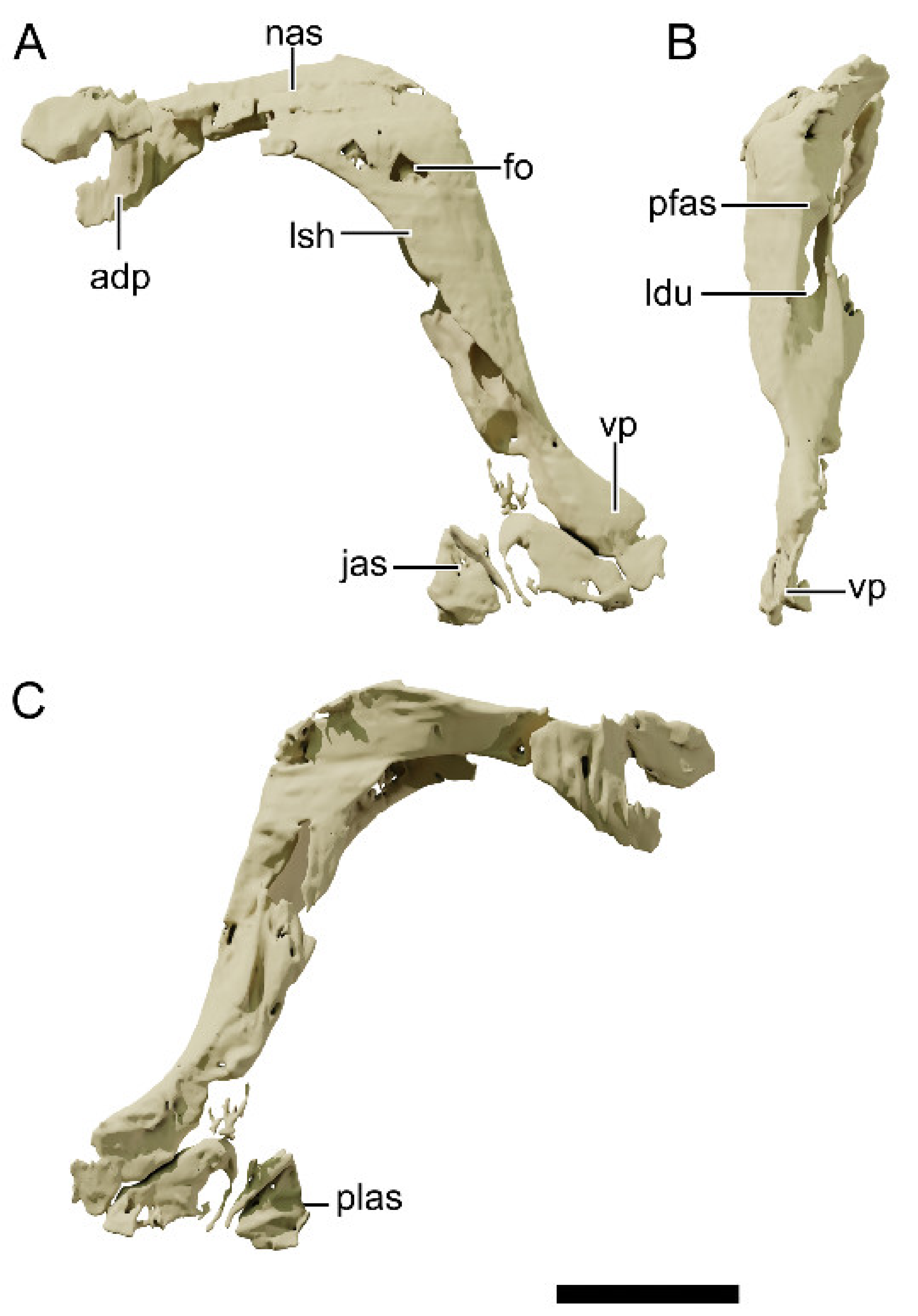
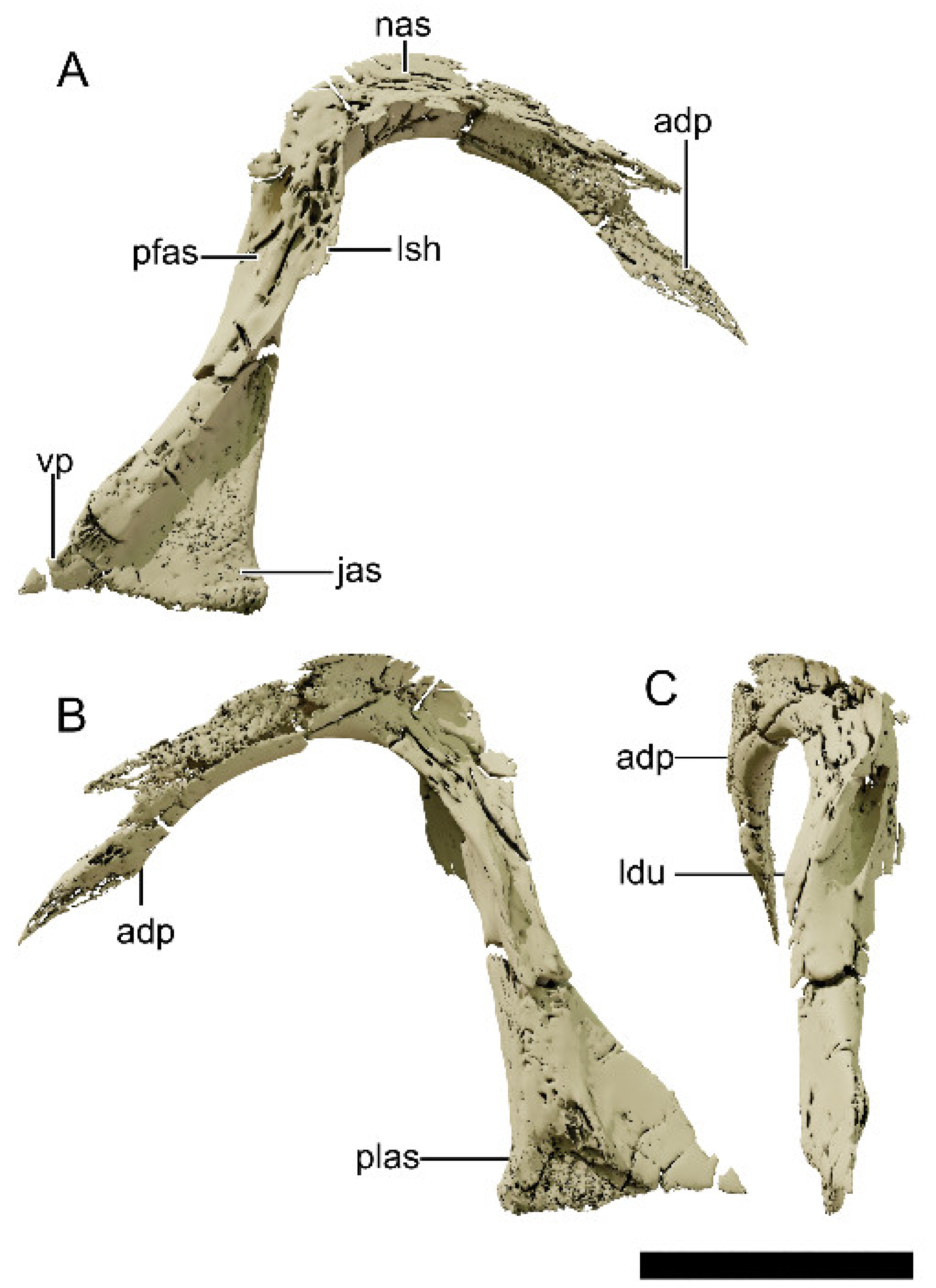
4.8.5. Lacrimal
4.8.6. Prefrontal
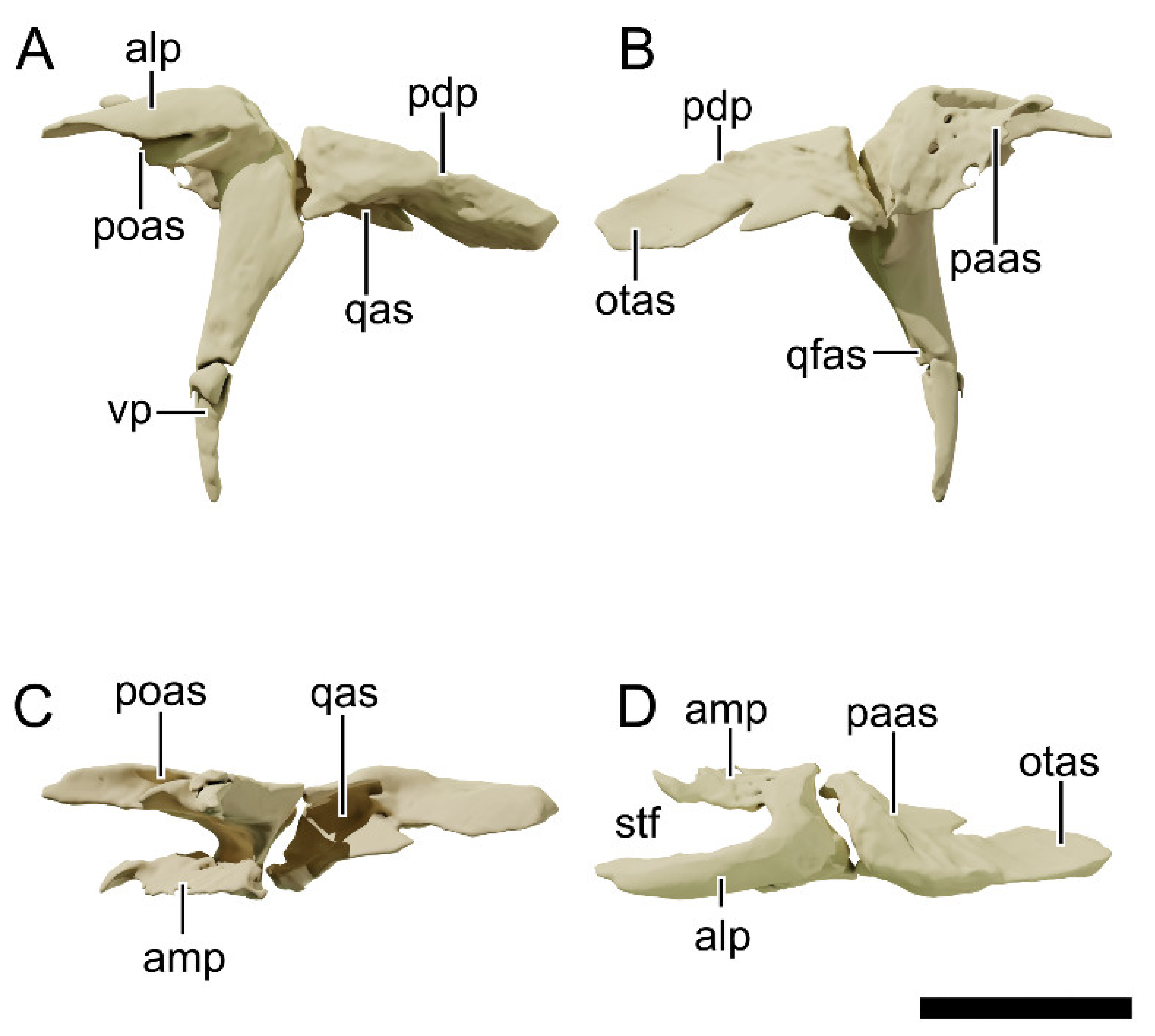
4.8.7. Postorbital
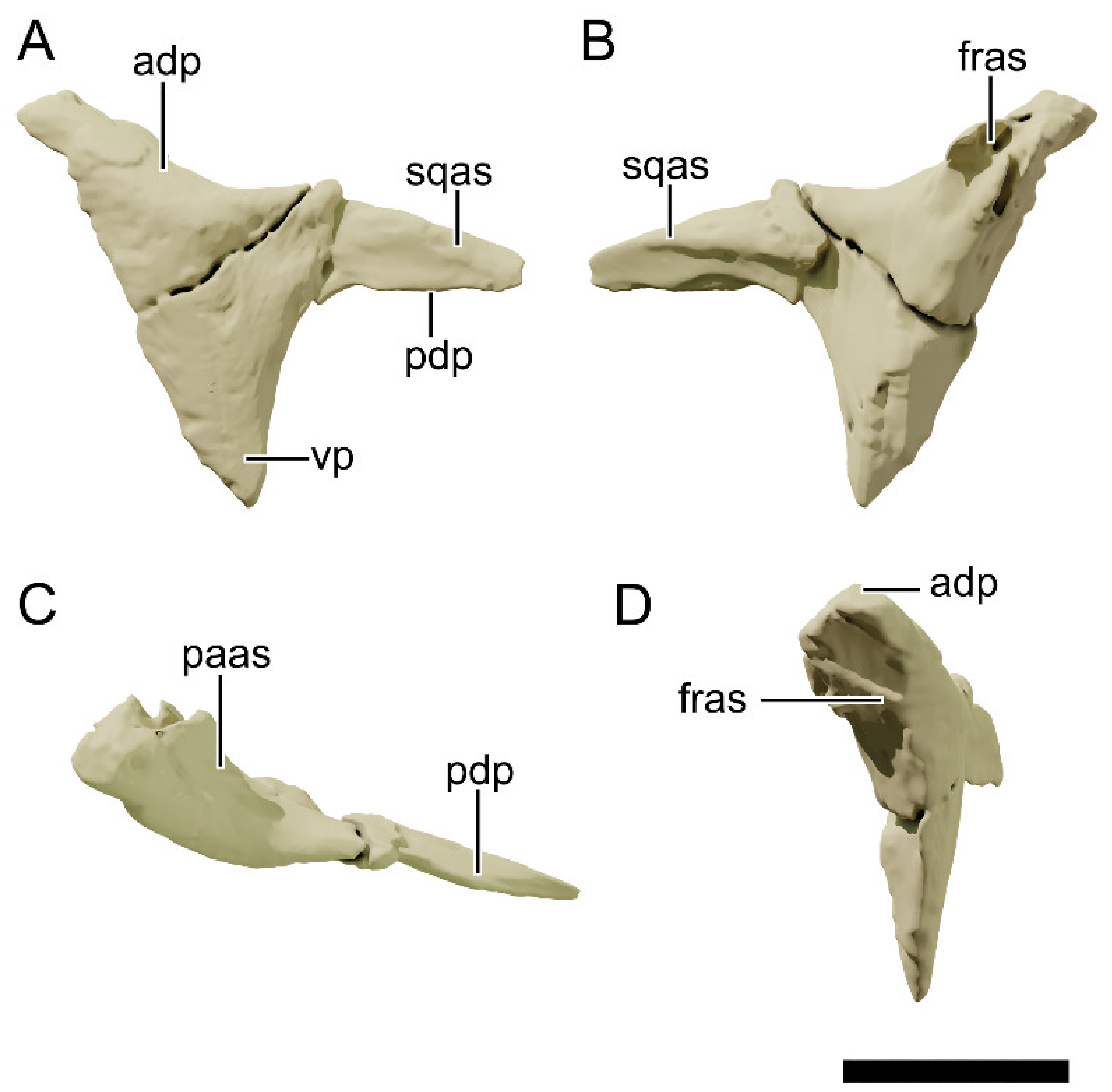
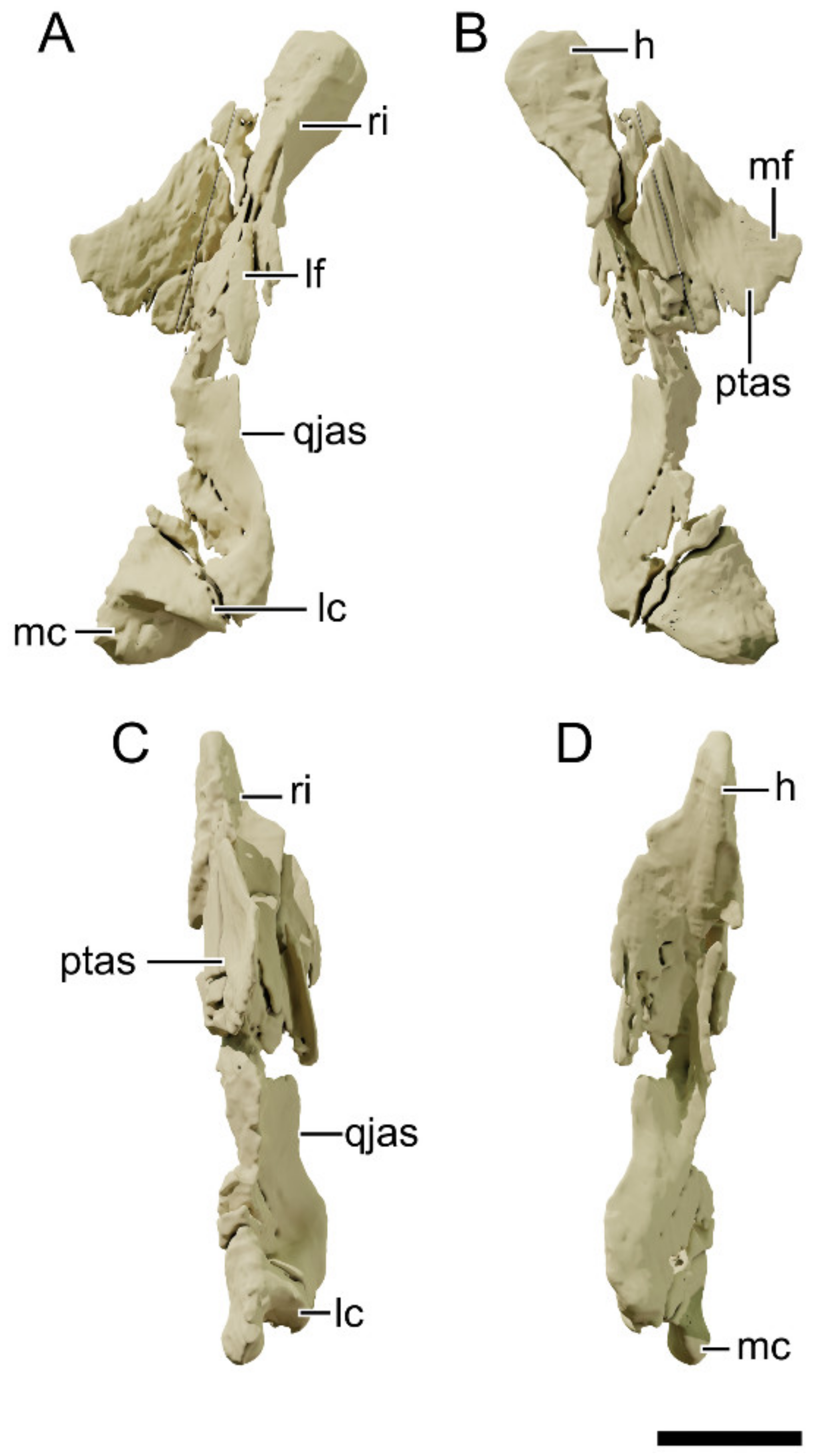
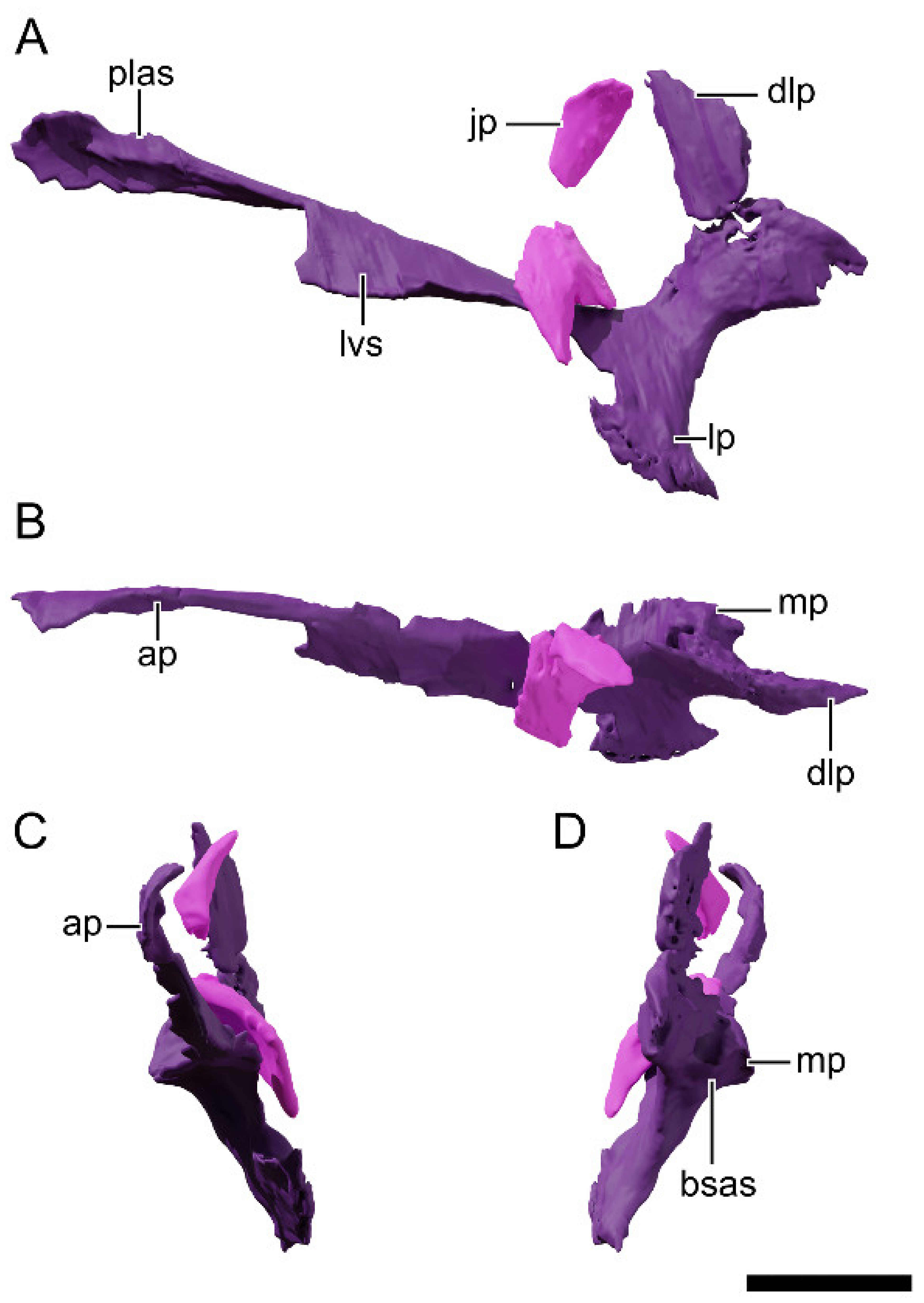
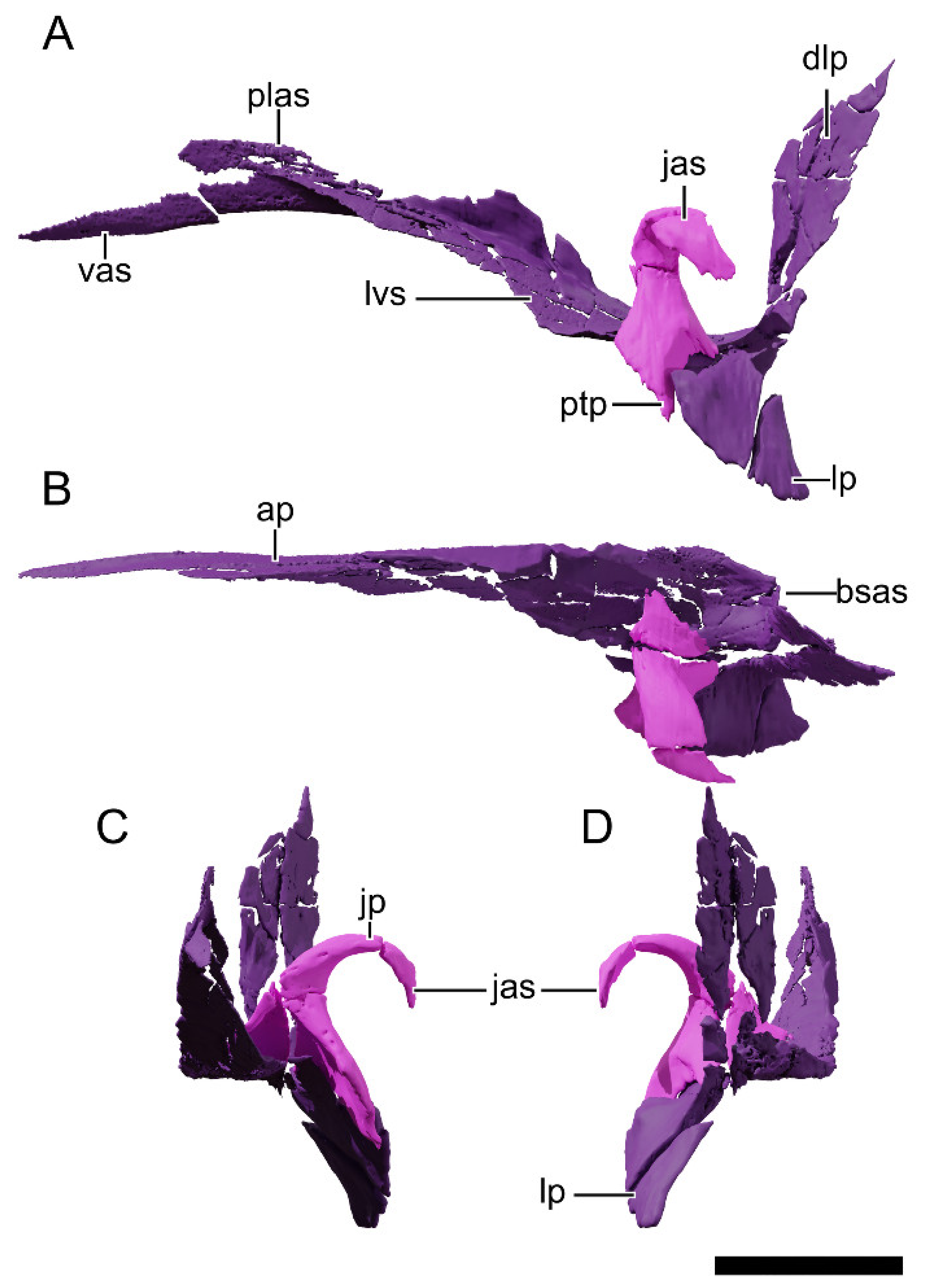
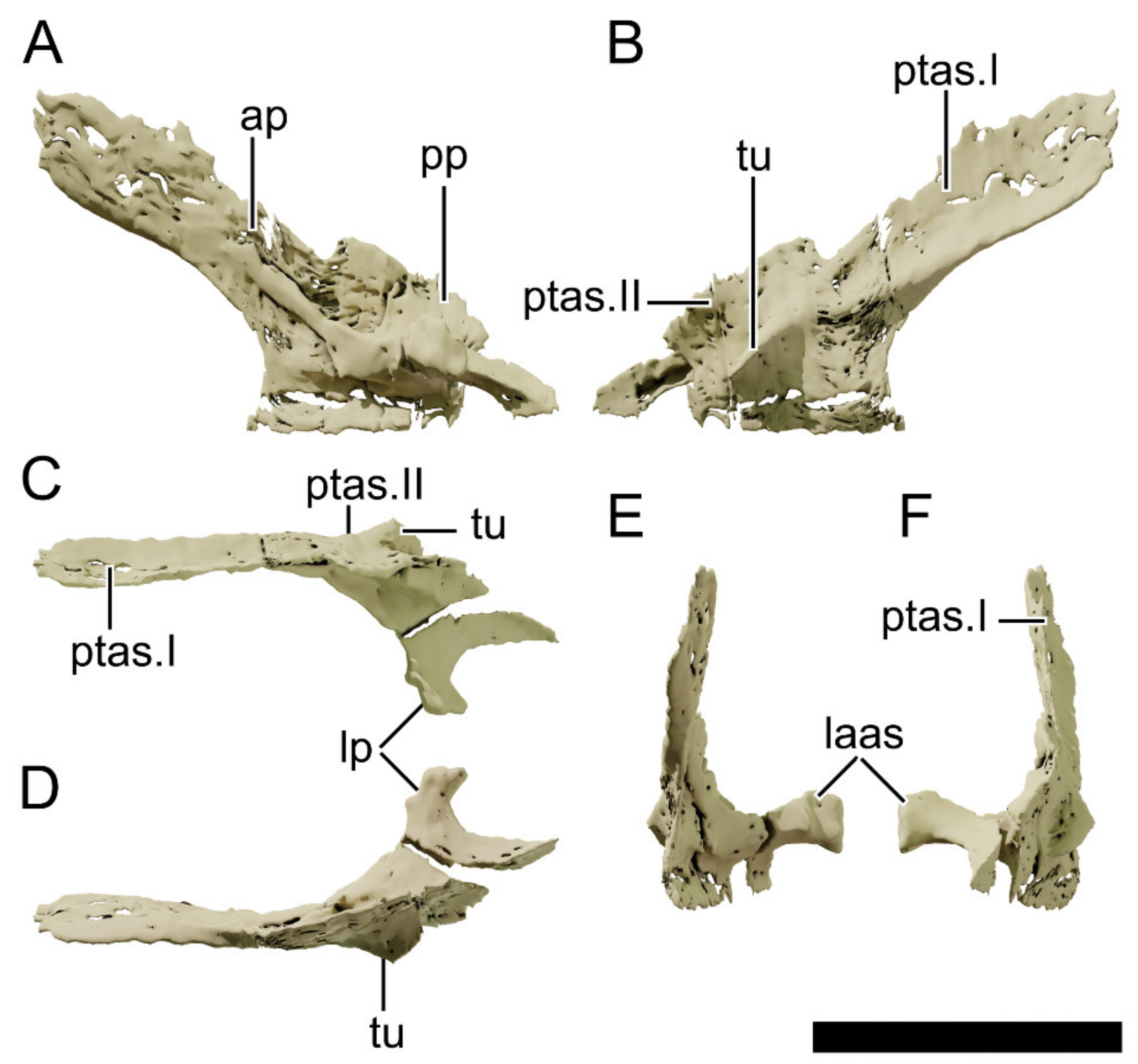
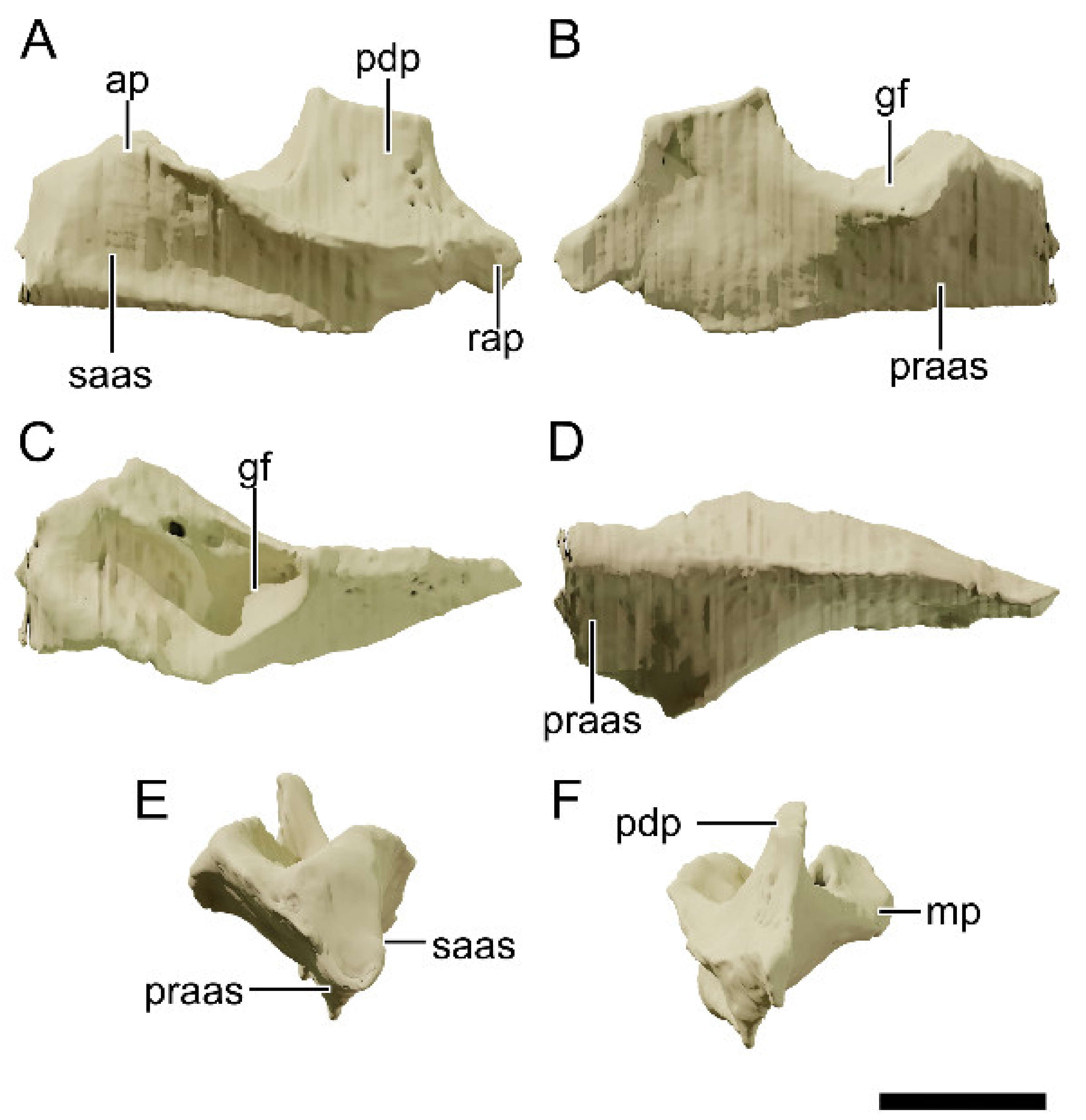
4.8.8. Squamosal
4.8.9. Jugal
4.8.10. Quadratojugal
4.8.11. Quadrate
4.8.12. Frontal
4.8.13. Parietal
4.8.14. Orbitosphenoid
4.8.15. Laterosphenoid
4.8.16. Otoccipital
4.8.17. Basioccipital
4.8.18. Basisphenoid
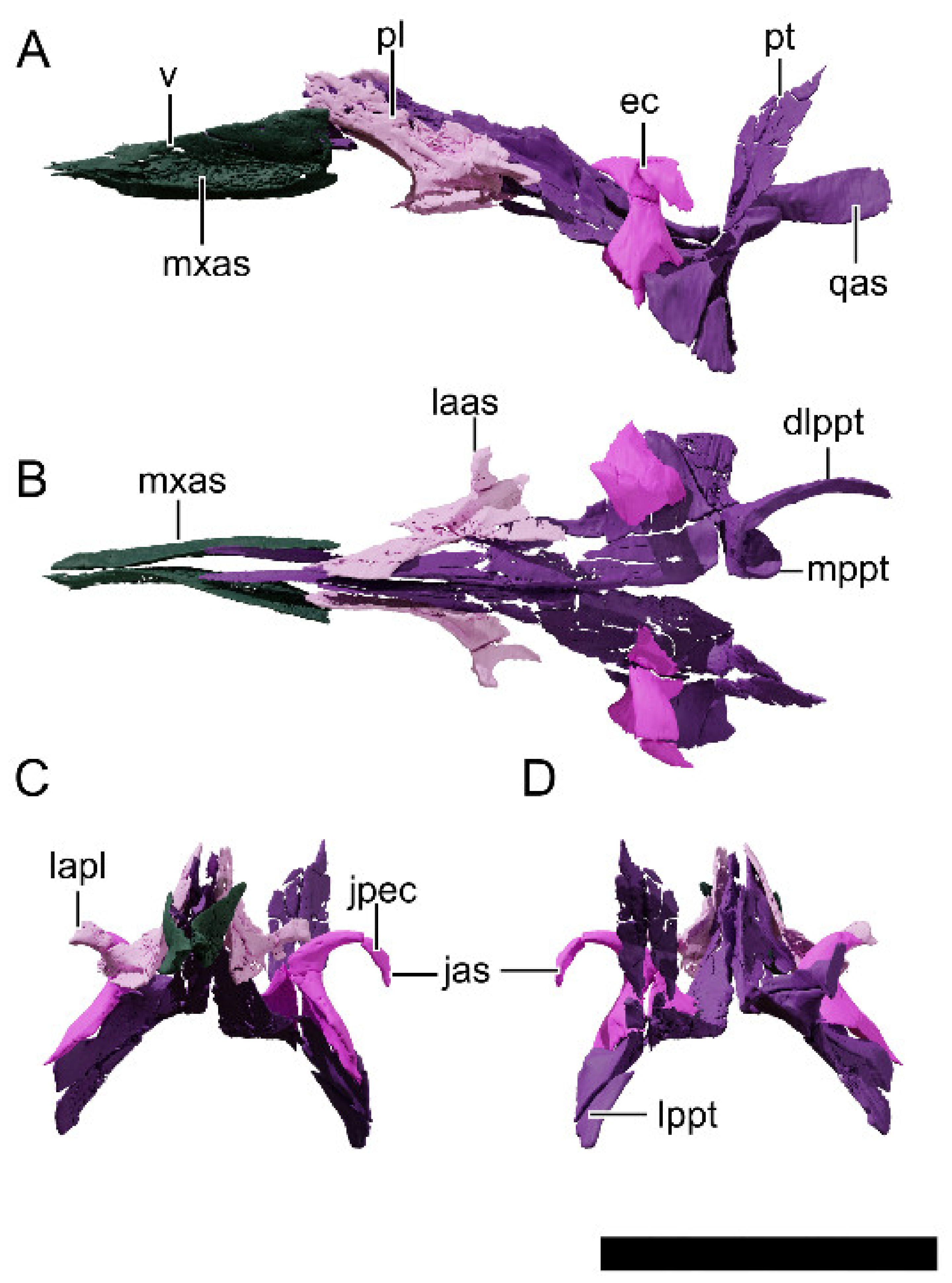
4.8.19. Palate
4.8.20. Pterygoid
4.8.21. Ectopterygoid
4.8.22. Palatine
4.8.23. Vomer
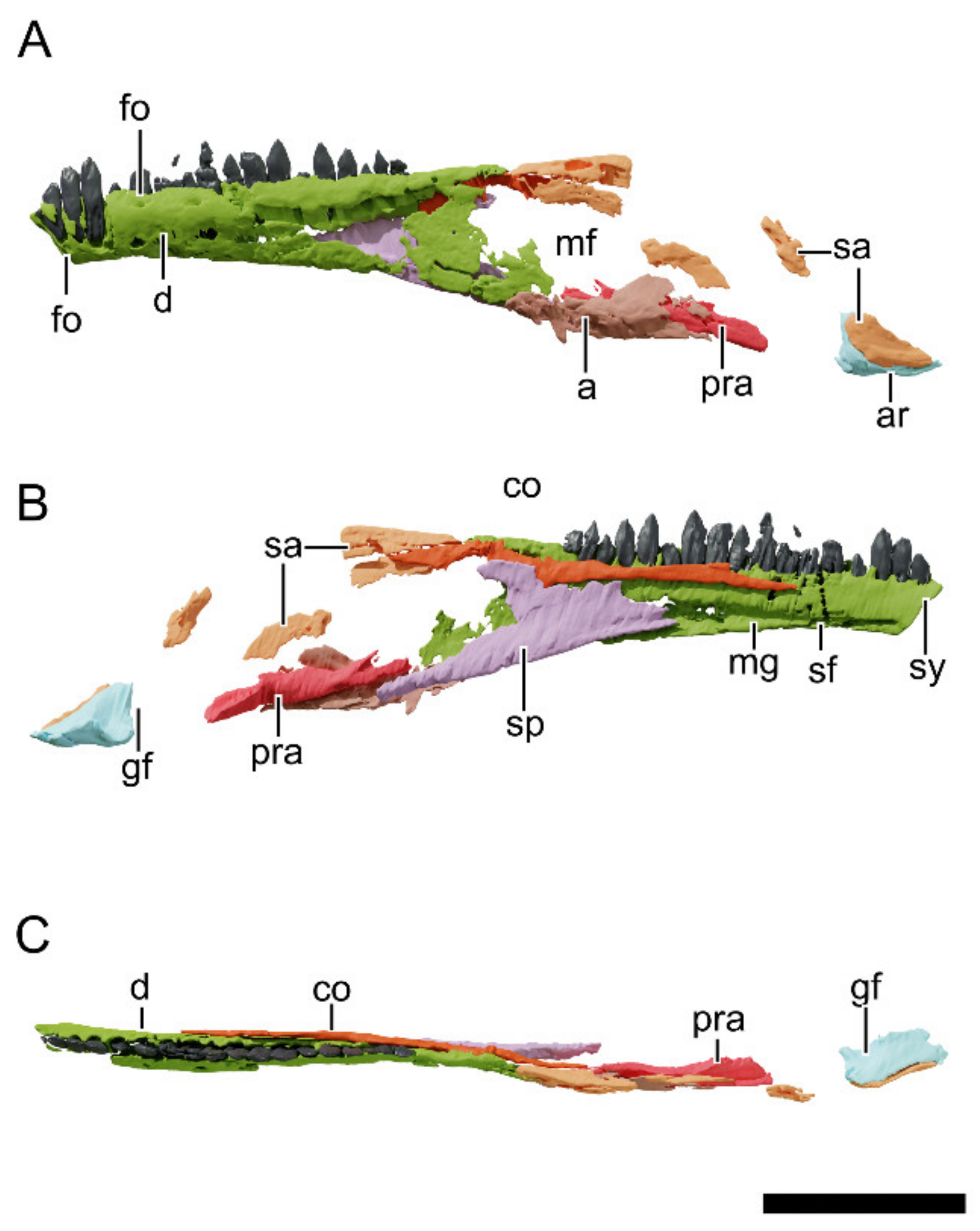
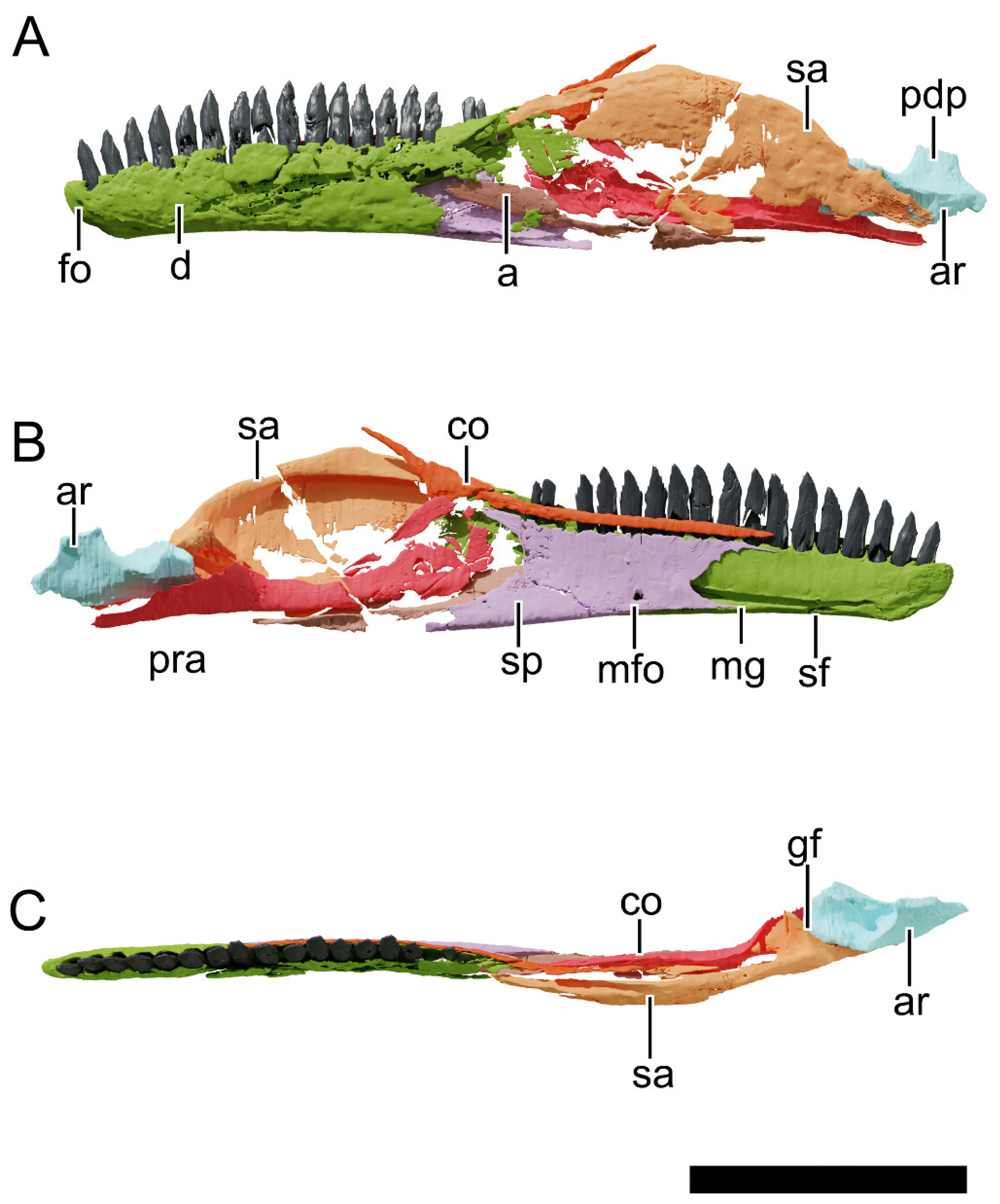
4.8.24. Dentary
4.8.25. Splenial
4.8.26. Intercoronoid/Coronoid
4.8.27. Surangular
4.8.28. Angular
4.8.29. Prearticular
4.8.30. Articular
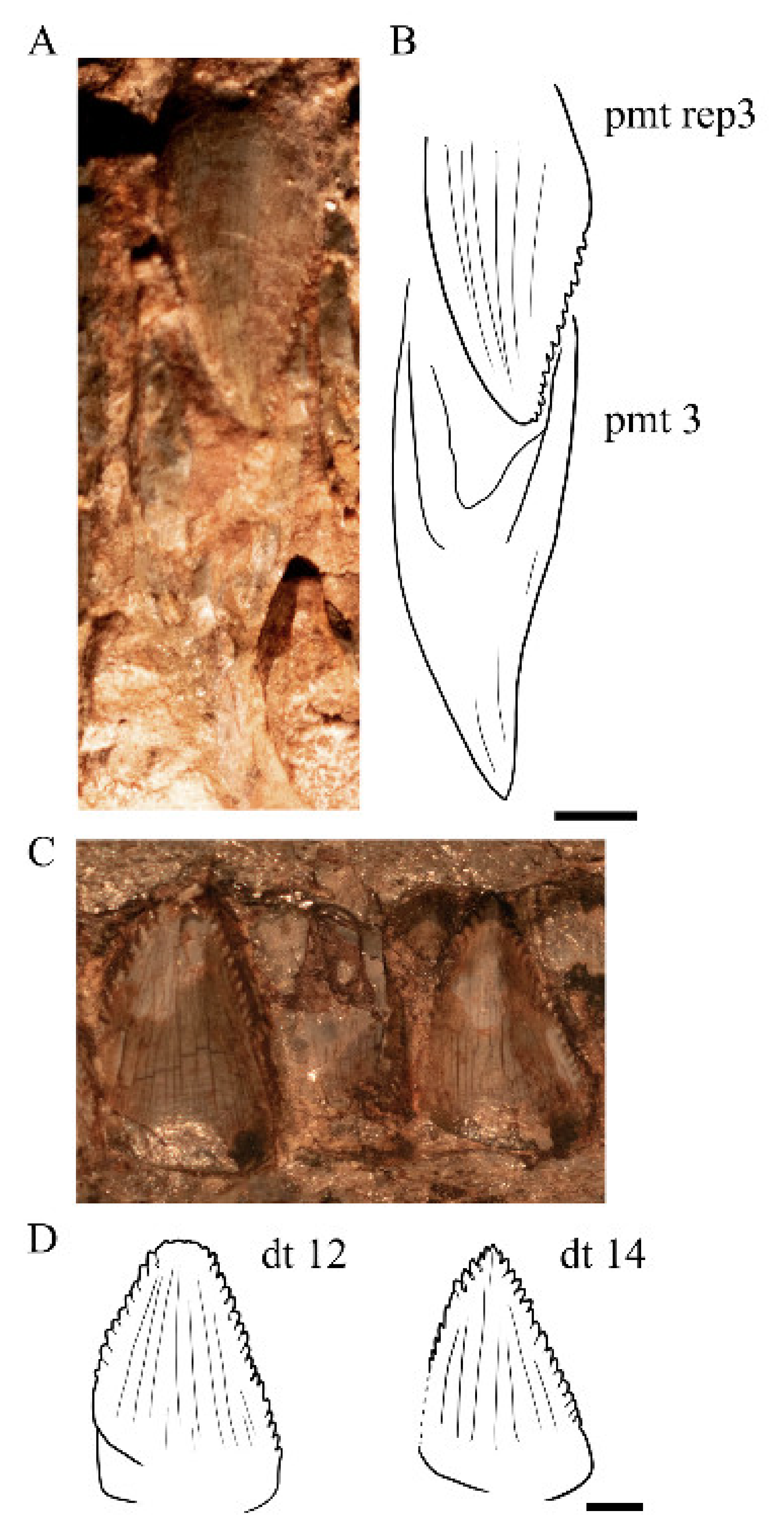
4.8.31. Dentition
4.8.32. Hyoid
4.8.33. Sclerotic Ring
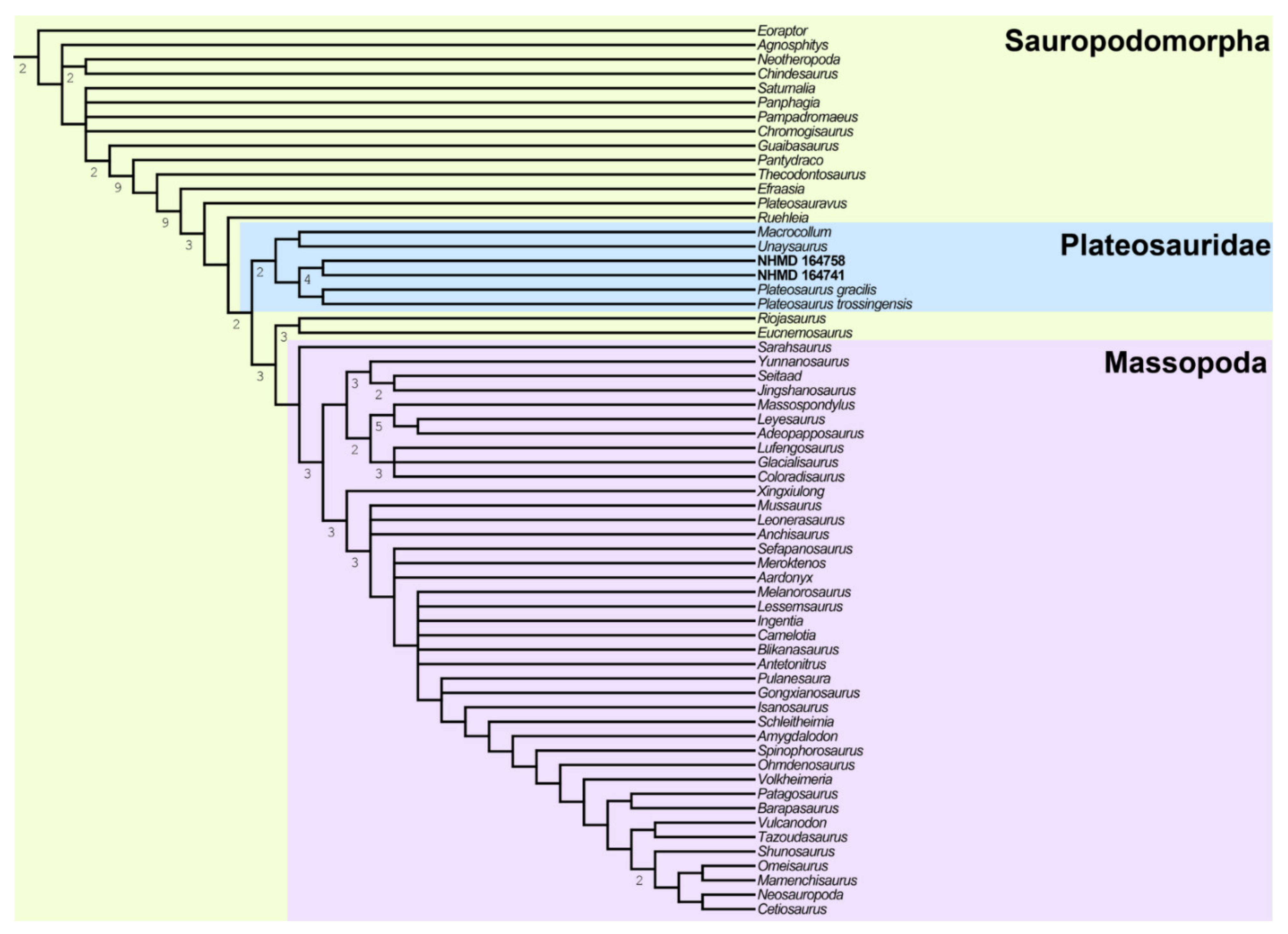
4.9. Phylogenetic Analysis
5. Discussion
5.1. Arguments for a New Taxon and Comparisons
5.2. Arguments for a Single New Species
5.3. Ontogeny
5.4. Paleobiogeographic and Chronological Implications
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyer, H. Briefliche mitteilung an Prof. Bronn über Plateosaurus engelhardti. Neues Jahrb. Mineral. Geogn. Geol. Petrefakten-Kunde 1837, 24, 314–316. [Google Scholar]
- Yates, A.M. The species taxonomy of the sauropodomorph dinosaurs from the Lowenstein Formation (Norian, Late Triassic) of Germany. Palaeontology 2003, 46, 317–337. [Google Scholar] [CrossRef]
- Galton, P.M. Prosauropod Dinosaur Plateosaurus (=Gresslyosaurus) (Saurischia: Sauropodomorpha) from the Upper Triassic of Switzerland. Geol. Palaeontol. 1986, 20, 167–183. [Google Scholar]
- Rütimeyer, L. Reptilien knochen aus dem Keuper von Liestal. Verh. Schweiz. Nat. Ges. 1856, 41, 62–64. [Google Scholar]
- Huene, F.F. Über die Trias-Dinosaurier Europas. Z. Dtsch. Geol. Ges. 1905, 57, 345–349. [Google Scholar]
- Huene, F.F. Die Dinosaurier der Europäischen Triasformation mit Berücksichtigung der Aussereuropäischen Vorkommnisse; G. Fischer: Jena, Germany, 1908; Volume 1. [Google Scholar]
- Fraas, E. Die neuesten Dinosaurierfunde in der Schwäbischen Trias. Naturwissenschaften 1913, 1, 1097–1100. [Google Scholar] [CrossRef] [Green Version]
- Yates, A.M. Solving a dinosaurian puzzle: The identity of Aliwalia Rex Galton. Hist. Biol. 2007, 19, 93–123. [Google Scholar] [CrossRef]
- Yates, A.M. A revision of the problematic sauropodomorph dinosaurs from Manchester, Connecticut and the Status of Anchisaurus Marsh: The taxonomic status of Anchisaurus. Palaeontology 2010, 53, 739–752. [Google Scholar] [CrossRef]
- Galton, P.M.; Kermack, D. The anatomy of Pantydraco Caducus, a very basal Sauropodomorph Dinosaur from the Rhaetian (Upper Triassic) of South Wales, UK. Rev. Palóbiologie 2010, 29, 341–404. [Google Scholar]
- Apaldetti, C.; Martinez, R.N.; Alcober, O.A.; Pol, D. A new basal Sauropodomorph (Dinosauria: Saurischia) from Quebrada Del Barro formation (Marayes-El Carrizal Basin), Northwestern Argentina. PLoS ONE 2011, 6, e26964. [Google Scholar] [CrossRef]
- Rauhut, O.W.M.; Holwerda, F.M.; Furrer, H. A derived Sauropodiform Dinosaur and other sauropodomorph material from the Late Triassic of Canton Schaffhausen, Switzerland. Swiss J. Geosci. 2020, 113, 8. [Google Scholar] [CrossRef]
- Galton, P.M. Case 3560 Plateosaurus Engelhardti Meyer, 1837 (Dinosauria, Sauropodomorpha): Proposed replacement of unidentifiable name-bearing type by a neotype. Bull. Zool. Nomencl. 2012, 69, 203–212. [Google Scholar] [CrossRef]
- ICZN. Opinion 2435 (Case 3560) Plateosaurus Meyer, 1837 (Dinosauria, Sauropodomorpha): New type species designated. Bull. Zool. Nomencl. 2019, 76, 144–145. [Google Scholar] [CrossRef]
- Yates, A.M. The first complete skull of the triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria). Spec. Pap. Palaeontol. 2007, 77, 9–55. [Google Scholar]
- Chapelle, K.E.J.; Choiniere, J.N. A revised cranial description of Massospondylus Carinatus owen (Dinosauria: Sauropodomorpha) based on computed tomographic scans and a review of cranial characters for basal Sauropodomorpha. PeerJ 2018, 6, e4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapelle, K.E.J.; Barrett, P.M.; Botha, J.; Choiniere, J.N. Ngwevu Intloko: A new early sauropodomorph dinosaur from the lower jurassic elliot formation of South Africa and comments on cranial ontogeny in Massospondylus carinatus. PeerJ 2019, 7, e7240. [Google Scholar] [CrossRef] [Green Version]
- McPhee, B.W.; Bittencourt, J.S.; Langer, M.C.; Apaldetti, C.; Da Rosa, Á.A.S. Reassessment of Unaysaurus Tolentinoi (Dinosauria: Sauropodomorpha) from the Late Triassic (Early Norian) of Brazil, with a consideration of the evidence for monophyly within non-sauropodan sauropodomorphs. J. Syst. Palaeontol. 2020, 18, 259–293. [Google Scholar] [CrossRef]
- Müller, R.T. Craniomandibular osteology of Macrocollum Itaquii (Dinosauria: Sauropodomorpha) from the Late Triassic of Southern Brazil. J. Syst. Palaeontol. 2020, 18, 805–841. [Google Scholar] [CrossRef]
- Leal, L.A.; Azevedo, S.A.K.; Kellner, A.W.A.; Rosa, Á.A.S. A new early dinosaur (Sauropodomorpha) from the Caturrita Formation (Late Triassic), Paraná Basin, Brazil. Zootaxa 2004, 690, 1–24. [Google Scholar] [CrossRef]
- McPhee, B.W.; Choiniere, J.N.; Yates, A.M.; Viglietti, P.A. A second species of Eucnemesaurus Van Hoepen, 1920 (Dinosauria, Sauropodomorpha): New information on the diversity and evolution of the sauropodomorph fauna of South Africa’s Lower Elliot Formation (Latest Triassic). J. Vertebr. Paleontol. 2015, 35, e980504. [Google Scholar] [CrossRef]
- Müller, R.T.; Langer, M.C.; Dias-da-Silva, S. An exceptionally preserved association of complete dinosaur skeletons reveals the oldest long-necked sauropodomorphs. Biol. Lett. 2018, 14, 20180633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novas, F.E.; Ezcurra, M.D.; Chatterjee, S.; Kutty, T.S. New dinosaur species from the upper Triassic upper Maleri and lower Dharmaram formations of Central India. Earth Environ. Sci. Trans. R. Soc. Edinb. 2011, 101, 333–349. [Google Scholar] [CrossRef]
- Novas, F.E.; Ezcurra, M.D.; Chatterjee, S.; Kutty, T.S. Late Triassic Continental Vertebrates and Depositional Environments of the Fleming Fjord Formation, Jameson Land, East. Greenland; Jenkins, F.A., Shubin, N.H., Amaral, W.W., Gatesy, S.M., Schaff, C.R., Clemmensen, L.B., Downs, W.R., Davidson, A.R., Bonde, N., Osbaeck, F., Eds.; Meddelelser om Grønland Geoscience: Brenderup, Denmark, 1994; ISBN 978-87-601-4573-5. [Google Scholar]
- Clemmensen, L.B.; Kent, D.W.; Mau, M.; Mateus, O.; Milàn, J. Triassic lithostratigraphy of the Jameson land basin (Central East Greenland), with emphasis on the New Fleming Fjord Group. Bull. Geol. Soc. Den. 2020, 68, 95–132. [Google Scholar] [CrossRef]
- Marzola, M.; Mateus, O.; Milàn, J.; Clemmensen, L.B. A review of Palaeozoic and Mesozoic Tetrapods from Greenland. Bull. Geol. Soc. Den. 2018, 66, 21–46. [Google Scholar] [CrossRef]
- Guarnieri, P.; Brethes, A.; Rasmussen, T.M. Geometry and kinematics of the Triassic rift basin in Jameson Land (East Greenland): Triassic rift basin East Greenland. Tectonics 2017, 36, 602–614. [Google Scholar] [CrossRef]
- Kent, D.V.; Clemmensen, L.B. Northward dispersal of dinosaurs from Gondwana to Greenland at the Mid-Norian (215–212 Ma, Late Triassic) dip in atmospheric pCO2. Proc. Natl. Acad. Sci. USA 2021, 118, e2020778118. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, L.B.; Kent, D.V.; Jenkins, F.A. A late triassic lake system in East Greenland: Facies, depositional cycles and palaeoclimate. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 140, 135–159. [Google Scholar] [CrossRef] [Green Version]
- Sellwood, B.W.; Valdes, P.J. Mesozoic climates: General circulation models and the rock record. Sediment. Geol. 2006, 190, 269–287. [Google Scholar] [CrossRef]
- Agnolin, F.L.; Mateus, O.; Milàn, J.; Marzola, M.; Wings, O.; Adolfssen, J.S.; Clemmensen, L.B. Ceratodus Tunuensis, Sp. Nov., a new lungfish (Sarcopterygii, Dipnoi) from the Upper Triassic of Central East Greenland. J. Vertebr. Paleontol. 2018, 38, e1439834. [Google Scholar] [CrossRef]
- Sulej, T.; Wolniewicz, A.; Bonde, N.; Błażejowski, B.; Niedźwiedzki, G.; Tałanda, M. New perspectives on the Late Triassic vertebrates of East Greenland: Preliminary results of a Polish−Danish palaeontological expedition. Pol. Polar Res. 2014, 35, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Clemmensen, L.B.; Milàn, J.; Adolfssen, J.S.; Estrup, E.J.; Frobøse, N.; Klein, N.; Mateus, O.; Wings, O. The vertebrate-bearing Late Triassic Fleming Fjord Formation of Central East Greenland revisited: Stratigraphy, palaeoclimate and new palaeontological data. Geol. Soc. Lond. Spec. Publ. 2016, 434, 31–47. [Google Scholar] [CrossRef]
- Marzola, M.; Mateus, O.; Shubin, N.H.; Clemmensen, L.B. Cyclotosaurus Naraserluki Sp. Nov., a new Late Triassic Cyclotosaurid (Amphibia, Temnospondyli) from the Fleming Fjord Formation of the Jameson Land Basin (East Greenland). J. Vertebr. Paleontol. 2017, 37, e1303501. [Google Scholar] [CrossRef] [Green Version]
- Niedźwiedzki, G.; Sulej, T. Theropod dinosaur fossils from the Gipsdalen and Fleming fjord formations (Carnian-Norian, Upper Triassic), East Greenland. In Proceedings of the 34th Nordic Geological Winter Meeting, Oslo, Norway, 8–10 January 2020; Nakrem, H.A., Hus, A.M., Eds.; Geological Society of Norway: Oslo, Norway, 2020; Volume 2020, p. 151. [Google Scholar]
- Milàn, J.; Octávio, M.; Mau, M.; Rudra, A.; Sanei, H.; Clemmensen, L.B. A possible phytosaurian (Archosauria, Pseudosuchia) coprolite from the Late Triassic Fleming Fjord Group of Jameson Land, Central East Greenland. Bull. Geol. Soc. Den. 2021, 69, 71–80. [Google Scholar] [CrossRef]
- Jenkins, F.A.; Gatesy, S.M.; Shubin, N.H.; Amaral, W.W. Haramiyids and Triassic mammalian evolution. Nature 1997, 385, 715–718. [Google Scholar] [CrossRef]
- Sulej, T.; Krzesiński, G.; Tałanda, M.; Wolniewicz, A.S.; Błażejowski, B.; Bonde, N.; Gutowski, P.; Sienkiewicz, M.; Niedźwiedzki, G. The earliest-known mammaliaform fossil from Greenland sheds light on origin of mammals. Proc. Natl. Acad. Sci. USA 2020, 117, 26861–26867. [Google Scholar] [CrossRef] [PubMed]
- Pretto, F.A.; Langer, M.C.; Schultz, C.L. A new dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Brazil provides insights on the evolution of Sauropodomorph body plan. Zool. J. Linn. Soc. 2019, 185, 388–416. [Google Scholar] [CrossRef] [Green Version]
- Cabreira, S.F.; Kellner, A.W.A.; Dias-da-Silva, S.; Roberto da Silva, L.; Bronzati, M.; de Almeida Marsola, J.C.; Müller, R.T.; de Souza Bittencourt, J.; Batista, B.J.; Raugust, T.; et al. A unique Late Triassic Dinosauromorph assemblage reveals dinosaur ancestral anatomy and Diet. Curr. Biol. 2016, 26, 3090–3095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, R.T.; Langer, M.C.; Bronzati, M.; Pacheco, C.P.; Cabreira, S.F.; Dias-da-Silva, S. Early evolution of Sauropodomorphs: Anatomy and phylogenetic relationships of a remarkably well-preserved dinosaur from the Upper Triassic of Southern Brazil. Zool. J. Linn. Soc. 2018, 184, 1187–1248. [Google Scholar] [CrossRef] [Green Version]
- Müller, R.T.; Ferreira, J.D.; Pretto, F.A.; Bronzati, M.; Kerber, L. The endocranial anatomy of Buriolestes Schultzi (Dinosauria: Saurischia) and the early evolution of brain tissues in sauropodomorph dinosaurs. J. Anat. 2021, 238, 809–827. [Google Scholar] [CrossRef]
- Bonaparte, J.F. Coloradia Brevis n. g. et n. Sp. (Saurischia, Prosauropoda), a Plateosaurid dinosaur from the Los Colorados Formation, Upper Triassic of La Rioja, Argentina. Ameghiniana 1978, 15, 7. [Google Scholar]
- Apaldetti, C.; Martinez, R.N.; Pol, D.; Souter, T. Redescription of the skull of Coloradisaurus Brevis (Dinosauria, Sauropodomorpha) from the Late Triassic Los Colorados Formation of the Ischigualasto-Villa Union Basin, Northwestern Argentina. J. Vertebr. Paleontol. 2014, 34, 1113–1132. [Google Scholar] [CrossRef]
- Galton, P.M. Cranial anatomy of the prosauropod dinosaur Plateosaurus from the Knollenmergel (Middle Keuper, Upper Triassic) of Germany. II. All the cranial material and details of soft-part anatomy. Geol. Palaeontol. 1985, 19, 119–159. [Google Scholar]
- Bronzati, M.; Rauhut, O.W.M. Braincase redescription of Efraasia Minor. Huene, 1908 (Dinosauria: Sauropodomorpha) from the Late Triassic of Germany, with comments on the evolution of the sauropodomorph braincase. Zool. J. Linn. Soc. 2018, 182, 173–224. [Google Scholar] [CrossRef]
- Barrett, P.M.; Upchurch, P.; Xiao-Lin, W. Cranial osteology of Lufengosaurus Huenei young (Dinosauria: Prosauropoda) from the lower jurassic of Yunnan, People’s Republic of China. J. Vertebr. Paleontol. 2005, 25, 806–822. [Google Scholar] [CrossRef]
- Barrett, P.M. A new basal sauropodomorph dinosaur from the Upper Elliot Formation (Lower Jurassic) of South Africa. J. Vertebr. Paleontol. 2009, 29, 1032–1045. [Google Scholar] [CrossRef]
- Langer, M.C.; McPhee, B.W.; Marsola, J.C.D.A.; Roberto-Da-Silva, L.; Cabreira, S.F. Anatomy of the dinosaur Pampadromaeus Barberenai (Saurischia—Sauropodomorpha) from the Late Triassic Santa Maria Formation of Southern Brazil. PLoS ONE 2019, 14, e0212543. [Google Scholar] [CrossRef]
- Martinez, R.N.; Alcober, O.A. A Basal Sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS ONE 2009, 4, e4397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galton, P.M. The Prosauropod Dinosaur Plateosaurus Meyer, 1837 (Saurischia: Sauropodomorpha; Upper Triassic). II. Notes on the referred species. Rev. Paléobiologie 2001, 20, 435–502. [Google Scholar]
- Prieto-Márquez, A.; Norell, M.A. Redescription of a Nearly complete skull of Plateosaurus (Dinosauria: Sauropodomorpha) from the Late Triassic of Trossingen (Germany). Am. Mus. Novit. 2011, 3727, 1–58. [Google Scholar] [CrossRef]
- Lallensack, J.N.; Teschner, E.; Pabst, B.; Sander, P.M. New skulls of the Basal Sauropodomorph Plateosaurus Trossingensis from Frick, Switzerland: Is there more than one species? Acta Palaeontol. Pol. 2021, 66, 1–28. [Google Scholar] [CrossRef]
- Langer, M.C.; Abdala, F.; Richter, M.; Benton, M.J. A Sauropodomorph dinosaur from the Upper Triassic (Carnian) of Southern Brazil. Académie Sci. 1999, 329, 511–517. [Google Scholar]
- Mallison, H.; Wings, O. Photogrammetry in paleontology—A practical guide. J. Paleontol. Tech. 2014, 12, 1–31. [Google Scholar]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Owen, R. Report on British Fossil Reptiles. Part. II; Report of the British Association for the Advancement of Science; British Association for the Advancement of Science: London, UK, 1842. [Google Scholar]
- Seeley, H.G. I. On the classification of the fossil animals commonly named Dinosauria. Proc. R. Soc. Lond. 1887, 43, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Huene, F.F. Die Fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte; Verlag von Gebrüder Borntraeger: Leipzic, Germany, 1932. [Google Scholar]
- Marsh, O.C. On the affinities and classification of the dinosaurian reptiles. Am. J. Sci. 1895, 3, 483–498. [Google Scholar] [CrossRef]
- Clemmensen, L.B. Triassic lithostratigraphy of East Greenland between Scoresby Sund and Kejser Franz Josephs Fjord. Grønlands Geol. Undersøgelse Bull. 1980, 139, 56. [Google Scholar]
- Young, C.-C. A Complete Osteology of Lufengosaurus Heunei Young (Gen. et Sp. Nov.) from Lufeng, Yunnan, China. Geol. Surv. China 1941, 7, 53. [Google Scholar]
- Galton, P.M.; Upchurch, P. 12. Prosauropoda. In The Dinosauria, 2nd ed.; University of California Press: Berkeley, CA, USA, 2004; pp. 232–258. [Google Scholar]
- Sander, P.M.; Klein, N. Developmental plasticity in the life history of a prosauropod dinosaur. Science 2005, 310, 1800–1802. [Google Scholar] [CrossRef] [PubMed]
- Galton, P.M. Cranial anatomy of the prosauropod dinosaur Plateosaurus from the Knollenmergel (Middle Keuper, Upper Triassic) of Germany. I. Two complete skulls from Trossingen/Württ. with comments on the diet. Geol. Palaeontol. 1984, 18, 139–171. [Google Scholar]
- Klein, N.; Sander, P.M. Bone histology and growth of the prosauropod dinosaur Plateosaurus Engelhardti von Meyer, 1837 from the Norian Bonebeds of Trossingen (Germany) and Frick (Switzerland). Spec. Pap. Palaeontol. 2007, 77, 169–206. [Google Scholar]
- Sander, P.M. The norian Plateosaurus Bonebeds of Central Europe and Their yaphonomy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1992, 93, 255–299. [Google Scholar] [CrossRef]
- Moser, M. Plateosaurus engelhardti Meyer, 1837 (Dinosauria: Sauropodomorpha) aus dem Feuerletten (Mittelkeuper; Obertrias) von Bayern. Zitteliana 2003, 24, 3–186. [Google Scholar]
- Hurum, J.H.; Bergan, M.; Müller, R.; Nystuen, J.P.; Klein, N. A Late Triassic dinosaur bone, Offshore Norway. Norwigian J. Geol. 2006, 86, 117–123. [Google Scholar]
- Mallison, H. The digital Plateosaurus I: Body mass, mass distribution and posture assessed using Cad and Cae on a digitally mounted complete skeleton. Palaeontol. Electron. 2010, 13, 26. [Google Scholar]
- Nau, D.; Lallensack, J.N.; Bachmann, U.; Sander, P.M. Postcranial osteology of the first early-stage juvenile skeleton of Plateosaurus Trossingensis (Norian, Frick, Switzerland). Acta Palaeontol. Pol. 2020, 65, 679–708. [Google Scholar] [CrossRef]
- Langer, M.C.; Ramezani, J.; Da Rosa, Á.A.S. U-Pb age constraints on dinosaur rise from South Brazil. Gondwana Res. 2018, 57, 133–140. [Google Scholar] [CrossRef]
- Nesbitt, S.J.; Irmis, R.B.; Parker, W.G. A Critical re-evaluation of the Late Triassic dinosaur taxa of North America. J. Syst. Palaeontol. 2007, 5, 209–243. [Google Scholar] [CrossRef] [Green Version]
- Marsh, O.C. Names of extinct reptiles. Am. J. Sci. 1885, 29, 169. [Google Scholar]
- Rowe, T.B.; Sues, H.-D.; Reisz, R.R. Dispersal and diversity in the earliest North American Sauropodomorph dinosaurs, with a description of a New Taxon. Proc. R. Soc. B 2011, 278, 1044–1053. [Google Scholar] [CrossRef] [Green Version]







































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
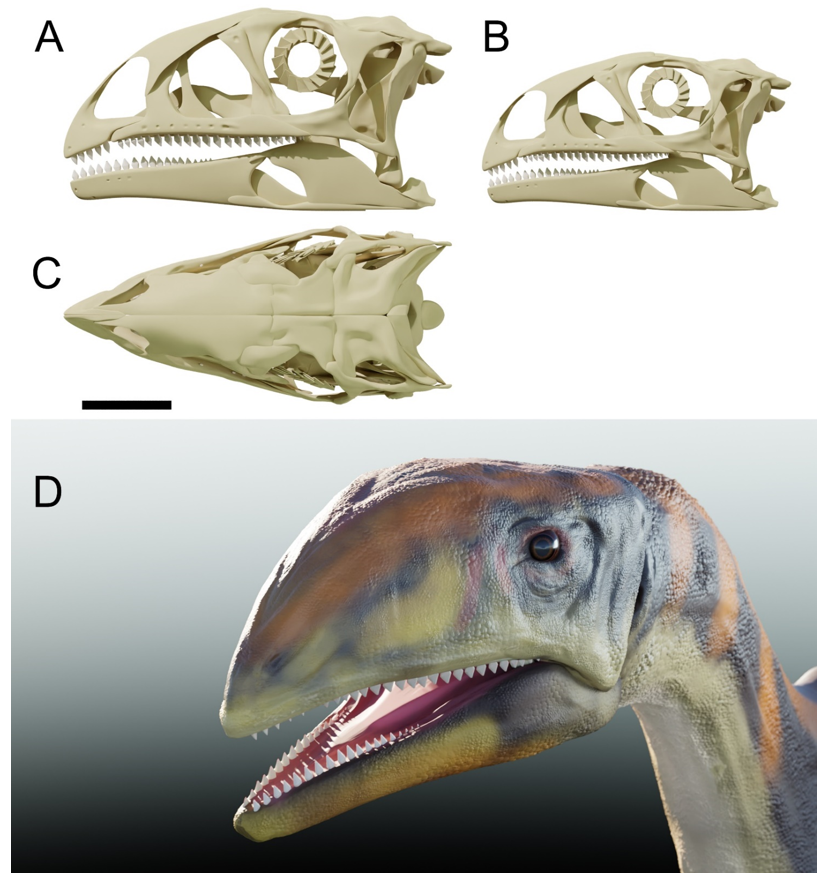{kind=link}
| Taxon | Specimen(s) | Source(s) |
|---|---|---|
| Bagualosaurus agudoensis | UFRGS-PV-1099-T | [39] |
| Buriolestes schultzi | CAPPA/UFSM 0035; ULBRA-PVT280 | [40,41,42] |
| Coloradisaurus brevis | PVL 3967 | [43,44] |
| Efraasia minor | SMNS 12667 | [2,45,46] |
| Leyesaurus marayensis | PVSJ 706 | [11] |
| Lufengosaurus huenei | IVPP V15 | [47] |
| Macrocollum itaquii | CAPPA/UFSM 0001a; CAPPA/UFSM 0001b | [19,22] |
| Massospondylus carinatus | BP/1/5241; BP/1/4934 | [16] |
| Massospondylus kaalae | SAM-PK-K1325 | [48] |
| Ngwevu intlokoi | BP/1/4779 | [17] |
| Pampadromaeus berberenai | ULBRA-PVT016 | [49] |
| Panphagia protos | PVSJ 874 | [50] |
| Plateosaurus trossingensis | AMNH FARB 6810; GPIT-PV-30704; MB.R.1937; MSF 07.M; MSF 08.M; MSF 08.H; MSF 09.2; MSF 11.4; MSF 12.3; MSF 15.4; MSF 15.8; MSF 16.1; MSF 17.4; MSF 23; MSF 33; NAAG_00011238; NAAG_ 00011239; SMA 09.1; SMNS 12949; SMNS 12950; SMNS 13200; SMNS 52968 | [2,3,51,52,53] |
| Plateosaurus gracilis | GPIT 18318a | [2] |
| Saturnalia tupiniquim | MCP 3845-PV | [54] |
| Measurements (in mm) | NHMD 164741 | NHMD 164758 |
|---|---|---|
| Skull anteroposterior length (from the anterior tip of the premaxilla to posterior margin of occipital condyle) | 243.7 * | 167.9 |
| Skull maximum dorsoventral height at orbit including the mandible | 112.7 | 93.6 |
| Skull dorsoventral height at orbit excluding mandible | 75.2 | 58.3 |
| Rostrum dorsoventral height (measured at the posterior margin of the external naris level) | 57.9 | 40.9 |
| External naris maximum anteroposterior length | 35.1 | 25.1 |
| Orbit length | 52.5 * | 49.9 |
| Orbit height | 46.5 * | 41.0 |
| Premaxilla maximum anteroposterior length | - | 36.9 |
| Premaxilla alveolar anteroposterior length | - | 24.3 |
| Premaxilla maximum dorsoventral height | 40.3 | 37.8 |
| Narial fossa anteroposterior length | 28.7 | 24.9 |
| Maxilla anteroposterior maximum length | 129.9 | 95.5 |
| Maxilla anteroposterior alveolar length | 118.8 | 85.7 |
| Maxilla dorsoventral height (from the dorsal tip of the dorsal process to ventral margin of maxilla) | 57.0 | 42.0 |
| Antorbital fossa maximum anteroposterior length | 46.5 | 34.8 |
| Nasal anteroposterior length | 98.7 | 80.0 |
| Lacrimal dorsoventral height | 64.5 | 41.9 |
| Lacrimal maximum length of dorsal region | 43.1 | 37.5 |
| Prefrontal dorsoventral maximum height | - | 32.3 |
| Frontal maximum anteroposterior length | 51.6 | 42.3 |
| Frontal maximum mediolateral width | 49.1 | 18.7 |
| Postorbital maximum length | 48.6 | 25.5 |
| Postorbital maximum height | 40.8 | 41.6 |
| Squamosal maximum length | 49.5 | - |
| Squamosal maximum height | 38.9 | - |
| Jugal maximum anteroposterior length | 57.8 | 48.9 |
| Jugal height under the orbit | 11.3 | 10.5 |
| Quadrate dorsoventral height | 91.8 | - |
| Quadrate mediolateral width at condylar region | 13.9 | 10.8 |
| Quadrate pterygoid flange anteroposterior length | 24.3 | 17.8 |
| Pterygoid maximum anteroposterior length | 92.4 | 83.8 |
| Pterygoid maximum dorsoventral height | 52.4 | 46.6 |
| Ectopterygoid maximum length of medial flange | 29.8 | 20.5 |
| Ectopterygoid maximum mediolateral width | 22.4 | 21.6 |
| Palatine maximum anteroposterior length | 31.3 | 39.8 |
| Palatine maximum mediolateral width | 9.1 | 13.2 |
| Vomer maximum anteroposterior length | - | 42.6 |
| Mandible maximum length | 210.1 * | 167.1 |
| Mandible maximum height | 45.6 | 33.8 |
| Dentary maximum length | 118.0 * | 100.1 |
| Dentary maximum height | 34.4 | 22.1 |
| Surangular maximum length | 118.1 | 79.8 |
| Surangular maximum height | 44.8 | 25.5 |
| Squamosal—Posterior Process Length to Anterolateral Process Length | ||
|---|---|---|
| Issi saaneq | NHMD 164741 | 1.31 |
| Pl. trossingensis | AMNH FARB 6810 | 0.55 |
| Pl. trossingensis | MSF 16.1 | 0.41 |
| Pl. trossingensis | NAAG_00011238 | 0.48 |
| Pl. trossingensis | MSF 12.3 | 0.53 |
| Pl. trossingensis | MSF 15.4 | 0.83 |
| Mac. itaquii | CAPPA/UFSM 0001b | 0.77 |
| Mas. carinatus | BP/1/5241 | 0.86 |
| Bu. schultzi | CAPPA/UFSM 0035 | 0.88 |
| Quadrate—Dorsoventral Height to Rostrum Dorsoventral Height | ||
|---|---|---|
| Issi saaneq | NHMD 164741 | 1.57 |
| Pl. trossingensis | AMNH FARB 6810 | 1.18 |
| Pl. trossingensis | NAAG_00011238 | 1.30 |
| Pl. trossingensis | MSF 11.4 | 1.27 |
| Pl. trossingensis | MSF 15.4 | 1.13 |
| Mac. itaquii | CAPPA/UFSM 0001a | 1.28 |
| Bu. schultzi | CAPPA/UFSM 0035 | 1.40 |
| Mas. carinatus | BP/1/5241 | 1.12 |
| N. intlokoi | BP/1/4779 | 1.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beccari, V.; Mateus, O.; Wings, O.; Milàn, J.; Clemmensen, L.B. Issi saaneq gen. et sp. nov.—A New Sauropodomorph Dinosaur from the Late Triassic (Norian) of Jameson Land, Central East Greenland. Diversity 2021, 13, 561. https://doi.org/10.3390/d13110561
Beccari V, Mateus O, Wings O, Milàn J, Clemmensen LB. Issi saaneq gen. et sp. nov.—A New Sauropodomorph Dinosaur from the Late Triassic (Norian) of Jameson Land, Central East Greenland. Diversity. 2021; 13(11):561. https://doi.org/10.3390/d13110561
Chicago/Turabian StyleBeccari, Victor, Octávio Mateus, Oliver Wings, Jesper Milàn, and Lars B. Clemmensen. 2021. "Issi saaneq gen. et sp. nov.—A New Sauropodomorph Dinosaur from the Late Triassic (Norian) of Jameson Land, Central East Greenland" Diversity 13, no. 11: 561. https://doi.org/10.3390/d13110561
APA StyleBeccari, V., Mateus, O., Wings, O., Milàn, J., & Clemmensen, L. B. (2021). Issi saaneq gen. et sp. nov.—A New Sauropodomorph Dinosaur from the Late Triassic (Norian) of Jameson Land, Central East Greenland. Diversity, 13(11), 561. https://doi.org/10.3390/d13110561