Appendix A
Character Descriptions
State (0) is reserved for absence. Citations are provided where significant overlap with a previously created character has occurred and/or where the character has been modified based on assessment of previously created characters. Citations of previously created characters are not meant to represent a comprehensive list of character overlap, and largely comprise the characters (and citations therein) of Musser and Cracraft [
44], due to our building on this dataset. Characters are anglicized as much as possible.
Skull: Rostrum
1. Premaxilla, dorsal aspect of rostral apex, distinct pair of foramina: absent (0); present (1); present, with associated rostral sulcus (2). Musser and Cracraft [
44], character 1.
2. Premaxilla, rostral apex, curvature: slightly curved ventrally (1); markedly curved inferiorly, hooked (2); lacking inferior curvature so that bill appears to be flat or recurved superiorly (3). Musser and Cracraft [
44], character 4.
3. Premaxilla, rostral apex, exceptionally mediolaterally wide: no (1); yes (2).
4. Premaxilla, processus frontalis, dorsally inflated: no (1); yes (2).
5. Premaxilla, processus frontalis, caudal terminus and associated suture between premaxilla and nasals: triangular (1); rounded (2). Noncomparable where suture obscured by nasal and/or frontal.
6. Rostrum, curvature, not including anterior terminus of premaxilla: lacking inferior curvature so that bill appears to be flat or recurved superiorly (1); curved inferiorly (2); curved extremely inferiorly, distal half of rostrum essentially perpendicular to proximal half (3); distal half of rostrum recurved dorsally (4).
7. Rostrum, dorsoventrally compressed so that maxillary process of nasal bar and ventral bar of rostrum almost at same level as processus frontalis of premaxilla: no (1); yes (2). Musser and Cracraft [
44], character 10; Livezey and Zusi [
95], character 280.
8. Rostrum, ventral aspect, tomial crest, caudomedial extent and ventral enclosure of rostrum and consequent shape of ventromedial fenestra: almost completely fused along midline to border of antorbital angle so that rostrum essentially closed by tomial crest ventrally, slit-like fenestra along midline may be visible in ventral aspect (1); fused along about 1/2 of midline or less near rostral apex so that pair of cylindrical cavities at apex visible but separated toward antorbital fenestra, creating “leaf-shaped” fenestra that is wider near antorbital fenestra (2); ventral enclosure of rostrum lost completely (3); like state 2 but over 1/2 of rostral portion is enclosed, pair of concavities concealed by tomial crest (4); completely closed ventrally by inflated bone (5).
9. Rostrum, tomial crest, lateral margins, ventral extension: essentially none, tomial crest at same level of ventral face of rostrum (1); marked caudal to apex (2); marked at rostral apex (3). Not applicable if rostrum not closed ventrally by tomial crest.
10. Nares, caudal margin: acuminate (1); rounded (2). Musser and Cracraft [
44], character 6.
11. Nares, length as compared to total length of rostrum: less than 1/2 of rostrum (1); 1/2 of rostrum (2); over 1/2 length of rostrum (3).
12. Nares, nasal sulcus beginning from rostral limit of nares: present, complete to terminal end of rostrum (1); lost or truncated (2). Musser and Cracraft [
44], character 9; Livezey and Zusi [
95], character 272.
13. Nares, rostral margin: rounded (1); acuminate (2).
14. Nares, internal bony septum ankylosed to ventral face of dorsal bar along midline: absent (0); present (1).
15. Nares, caudal terminal end, caudal margin relative to caudal margin of zona flexoria craniofacialis (defined here as the caudal margin of processus frontalis of premaxilla), site: rostral or subequal to rostral margin of zona flexoria craniofacialis, well within margins of flexor site (1); reach caudal margin or extend beyond caudal margin of zona flexoria craniofacialis so that premaxillary and maxillary processes of nasals essentially separated (2). Musser and Cracraft [
44], character 13; Livezey and Zusi [
95], character 341; Mayr and Clarke [
63], character 6.
16. Nares, caudal margin, thickness of dorsal limit of maxillary process of nasals compared to thickness of ventral limit: subequal in rostrocaudal width, narial bar often thin and rod like (1); wider at dorsal end (2); wider at ventral end (3).
17. Nasals, maxillary processes, ventral termini, lack of fusion to ventral rostrum: present (1); lost, fused (2).
18. Nasals, maxillary processes: rostrocaudally flattened (1); mediolaterally flattened (2); both, narial bars mediolaterally flattened rostrally and rostrocaudally flattened caudally (3).
19. Zona flexoria craniofacialis, depth: slight to moderate (1); deep, concave and fissure-like (2).
20. Zona flexoria craniofacialis, mediolateral alignment of caudal margin of processus frontalis of premaxilla and caudal margin of premaxillary (and maxillary, if separated as in schizorhiny) processes of nasal bone: absent, processus frontalis rostral to premaxillary processes of nasal bone (1); present, caudal termini of processes aligned mediolaterally (2); lost, caudal margin of processus frontalis of nasal bone extends further caudally than those of premaxillary processes of nasal bone (3). Not comparable when frontonasal suture indiscernible or where caudal termini of sutures are obscured by frontal bone. Musser and Cracraft [
44], character 14; Livezey and Zusi [
95], character 603.
21. Nasals and/or frontal, (typically) subtriangular depression just caudal to caudal limit of processus frontalis of premaxilla, status (regardless of furrowing of interorbital area): present (1); lost (2).
22. Nasals and/or frontal, rostral depression (if present), craniocaudal extent: limited within rostral half of orbit (1); extends caudally beyond rostral half of orbit (2). Noncomparable where rostral depression absent.
23. Frontal, dorsal face caudal to zona flexoria craniofacialis, dorsal inflation, status and magnitude: absent (1); present, extreme, extends caudally along frontal bone (2).
24. Frontal, rostrally protruding so that processus frontalis of premaxilla (and sometimes premaxillary processes of nasal bone) is obscured: no (1); yes, small, 1–2 rounded rostral bulb(s) protruding dorsal to proccessus frontalis of nasal bone (2); yes, mediolaterally wide, rostrally protruding lamina (3); yes, extremely dorsally inflated frontal bone protrudes dorsal to proccessus frontalis of nasal bone (4).
25. Rostrum, dorsal face, midline length from the apex of the premaxilla to the zona flexoria craniofacialis relative to that of the cranium: 0.40–2.00 (1); 2.01–3.00 (2). Musser and Cracraft [
44], character 2; Livezey and Zusi [
95], character 260.
26. Antorbital angle, general shape: subtriangular (1); ovoid, craniocaudally elongate (2).
27. Antorbital angle, angle created by ankylosis of maxillary process of nasal and maxilla: large, essentially 90 degrees (1); small, approximately 45 degrees (2). Noncomparable if antorbital fenestra is ovoid.
28. Mesethmoid, bifurcation and ankylosis to maxillary processes of nasals so that mesethmoid is inflated and visible within caudal portion of nares: no (1); yes (2).
29. Frontonasal suture, elevated dorsally: no (1); yes (2).
30. Orbital margin, salt gland sulcus: absent or non-distinct (0); present (1). Musser and Cracraft [
44], character 17; Mayr and Clarke [
63], character 25; Livezey [
113], character 72.
31. Orbital margin, salt gland sulcus (if present), depth: shallow (1); deep, typically mediolaterally expansive (2). Noncomparable where absent.
32. Interorbital area, midline: convex and not furrowed (1); furrowed along midline, concave (2). Musser and Cracraft [
44], character 18; Livezey [
113], character 70.
33. Cranium, dorsal aspect, fronto-parietal area, pair of prominent, horn-like protuberances: absent (0); present (1). Musser and Cracraft [
44], character 19.
34. Cranium (postorbital braincase), general form: craniocaudally elongate (1); craniocaudally compressed (2). Musser and Cracraft [
44], character 16; Livezey and Zusi [
95], character 7.
35. Frontoparietal suture, status in adults: present (1); lost (2). Musser and Cracraft [
44], character 20; Livezey and Zusi [
95], character 213.
36. Maxilla, tomial crest, tomial angle, extension of caudal terminal end as distinct tubercle or short process caudal to jugomaxillary suture and lateral to, and variably free from, arc of jugum: absent (0); present (1).
37. Maxilla, tomial crest, tomial angle, extension of caudal terminal end as distinct tubercle or short process caudal to jugomaxillary suture and lateral to, and variably free from, arc of jugum (if present), shape: triangular (1); subrectangular (2). Noncomparable where absent. Musser and Cracraft [
44], character 21; Livezey and Zusi [
95], character 408.
Skull: Cranium
38. Maxillopalatine processes, fused along midline: yes (1); no (2).
39. Maxillopalatine processes, shape of caudal terminus: acuminate (1); rounded (2).
40. Maxillopalatine processes, orientation of flattening: dorsoventrally flattened (1); mediolaterally flattened (2).
41. Maxillopalatine processes, craniocaudal elongation: present, extreme (1); lost (2); present, moderate (3).
42. Maxillopalatine processes, ventral margin, dorsoventral location relative to palatines: ventral or subequal to palatines (1); dorsal to palatines (2).
43. Maxillopalatine processes, extensive ankylosis to and/or contact with palatines: present (1); lost (2).
44. Maxillopalatine processes, enclosure of lateral (when mediolaterally flattened) or dorsal (when dorsoventrally flattened) face so that inflated, cone-like structure is created: not enclosed (1); partially enclosed (2); completely enclosed (3). Musser and Cracraft [
44], character 22; Livezey and Zusi [
95], character 420.
45. Maxillojugal pons, ventrolateral fenestra enclosed by caudal strut that is ankylosed to maxillopalatine processes and jugal: not enclosed (1); enclosed by bony projection (2). Musser and Cracraft [
44], character 12; Livezey and Zusi [
95], character 290.
46. Maxillojugal pons, craniocaudal attachment point of medial margin of strut: within rostral half of maxillopalatine processes (1); within caudal half of maxillopalatine processes (2). Noncomparable where maxillojugal pons absent.
47. Maxillojugal pons, caudal strut (if present), shape: craniocaudally thick, medial margin at least half craniocaudal length of lateral margin (1); tapered, lateral margin being craniocaudally more elongate than medial margin, medial portion of strut typically twisted rostrally (2). Noncomparable where maxillojugal pons absent.
48. Maxillojugal pons, ventrolateral fenestra, shape: small, circular (1); craniocaudally elongate, ovoid (2). Noncomparable where maxillojugal pons absent.
49. Orbital margin, crista-like orbital margin or supraorbital crest (see Musser and Cracraft [
44]): absent, orbital margins not laterally extended and are mostly or completely rounded (0); present (1). Musser and Cracraft [
44], character 24.
50. Orbital margin, supraorbital crest (if present): full, projecting dorsolaterally with a sharp, crista-like margin, creates a sharp and prominent supraorbital crest that extends from the postorbital process to the lacrimal (1); partial, like state only present along caudal half of orbit (2); crista-like margin prominent and present but projected ventrally/laterally, does not extend dorsal to frontal bone as in states 2 and 3 (3). Noncomparable where absent.
51. Orbital margin, pair of large, circular foramina near zona flexoria craniofacialis, typically within salt gland sulcus: absent (0); present (1).
52. Orbital margin, large pores: absent (0); present (1). Musser and Cracraft [
44], character 23.
53. Orbital margin, additional large foramina along orbital margin: absent (0); present (1).
54. Postorbital process, length: truncate (1); extremely elongate, subequal in length to height of quadrate (2). Musser and Cracraft [
44], character 26.
55. Postorbital process, orientation of apex: facing rostroventrally (1); facing ventrally (2).
56. Postorbital process, concavity: convex (1); concave along caudal length of process (2). Musser and Cracraft [
44], character 27.
57. Interorbital septum, mediolateral thickness: thin, in some places translucent (1); thick, opaque throughout (2). Musser and Cracraft [
44], character 28.
58. Lacrimofrontal suture: lost or indistinguishable (1); present (2).
59. Lacrimofrontal suture (if present), craniocaudal extent: incomplete, present in only rostral portion of lacrimal (1); present throughout dorsal margin of orbital process of lacrimal (2). Noncomparable where lacrimofrontal suture is lost or indistinguishable.
60. Lacrimal, site of ankylosis to nasals: ankylosed to both maxillary processes and frontal process of nasals and/or frontal bone (1); ankylosed to maxillary processes of nasals only (2); ankylosed to frontal process of nasals (3).
61. Lacrimals, site of ankylosis to nasals relative to zona flexoria craniofacialis (ZFC): at same level of ZFC (1); caudal to ZFC (2); rostral to ZFC (3).
62. Lacrimal, ankylosed to jugal or touching jugal bar: absent (0); present (1). Musser and Cracraft [
44], character 30; Livezey and Zusi [
95], character 195; Mayr and Clarke [
63], character 12.
63. Lacrimal, ankylosis to maxillopalatine processes/touching maxillopalatine processes: absent (0); present (1).
64. Lacrimal, supraorbital process, length: truncate (1); elongate (2).
65. Lacrimal, especially supraorbital process, inflated: no (1); yes (2).
66. Lacrimal, supraorbital process, general mediolateral width: narrow (1); wide (2).
67. Lacrimal, supraorbital process, gap present between interorbital area and caudal terminus of supraorbital process: no (1); yes (2). Musser and Cracraft [
44], character 31; Livezey and Zusi [
95], character 206.
68. Lacrimal, supraorbital process, tapered toward caudal apex: no (1); yes (2).
69. Lacrimal, supraorbital process, orientation of flattening: flattened dorsoventrally or craniocaudally (in line with coronal and/or transverse planes) (1); flattened mediolaterally (in line with parasagittal plane) (2); flattened ventrolaterally (3).
70. Lacrimal, rostral terminus of head of lacrimal, shape of apex: rounded or subrectangular (1); acuminate (2).
71. Lacrimal, rostral terminus of head of lacrimal, ankylosed to nasals: yes (1); no (2).
72. Lacrimal, rostral terminus of head of lacrimal, ankylosed to mesethmoid: no (1); yes (2).
73. Lacrimal, rostral terminus of head of lacrimal, extends rostrally with terminus medial to nasals: no (1); yes (2).
74. Lacrimal, additional rostral process along rostral margin of body ventral to rostral terminus of lacrimal head: absent (0); present (1).
75. Lacrimal, additional rostral process (if present), shape: rounded (1); acuminate (2); subrectangular (3). Noncomparable where absent and for Phoenicopterus due to incomplete ossification, and noncomparable where region fused to bill or mesethmoid or jugal.
76. Lacrimal, additional rostral process (if present), ankylosed to mesethmoid via bony strut: no (1); yes (2). Noncomparable where additional rostral process absent.
77. Lacrimal, body/orbital process, foramen: absent (0); present (1). Musser and Cracraft [
44], character 32; Livezey and Zusi [
95], character 197.
78. Lacrimal, descending process: wide and robust (1); narrow (2). Musser and Cracraft [
44], character 33; Clarke et al. [
114], character 11.
79. Lacrimal, descending process, formen or foramina: absent (0); present (1).
80. Lacrimal, descending process, convexity: concave (1); convex (2).
81. Lacrimal, descending process: elongate (1); truncate (2).
82. Lacrimal, ventral apex, shape: rounded or subrectangular (1); acuminate (2).
83. Lacrimal, ventral apex (or apices): terminal end(s) projects ventrally and/or caudally (1); terminal end(s) projects cranially (2). Musser and Cracraft [
44], character 36.
84. Ectethmoid, pneumatic foramen (exclusive of foramen orbitonasale laterale): absent (0); present, one (1); present, 2 or more (2). Noncomparable in absence of ectehmoid. Musser and Cracraft [
44], character 37.
85. Ectethmoid, size: absent (0); well developed (1); poorly developed (2). Musser and Cracraft [
44], character 38; Livezey and Zusi [
95], character 189; Mayr and Clarke [
63], character 14.
86. Ectethmoid, ankylosed to lacrimal: present (1); lost (2). Noncomparable where absent. Musser and Cracraft [
44], character 44.
87. Ectehmoid, foramen orbitonasalis medialis: v-shaped (1); small, circular (2); elongate, ovoid (3); lost (4). Noncomparable when absent due to lack of ankylosis of ectehmoid to ventral portion of antorbital area.
88. Olfactory sulcus, status and depth: shallow (1); deep (2).
89. Fonticulus interorbitalis, status: present (1); lost, only optic foramen present (2).
90. Fonticulus interorbitalis, craniocaudal extent: craniocaudally limited to caudal half of interorbital septum (1); craniocaudally extensive (2). Noncomparable where absent.
91. Fonticulus orbitocranialis, caudal extent: limited or essentially absent (1); extensive, continues along coronal plane of squamosal region (2).
92. Optic nerve foramen, completely confluent with closest lateral foramen/foramina (e.g., oculomotor nerve foramen, ophthalmic nerve foramen): no (1); yes (2).
93. Fonticulus interorbitalis and fonticulus orbitocranialis, confluent: no (1); yes (2). Noncomparable where absent.
94. Jugal, pronounced lateral bowing (convexity): absent, essentially straight (0); present and curved (1). Musser and Cracraft [
44], character 42; Livezey and Zusi [
95], character 298.
95. Jugal (especially maxillary portion), pronounced ventral bowing (convexity): absent (0); present (1). Musser and Cracraft [
44], character 43; Livezey and Zusi [
95], character 299.
96. Jugal, orientation of compression: absent, quadratojugal bar rounded (0); present, mediolateral compression evident so that bar is thin mediolaterally and thick dorsoventrally (1); present, dorsoventral compression evident so that bar is thin dorsoventrally and wide mediolaterally (2).
97. Jugal, slight medial bowing (concavity): absent (0); present (1).
98. Vomers, mediolaterally wide: yes (1); no (2). Mayr and Clarke [
63], character 20.
99. Vomers, forming a midline, narrow, and dorsoventrally high lamella: yes (1); no (2). Mayr and Clarke [
63], character 21.
100. Vomers, caudal ends not fused, more or less deeply cleft: no (1); yes (2). Mayr and Clarke [
63], character 19.
101. Palatines, contact with premaxilla or maxilla: palatines have contact with maxillae only (1); palatines have contact with premaxillae (2). Musser and Cracraft [
44], character 45; Cracraft and Clarke [
115], character 8.
102. Palatines, long and thin, especially anteriorly, poorly developed caudally, and widely separated anteriorly: absent (0); present (1).
103. Palatines, contact each other along midline along ventromedial margin: yes (1); no (2).
104. Palatines, ventral height of medial and lateral crests relative to each other: height of lateral crest greater than that of medial crest (1); heights subequal or crests barely visible (2); height of medial crest is greater than that of lateral crest (3). Musser and Cracraft [
44], character 46.
105. Palatines, inflated: no (1); yes (2).
106. Palatines, lateral portion: well-developed (1); rudimentary or vestigial (2). Musser and Cracraft [
44], character 47; Livezey and Zusi [
95], character 447; Mayr and Clarke [
63], character 16.
107. Palatines, lateral portion, marked caudolateral orientation: absent (0); present (1).
108. Palatines, ventrally raised oblique crest that extends from lateral to medial margins of ventral face: absent (0); present (1). Musser and Cracraft [
44], character 50; Livezey and Zusi [
95], character 457.
109. Palatines, ventrally raised oblique crest (if present), rostrocaudal location: located at level of caudolateral angle (1); located further rostrally, typically at rostrocaudal midpoint of palatines (2). Noncomparable where absent.
110. Palatines, caudomedial angle, pneumatic foramen: absent (0); present (1). Noncomparable where absent. Musser and Cracraft [
44], character 51.
111. Palatines, choanalis, ventral lamella, medial separation of bilateral lamellae: moderate (1); great (2). Musser and Cracraft [
44], character 53; Livezey and Zusi [
95], character 444.
112. Palatines, choanalis, ventral lamella, rostrocaudal position of caudomedial angle relative to that of caudolateral angle: coincident (1); rostral (2); caudal (3). Not comparable where caudolateral angle and/or caudomedial angle absent. Musser and Cracraft [
44], character 55.
113. Palatines, caudolateral angle: absent (0); present (1).
114. Palatines, caudolateral angle, position relative to area of ankylosis with pterygoids: rostral (1); subequal (2); caudal (3). Not comparable where caudolateral angle absent.
115. Palatines, caudolateral angle, shape: rounded (1); acuminate (2). Not comparable where caudolateral angle absent.
116. Palatines, pterygoid process, pneumatic foramen: present (1); lost (2). Not comparable in absence of pterygoid process. Musser and Cracraft [
44], character 57.
117. Pterygopalatine juncture, form: syndesmosis and pterygo-palatina propria, extensive rostrocaudally, caudal terminus approaching processus quadraticus pterygoidei (1); articulatio mesipterygo-palatina, with rudimentary gomphosis intrapterygoidea (2); articulatio pterygo-palatina simplex (3); articulatio mesipterygo-palatina, with complete gomphosis intrapterygoidea (4). Musser and Cracraft [
44], character 58; Livezey and Zusi [
95], character 601.
118. Palatines and pterygoids: sutured (1); segmented (articulated) (2). Musser and Cracraft [
44], character 59; Cracraft and Clarke [
115], character 6.
119. Pterygoid, facies articularis palatina, dorsoventral site of pterygopalatine juncture relative to parasphenoid rostrum (regardless of articulation via basipterygoid processes): slightly ventral, articulatio pterygo-rostroparasphenoidalis absent (1); on rostrum, articulatio pterygo-rostroparasphenoidalis present (2); markedly ventral, articulation of pterygo-rostroparasphenoidalis absent (3). Musser and Cracraft [
44], character 61; Livezey and Zusi [
95], character 480.
120. Pterygoid, rostral end markedly widened: absent (0); present (1). Noncomparable for Paleognathae. Bertelli et al. [
14], character 9; Mayr [
116].
121. Pterygoid, dorsomedial margin, craniocaudally elongate crista that is well projected dorsally and/or medially, may or may not have a facet for articulation with basipterygoid processes, rostrocaudal length: absent (0); present (1).
122. Pterygoid, dorsomedial margin, craniocaudally elongate crista that projects dorsally and/or medially (if present), rostrocaudal length: truncate, typically the length of roughly 1/4–1/2 of corpus (1); elongate, makes up entire length of pterygoid (2). Noncomparable where crista absent.
123. Pterygoid, dorsolateral margin, concavity: lost or slight (1); deep (2).
124. Pterygoid, ventromedial concavity: lost or slight (1); deep (2).
125. Pterygoid, area immediately rostromedial to area of articulation with quadrate, thin, accessory projection located on the dorsal face: absent (0); present (1).
126. Pterygoid, robust process that projects from medial margin of pterygoid medially towards basisphenoid for articulation with basipterygoid processes: absent (0); present (1).
127. Basisphenoid, basipterygoid processes: absent (0); present (1). Musser and Cracraft [
44], character 64; Cracraft and Clarke [
115], character 33.
128. Basisphenoid, basipterygoid processes, location: well rostral to hypoglossal canals (typically well rostral to basioccipital) (1); just rostral to hypoglossal canals (typically within margin of basioccipital) (2). Noncomparable where absent.
129. Basisphenoid, basipterygoid processes, ankylosed to pterygoids: yes (1); no (2). Noncomparable where absent.
130. Basisphenoid, basipterygoid processes, length: elongate (1); truncate (2). Noncomparable where absent.
131. Basisphenoid, basipterygoid processes, facet for articulation with pterygoids, shape: ovoid (1); acuminate (2). Noncomparable where absent. Musser and Cracraft [
44], character 93; Mayr and Clarke [
63], character 24.
132. Basiparasphenoid plate is inflated, rounded and broad: absent (0); present (1). Mayr and Clarke [
63], character 26; Cracraft and Clarke [
115]; Cracraft [
117].
133. Basiparasphenoid plate, well-marked depression on posterolateral side of basiosphenoid plate containing numerous foramina for arteries and cranial nerves: absent (0); present (1). Mayr and Clarke [
63], character 26; Cracraft and Clarke [
115]; Cracraft [
117].
134. Quadratopterygoid juncture: articulatio complex, involving both broad contact on facies medialis of processus orbitalis supplementary to condylus pterygoideus quadraticum (0); articulatio duplex, moderate dorsal extension on medial face of orbital process combined with condylus pterygoideus (1); articulatio simplex, retaining vestigial contact on basis of processus orbitalis in addition to primary articulatio with condylus pterygoideus (2); articulatio simplex, virtually limited to condylus pterygoideus (3). Musser and Cracraft [
44], character 65; Livezey and Zusi [
95], character 600; Mayr and Clarke [
63], character 26.
135. Temporal fossa: mostly only visible from lateral view, terminal end located ventrocranially to nuchal crest (1); edges of temporal fossae almost meet or do meet above nuchal crest (2). Musser and Cracraft [
44], character 66; Bertelli et al. [
14], character 19.
136. Temporal fossa, crest: shallow, at same level of top of skull (1); consists of a distinctive crest that is raised prominently (2). Musser and Cracraft [
44], character 67.
137. Zygomatic process, status: present (1); lost (2). Mayr and Clarke [
63], character 33.
138. Zygomatic process, length: truncate (1); elongate, reaches level of base of orbital process of quadrate (2). Noncomparable where absent. Musser and Cracraft [
44], character 68; Livezey and Zusi [
95], character 146.
139. Zygomatic process, ankylosed to postorbital process: no (1); yes (2). Noncomparable where absent.
140. Zygomatic process, divided: no (1); yes (2). Noncomparable where absent or if aponeurosis present. Musser and Cracraft [
44], character 69.
141. Zygomatic process, dorsal and ventral branches, relative rostrocaudal position if divided: dorsal branch more elongate cranially than ventral branch (1); subequal (2). Noncomparable where absent or if aponeurosis present.
142. Zygomatic process, articular notch for lateral head of quadrate: notch facing cranially (1); notch angled ventrally (2). Noncomparable in absence of the zygomatic process. Musser and Cracraft [
44], character 70.
143. Zygomatic process, ossifications (aponeurosis): absent (0); present (1). Noncomparable where absent.
144. Zygomatic process, rostrocaudal orientation: at level of postorbital process (1); significantly caudal to postorbital process (2). Noncomparable where absent.
145. Processus suprameaticus: absent as distinct processus, in most or all cases the homologous bone evidently continuous as rostral margin of meatus acusticus externus (0); present as variably prominent postorbital process (1). Musser and Cracraft [
44], character 71; Livezey and Zusi [
95], character 143.
146. Processus suprameaticus (if present), shape: subrectangular (1); acuminate (2). Noncomparable where absent.
147. Occipital, foramen dorsomediana, cf. foramen (ostium): absent, foramina bilaterally symmetrical within occipital region (0); present, distinctly dorsal, often proximate to transverse nuchal crest (1). Musser and Cracraft [
44], character 72; Livezey and Zusi [
95], character 77.
148. Occipital, transverse nuchal crest, dorsal portion: shallow or barely visible (1); distinct and prominent, raised dorsally (2). Musser and Cracraft [
44], character 73.
149. Occipital, fonticuli occipitalis: absent (0); present, large, ovoid and perforate (1). Musser and Cracraft [
94], character 74; Mayr and Clarke [
63], character 27; Livezey [
118], character 9; Ericson [
119], character 1.
150. Occipital, foramen v. occipitalis externae: present or very distinctly visible (1); absent or smoothed to the point of being hardly or not visible (2). Musser and Cracraft [
44], character 76.
151. Occipital, supraoccipital eminence: right and left portions of occipital complex indistinguishable or have very faint, smoothed division (1); occipital complex distinctly separated by median nuchal crest, appearance of a line or extremely bulbous process dividing the complex (2). Musser and Cracraft [
44], character 77.
152. Occipital, foramen magnum: round or dorsoventrally ovoid (1); lateromedially elongate (2). Musser and Cracraft [
44], character 78; Livezey and Zusi [
95], character 27.
153. Occipital, foramen n. abducentis: round (1); craniocaudally elongate (2). Musser and Cracraft [
44], character 79.
154. Occipital, occipital condyle, form: essentially circular (1); distinctly bilobate or reniform, lobes partitioned by medial condylar notch, lateromedially elongate (2); essentially round but flattened along ventral margin of foramen magnum (3). Musser and Cracraft [
44], character 80; Livezey and Zusi [
95], character 21.
155. Occipital, occipital condyle, rostrocaudal position relative to exoccipital, processus paroccipitalis: rostral (1); approximately equal or caudal (2). Musser and Cracraft [
44], character 81; Livezey and Zusi [
95], character 24.
156. Occipital, subcondylar fossa: deep (1); shallow (2); lost (3). Musser and Cracraft [
44], character 82.
157. Occipital, hypoglossal nerve foramen: circular (1); elongate (2). Musser and Cracraft [
44], character 83.
158. Occipital, regardless of supraoccipital eminence or exoccipital, rostrocaudal position relative to nuchal crest: caudal (1); subequal or rostral (2).
159. Occipital, dorsal apex of nuchal crest, dorsoventral position: subequal or ventral to dorsal base of postorbital process (1); dorsal to postorbital process (2).
160. Occipital, basioccipital and basiparasphenoid: medially compressed (1); laterally splayed (2). Musser and Cracraft [
44], character 86.
161. Occipital, fossa parabasalis: absent (0); present (1). Musser and Cracraft [
44], character 87; Livezey and Zusi [
95], character 120.
162. Occipital, fossa parabasalis (if present), depth: shallow (1); deep (2). Noncomparable where absent. Musser and Cracraft [
44], character 87; Livezey and Zusi [
95], character 120.
163. Occipital, basioccipital, crista basilaris transversa: absent or barely visible (0); present (1).
164. Basiparasphenoid, pneumatic foramen in center: absent (0); present (1). Musser and Cracraft [
44], character 92.
165. Occipital, basioccipital, concave: no, convex (1); yes (2).
166. Basiparasphenoid, processus medialis parasphenoidalis, alignment: lateral (1); ventral (2).
167. Basiparasphenoid, eustacian tubes fossa: deep (1); shallow (2). Noncomparable where basisphenoid covering homologous site. Musser and Cracraft [
44], character 94.
168. Pila otica, status: present (1) lost, only cotyla quadrati otici present so that recessus tympanicus completely exposed (2).
169. Squamosal, laterally splayed: no (1); yes (2).
170. Dorsally projecting process on quadrate anterior to quadrate-prootic articulation (attachment for m. adductor mandibulae externum, pars profundus): absent or very poorly developed (0); present and well developed (1).
171. Quadrate, otic process, lateral face: convex or flat (1); slightly concave (2); deeply concave (3). Musser and Cracraft [
44], character 96.
172. Quadrate, otic process, medial head, markedly extended caudally compared to lateral head: no, positions subequal (1); yes (2). Noncomparable where heads not well separated.
173. Quadrate, otic process, lateral head, rostrocaudally compressed: no (1); yes (2). Noncomparable where heads not well separated.
174. Quadrate, medial condyle: present (1); essentially absent, linear (2).
175. Quadrate, lateral capitulum, rostrocaudal position relative to quadratojugal articulation: quadratojugal articulation rostral (1); subequal (2); caudal to lateral head (3).
176. Quadrate, tuberculum subcapitulare under lateral capitulum, status: absent (0); present (1).
177. Quadrate, tuberculum subcapitulare (if present), prominence: prominent, tuberculate (1); diminutive, barely raised scar (2). Noncomparable where absent.
178. Quadrate, tuberculum subcapitulare (if present), ankylosis to zygomatic process: no (1); yes (2). Noncomparable where absent.
179. Quadrate, otic process, crista tympanica, prominence: absent or indiscernible (1); present, shallow (2); present, extremely prominent crista (3).
180. Quadrate, otic process, crista tympanica, ventral apex, dorsoventral location: within dorsal half of quadrate or at midpoint (1); within ventral half of quadrate (2). Not comparable where crista absent.
181. Quadrate, otic process, crista tympanica, mediolateral location relative to lateral capitulum: along midline of lateral capitulum (1); encompasses entire lateral capitulum (2). Not comparable where crista absent.
182. Quadrate, otic process, cotylae: cotylae confluent (1); cotylae completely separated (2). Musser and Cracraft [
44], characters 97–98; Livezey and Zusi [
95], characters 150 and 551; Mayr and Clarke [
63], character 34.
183. Quadrate, otic process, caudal face dorsal to crista tympanica (or homologous site), deeply concave: no (1); yes (2).
184. Quadrate, otic process (caudal aspect), mediolateral bowing toward ventral end: no, linear (1); yes, lateral bowing (2).
185. Quadrate, otic process, capitulum, medial face, pneumatic foramen: present (1); lost (2). Musser and Cracraft [
44], character 99; Livezey and Zusi [
95], character 508.
186. Quadrate, otic process, capitulum, medial face, pneumatic foramen (if present), location with respect to crista medialis and crista tympanica: rostral to crista medialis and crista tympanicum (1); rostral to crista tympanicum but caudal to crista medialis (2); like state 1 but located further ventrally (3). Noncomparable where foramen absent.
187. Quadrate, otic process, capitulum, medial face, pneumatic foramen, dorsoventral location: present within dorsal 2/3 (1); present within ventral third (2). Noncomparable where foramen absent.
188. Quadrate, otic process, capitulum, caudal face, pneumatic foramen, status: present (1); lost (2). Musser and Cracraft [
44], character 100; Livezey and Zusi [
95], character 554.
189. Quadrate, otic process, capitulum, caudal face, pneumatic foramen, shape: circular (1); elongate and ovoid (2). Noncomparable where foramen absent.
190. Quadrate, orbital process, medial face, depressio protractoris, form: flattened or only slightly concave (1); deeply concave (2). Noncomparable where orbital process absent.
191. Quadrate, orbital process, status: present (1); lost (2). Musser and Cracraft [
44], character 102; Livezey and Zusi [
95], character 532.
192. Quadrate, orbital process, length relative to otic process: subequal (1); shorter than otic process (2); longer than otic process (3). Noncomparable where orbital process absent. Musser and Cracraft [
44], character 103; Livezey and Zusi [
95], character 534.
193. Quadrate, orbital process, medially bowed: yes (1); no (2). Noncomparable where orbital process absent.
194. Quadrate, orbital process, crista orbitalis, prominence: shallow (1); prominent (2). Noncomparable where orbital process absent.
195. Quadrate, orbital process, shape of terminus: terminus subequal or wider than orbital process (1); exceptionally tapered toward terminus (2). Noncomparable where orbital process absent. Musser and Cracraft [
44], character 104–105; Bertelli et al. [
14], character 25; Livezey and Zusi [
95], character 535 and 538.
196. Quadrate, medial condyle, orientation: aligned cranio-caudally (1); aligned medio-laterally (2). Musser and Cracraft [
44], character 107.
197. Quadrate, medial condyle, shape: convex and bulbous, undivided (1); bilobate, 2 lobes approximately equal in size (2); bilobate, lateral lobe distinctly smaller than medial lobe (3). Musser and Cracraft [
44], character 108; Livezey and Zusi [
95], character 518.
198. Quadrate, mandibular process, facies articulais pterygoidea (ventral face in those taxa having two): facies articularis, with slight anteromedial eminentia on basis (1); condylar, tubercular, or jugosublinear (2). Musser and Cracraft [
44], character 109; Livezey and Zusi [
95], character 523.
199. Quadrate, caudal condyle and mandibular articulation: present (1); lost (2).
200. Quadrate, caudal condyle, shape (caudal aspect): dorsoventrally compressed (1); bulbous (2). Noncomparable where caudal condyle absent.
201. Quadrate, lateral condyle, location relative to articulation with quadratojugal: caudal (1); rostral (2); subequal (3).
202. Quadrate, lateral condyle, relatively large and oriented lateromedially: absent (0); present (1).
203. Quadrate, lateral condyle, size compared to caudal condyle: smaller than caudal condyle (1); larger than caudal condyle (2); subequal (3). Noncomparable where caudal condyle absent.
204. Quadrate, caudal condyle, orientation (caudal aspect): medial margin ventral to lateral margin (1); horizontal (2); lateral margin ventral to medial margin (3). Noncomparable where caudal condyle absent.
205. Quadrate, caudal condyle, confluence with lateral condyle: present (1); lost (2). Noncomparable where caudal condyle absent.
206. Quadrate, lateral condyle, dorsoventral position relative to caudal condyle regardless of confluence: caudal condyle positioned only slightly dorsal to lateral condyle (1); lateral condyle terminates well ventral to caudal condyle (2). Noncomparable where caudal condyle absent.
207. Quadrate, facies articularis quadratojugalis: fovea and cotyla (1); incisura—concave, troughlike, raised margin lacking entirely or at least in two, geometrically opposing points (2). Musser and Cracraft [
44], character 114; Livezey and Zusi [
95], character 511.
208. Quadrate, lateral margin of processus lateralis dorsal to cotyla quadratojugalis, prominent tubercle present that projects rostrolaterally past cotyla quadratojugalis: absent (0); present (1). Noncomparable where cotyla/fovea absent. Musser and Cracraft [
44], character 40.
209. Quadrate, caudal condyle, medial to medial head of quadrate: no (1); yes (2). Noncomparable where cotyla/fovea absent.
210. Quadrate, facies articularis quadratojugalis, cotyla quadratojugalis, orientation: oriented laterally (1); oriented rostrally (2); oriented dorsally (3). Noncomparable where cotyla/fovea absent.
211. Quadrate, facies articularis quadratojugalis, cotyla quadratojugalis, exceptional dorsoventral thickening of the rostroventral margin of cotyla: absent (0); present (1). Noncomparable where cotyla/fovea absent.
212. Quadrate, lateral aspect, cotyla quadratojugalis and processus lateralis, caudodorsally oriented, elongate flange that is separate from lateral and caudal condyles: absent (0); present (1).
Skull: Mandible
213. Articular surface, having single centrally located ridge oriented rostrocaudally: absent (0); present (1).
214. Articular surface, posteromedial and lateral walls: absent (0); present (1).
215. Symphysis, lamina ventral to foramina neurovascularis, craniocaudal alignment with dorsal lamina of symphysis: subequal, dorsal and ventral lamina craniocaudally aligned (1); ventral lamina extends caudal to dorsal lamina (2). Musser and Cracraft [
44], character 115; Livezey and Zusi [
95], character 654.
216. Symphysis, length as proportion of total length of mandible (not including retroarticular process if present): short, less than 1/5 (1); intermediate, between 1/5 and 1/3 (2); long, between 1/3 and 1/2 (3). Musser and Cracraft [
44], character 116; Livezey and Zusi [
95], character 676.
217. Ramus, general form: medially compressed (1); laterally splayed so that ramus on each side is mediolaterally oriented (2). Musser and Cracraft [
44], character 117; Livezey and Zusi [
95], character 280.
218. Symphysis, orientation: straight or dorsally oriented (1); apex ventrally recurved (2).
219. Ramus, portion rostral to coronoid process, curvature: present, variably but distinctly decurved (1); obsolete, i.e., virtually straight (2). Musser and Cracraft [
44], character 119; Livezey and Zusi [
95], character 673.
220. Coronoid process, shape: rounded (1); subrectangular (2).
221. Tuberculum m. adductor mandibulae externus, pars articularis, caput externus (see tuberc. m. AME in Livezey and Zusi 2006), laterally extensive: absent (0); present (1).
222. Ventral angle: absent or indistinct (0); present, marked (1). Musser and Cracraft [
44], character 120; Livezey and Zusi [
95], character 681.
223. Fenestra caudalis mandibulae, size: small, essentially absent (1); large and perforate (2).
224. Fenestra rostralis mandibulae, size: essentially absent, thin and slit-like (0); substantial and transverse, completely or largely perforate (1). Musser and Cracraft [
44], character 121; Livezey and Zusi [
95], character 684.
225. Surangular, dorsoventrally thick: no (1); yes (2).
226. Surangular and dentary, dorsoventrally thicker than articular area/angular: no (1); yes (2).
227. Angulus mandibulae, dorsally prominent: no (1); yes (2). Noncomparable for Anseriformes.
228. Prearticular: extends to rostral margin of fenestra rostralis mandibulae (1); only extends to caudal margin of fenestra rostralis mandibulae (2).
229. Area of articulation with quadrate, lateral cotyle, mediolaterally compressed: no (1); yes (2).
230. Retroarticular process (if present), dorsoventral thickening: absent, processes thin and elongate (1); present, processes dorsoventrally thick and mediolaterally flattened (2).
231. Dentary, caudal development: strongly forked (1); weakly forked posteriorly into dorsal and ventral rami (2). Musser and Cracraft [
44], character 123; Cracraft and Clarke [
115], character 4.
232. Area of articulation with quadrate, lateral cotyle, orientation of rostral margin compared to that of caudal margin: oriented laterally (1); oriented medially (2). Musser and Cracraft [
44], character 124.
233. Area of articulation with quadrate, internal articular process, pneumatic foramen: absent (0); present (1).
234. Area of articulation with quadrate, fossa articularis quadratica, sulcus intercotylaris, pneumatic foramina: absent (0); present (1). Musser and Cracraft [
44], character 126; Livezey and Zusi [
95], character 696.
235. Area of articulation with quadrate, cotylae fossae articularis, tuberculum intercotylare: variably tuberculate with intervening depressions (1); single, centrally positioned, rostrocaudally oriented jugum (2).
236. Area of articulation with quadrate, internal articular process: truncate (1); elongate (2).
237. Area of articulation with quadrate, processus lateralis mandibulae, size: small, diminutive (1); prominent (2). Note: not to be confused with cotyla lateralis, it is located rostral to this feature.
238. Area of articulation with quadrate, dorsally projecting hook-like accessory projection present caudolateral to cotyla caudalis: absent (0); present, dorsoventrally elongate and hook-like (1); present but truncate and rounded, sometimes present as a pedestal-like projection with a flattened dorsal surface (2). Noncomparable where articulation for caudal condyle of quadrate is absent in mandible. Note: This character is not to be confused with a true retroarticular process seen in e.g., galloanserines and Phoenicopteridae. The difference is that while the hook-like process seen in Neoaves (e.g., Gruiformes) is only present caudolateral to the caudal condyle (between area of articulation of caudal and lateral condyles of quadrate), the true retroarticular process is medial to or within the margin of the caudal cotyla or medial to the cotyla lateralis where the articulation for the caudal condyle is absent. The true retroarticular process must incorporate medial lamina of the mandible. Some Sphenisciformes appear to have a true retroarticular process, but that is actually the area of the caudal fossa of the mandible that has been elevated and flattened dorsally. Musser and Cracraft [
44], character 129; Bertelli et al. [
14], character 26.
239. Dorsally projecting hook-like accessory projection present caudolateral to cotyla caudalis (if present), extreme caudal elongation of all dorsoventral lamina: absent or only slight caudal projection present (0); present, creates mediolaterally flattened “pseudo processus retroarticularius” (1). See note above. Noncomparable where absent.
240. True retroarticular process: absent (0); present (1).
241. True retroarticular process (if present), medial margin and impressio m. depressor mandibulae, hollow: no (1); yes (2). Noncomparable where absent.
242. Area of articulation with quadrate, extreme dorsoventral flattening: not present, area dorsoventrally robust (1); present (2).
243. Area of fossa caudalis/impressio m. depressor mandibulae, ventral apex, rostrocaudal position: apex subequal in rostrocaudal position to caudal margin of area of articulation with quadrate (1); rostral, apex rostral to caudal margin of area of articulation with quadrate (2); caudal, apex extended caudally so that caudal fossa visible in dorsal aspect (3).
244. Fossa caudalis, status and depth: absent (0); present, shallow (1); present, deep (2). Noncomparable where hollow.
Vertebrae
245. Number of presacral vertebrae: 20–22 (1); 18–19 (2); 23 or more (3). Mayr and Clarke [
63], character 55.
Vertebrae: Cervical Vertebrae
246. Atlas, processus ventralis corporis, location of caudal apex: caudal (1); ventrocaudal (2).
247. Atlas, fossa condyloidea, shape: a complete, circular facet (1); lateromedially elongate with a subrectangular facet, does not form circle (2).
248. Axis, spinous process, terminus caudal to corpus: no (1); yes (2).
249. Axis, ventral process: essentially absent (0); present, represented by variably thick, rounded or subangular crista (1); present, represented by a ventrally elongated, caudally deflected, rounded spina (2); present, represented by a ventrally elongated, bilaterally compressed, craniocaudally restricted lamina (3). Musser and Cracraft [
44], character 133; Livezey and Zusi [
95], character 783.
250. Axis, processus costalis: present, typically vestigial, i.e., represented only by head (1); lost (2). Musser and Cracraft [
44], character 134; Livezey and Zusi [
95], character 787; Mayr and Clarke [
63], character 50.
251. Axis, foramen transversarium: absent (0); present (1). Note: related in part to prominence of processus costalis. Musser and Cracraft [
44], character 135; Mayr and Clarke [
63], character 49; Livezey [
113], character 104.
252. Axis, lateral lamina of arcus, pneumatic foramen: absent (0); present (1). Musser and Cracraft [
44], character 136; Mayr and Clarke [
63], character 48; Livezey [
113], character 107.
253. Cervical vertebrae, marked heterogeneity of form involving relative elongation of intermediate elements (regardless of length): present (1); lost (2). Musser and Cracraft [
44], character 137; Livezey [
113], character 111.
254. Cervical vertebrae, corpus, dorsoventrally compressed: no (1); yes (2).
255. Cervical vertebrae, corpus, extremely craniocaudally elongate: no (1); yes (2).
256. Cervical vertebrae, section I, arcus interzygopophysialis lateralis: absent on all elements (0); present (1). Musser and Cracraft [
44], character 138; Livezey [
113], character 806.
257. Cervical vertebrae, section I, arcus vertebrae, spinous processes, craniocaudal location of apices: close to cranial margin or midpoint of vertebrae (1); reaching caudal margin of vertebrae (2).
258. Cervical vertebrae, section I, arcus vertebrae, spinous processes, shape: subrectangular (1); rounded (2); bifurcated (3).
259. Cervical vertebrae, section I, arcus vertebrae, spinous processes, dorsal lengths: dorsoventrally truncate (1); dorsoventrally elongate (2).
260. Cervical vertebrae, section II, arcus vertebrae, spinous processes, craniocaudal locations of apices: close to cranial margin of vertebrae (1); at midpoint of vertebrae (2).
261. Cervical vertebrae, section II, zygapophysis caudalis, dorsal portion (torus dorsalis): convex (1); concave (2).
262. Cervical vertebrae, section II, zygapophysis caudalis: craniocaudally elongate (1); craniocaudally truncate (2).
263. Cervical vertebrae, section II, costal processes: present throughout section II (1); present in cranial elements but absent in caudal elements of section II (2); absent throughout section II (3). Musser and Cracraft [
44], character 139; Livezey [
113], character 820.
264. Cervical vertebrae, section II, ansa costotransversaria: as craniocaudally elongate as at approximately half of vertebra (1); craniocaudally short so that ansa is as elongate as 1/3 of vertebra or less (2); like state 1 but toward cranial elements can be essentially as craniocaudally elongate as entire corpus (3).
265. Cervical vertebrae, section II, costal processes (if present), ansa costotransversaria, foramen: absent (0); present (1).
Vertebrae: Thoracic Vertebrae
266. Thoracic vertebrae, corpus, lateral face, pneumatic foramen: absent (0); present (1). Musser and Cracraft [
44], character 143; Livezey and Zusi [
95], character 850.
267. Thoracic vertebrae, corpus, bilateral compression manifested by virtually laminar structure of corpus between facies articulares craniali et caudalis: no, corpus cylindrical (1); present (2). Musser and Cracraft [
44], character 144; Livezey and Zusi [
95], character 858.
268. Thoracic vertebrae, dorsal margins of spinous processes, overlapping craniocaudally (excluding those within notarium, if present): no (1); yes (2).
269. Thoracic vertebrae, dorsal surface and spinous processes, extensive aponeurosis ossificans: no (1); yes (2).
270. Thoracic vertebrae, dorsal lamina of arcus, recessus dorsocranialis pneumatici: absent (0); present (1). Musser and Cracraft [
44], character 145; Livezey and Zusi [
95], character 866.
271. Thoracic vertebrae, dorsal lamina of arcus, fovea interzygopophysialis, pneumatic foramina: absent (0); present (1). Musser and Cracraft [
44], character 146; Livezey and Zusi [
95], character 867.
272. Thoracic vertebrae, transverse processes, craniocaudal gradual but distinct lateral elongation of processes in seratium among presynsacral elements: absent, transverse processes essentially of uniform width throughout the presynsacral elements or moderately reversed grade (0); present, transverse processes distinctly increasing in width throughout the presynsacral elements (1). Musser and Cracraft [
44], character 147; Livezey and Zusi [
95], character 875.
273. Thoracic and sacral vertebrae, caudalmost elements (cranial to synsacral vertebrae), facies articularis caudalis (of penultimate element) and facies articularis cranialis (of ultimate element), type: series completely heterocoelous (1); at least part of series with subround, central articular surfaces (e.g., amphicoelous/opisthocoelous) that lack the dorsoventral compression and saddle-shaped articular surface seen in heterocoelous vertebrae (2). Musser and Cracraft [
44], character 148; Livezey and Zusi [
95], character 888; Mayr and Clarke [
63], character 57.
274. Thoracic vertebrae, notarium: present (1); lost (2). Musser and Cracraft [
44], character 151; Livezey and Zusi [
95], character 892; Mayr and Clarke [
63], character 55.
275. Thoracic vertebrae, notarium (if present), number of incorporated vertebrae: 4 (1); 3 (2). Noncomparable where absent. Musser and Cracraft [
44], character 151; Livezey and Zusi [
95], character 892.
276. Thoracic vertebrae, notarium (if present), corpus notarii, crista ventralis notarii, fenestrae intercristales: typically present, ligamenta intercristales of adjacent vertebrae notarii (often robustly) ossified (1); typically lost, cristae ventrales demarcated ventrally by incisurae cristae, ligamenta intercristales of adjacent vertebrae notarii not ossified (2). Not comparable for taxa without a notarium. Musser and Cracraft [
44], character 152; Livezey and Zusi [
95], character 895.
277. Thoracic vertebrae, notarium (if present), arcus notarii, foramina (intervertebralia) interarcuales: present, small, diameter approximately equal to that of fovea costalis (1); present, obsolete, reduced to minute perforations or evidently absent (2). Not comparable for taxa without a notarium. Musser and Cracraft [
44], character 153; Livezey and Zusi [
95], character 896.
278. Thoracic vertebrae, ventral processes, widely bifurcated (lateral termini extending laterally beyond costal facets): no (1); yes (2).
279. Caudalmost presacral vertebrae with deep lateral excavations: no (1); yes (2). Mayr and Clarke [
63], character 58.
Vertebrae: Synsacrum and Caudal Vertebrae
280. Synsacrum (maximally including thoracic vertebrae): 14–18 (1); 10–13 (2). Musser and Cracraft [
44], character 154; Livezey and Zusi [
95], character 897; Mayr and Clarke [
63], character 91.
281. Synsacrum, cranialmost vertebrae caudales synsacri, lateral faces of vertebrae, penetration and visibility through foramen ilioischiadicum (lateral perspective): absent (0); present (1). Musser and Cracraft [
44], character 158; Livezey and Zusi [
95], character 933.
282. Free caudal vertebrae, number (including pygostyle): 5–6 (1); 7–8 (2); 9–10 (3).
283. Pygostyle, fused to last free caudal via spinous process of caudal vertebra(e): no (1); yes (2).
284. Pygostyle, fused to last free caudal(s) via ventral processes: no (1); yes (2).
285. Pygostyle, foramen in lateral lamina: absent (0); present (1).
286. Free caudal vertebrae, spinous processes, mediolaterally wide bifurcation throughout series, status: present (1); lost (2).
287. Free caudal vertebrae, ventral processes, status: present (1); lost (2).
288. Free caudal vertebrae, ventral processes (if present), general orientation of apex: cranioventral (1); cranial (2).
289. Free caudal vertebrae, spinous process, form: rounded (1); subrectangular (2).
290. Free caudal vertebrae, transverse processes: truncate (1); elongate (2).
291. Pygostyle, apex, shape: subrectangular, tapered (1); rounded (2); hamate (3); spatulate (4).
292. Pygostyle, apex, dorsoventral departure from major axis of pygostyle: absent (0); dorsal (1); ventral (2). Noncomparable if spatulate.
293. Pygostyle, craniocaudal length: elongate (1); truncate (2).
Sternum
294. Spina externa rostri: absent or obsolete (0); present (1). Musser and Cracraft [
44], character 159; Livezey and Zusi [
95], character 1157; Mayr and Clarke [
63], character 70.
295. Spina externa rostri, fused to furcula: no (1); yes (2).
296. Spina externa rostri, length: truncate, shorter than craniolateral processes (1); elongate, longer than craniolateral processes (2). Noncomparable where spina externa absent. Musser and Cracraft [
44], character 159; Livezey and Zusi [
95], character 1157; Mayr and Clarke [
63], character 70.
297. Spina externa rostri, shape: fan-shaped (1); spine-like (2). Mayr and Clarke [
63], character 70.
298. Spina externa rostri, completely bifurcated: no (1); yes (2).
299. Spina interna rostri: present (1); lost (2). Bertelli et al. [
14], character 45.
300. Spina interna rostri, fused to spina externa rostri: yes (1); no (2). Noncomparable where spina interna and spina externa absent. Bertelli et al. [
14], character 45.
301. Spina interna rostri (if present), pneumatic foramen: absent (0); present (1). Noncomparable where spina interna absent.
302. Spina externa rostri (if present), pneumatic foramen: absent (0); present (1). Noncomparable where spina externa absent.
303. Spina externa rostri (if present), crest along ventral midline: absent (0); present (1). Noncomparable where spina externa absent.
304. Spina externa rostri (if present), invagination of dorsal margin: present (1); lost (2).
305. Spina externa rostri (if present), exceptionally mediolaterally wide: no (1); yes (2).
306. Spina externa rostri (if present), lateral faces deeply concave: no (1); yes (2).
307. Depressiones (sulcus) articulares coracoidei, dorsal lip: reaches dorsally (1); reaches ventrally (2). Musser and Cracraft [
44], character 160.
308. Depressiones (sulcus) articulares coracoidei, ventral lip: reaches ventrally (1); reaches dorsally (2). Musser and Cracraft [
44], character 162.
309. Depressiones (sulcus) articulares coracoidei, crossed: no (1); yes (2). Mayr and Clarke [
63], character 69; Ericson [
119], character 34.
310. Processus craniolateralis: extremely long, creates fenestra (1); prominent but more truncate than state 1 (2); extremely truncate, barely projected (3). Musser and Cracraft [
44], character 164; Livezey and Zusi [
95], character 1141.
311. Processus craniolateralis with respect to major axis of carina, angle: extending laterally (1); extending rostrally (2). Musser and Cracraft [
44], character 165; Livezey and Zusi [
95], character 1142.
312. Processus craniolateralis, apex, shape: trapezoidal (1); rounded (2); cruciate (3).
313. Processus craniolateralis, apex, width compared to width of base: subequal (1); apex narrower than base (2).
314. Ventral face of sternum, base of processi craniolateralis and/or processus craniolateralis proprius, impressio origii m. sternocoracoidei: present (1); lost or extremely shallow (2). Musser and Cracraft [
44], character 166; Livezey and Zusi [
95], character 1149.
315. Visceral face of sternum, sulcus medianus sterni immediately caudal to cranial margin, pneumatic foramen and/or depression exclusive of pneumatic pores: absent (0); present, undivided, enclosing pneumatic pores and os spongiosum (1); present, divided medially by osseus lamina or trabecula, enclosing pneumatic pores and os spongiosum (2). Musser and Cracraft [
44], character 167; Livezey and Zusi [
95], character 1108.
316. Visceral face of sternum, sulcus medianus sterni and/or pneumatic foramen: begins within margin of or is immediately caudal to pila coracoidea (1); significantly caudal to pila coracoidea, approximately caudal to base of carina (2). Musser and Cracraft [
44], character 168; Livezey and Zusi [
95], character 1109.
317. Visceral face of sternum, pneumatic pores, exclusive of those included within pneumatic foramen: absent (0); present, at cranial margin (1); present, along medial sulcus (2); present, within both cranial margin and medial sulcus (3). Musser and Cracraft [
44], character 169; Livezey and Zusi [
95], character 1110.
318. Ventral face, facies muscularis, sulcus ventrolateralis (longitudinal trough on ventral surface of element immediately medial to processus costales): absent or indistinct (1); distinct, typically for length of costal margin (2). Musser and Cracraft [
44], character 171; Livezey [
113], character 153.
319. Costal margin, craniocaudal length relative to that of entire sternum along median axis: less than ¼ (1); between 1/4 and ¾ (2); greater than ¾ (3). Musser and Cracraft [
44], character 172; Livezey [
113], character 1114.
320. Costal margin, articular surfaces for sternal ribs: 4 or less (1); 5–7 (2); 8–9 (3). Musser and Cracraft [
44], character 173; Mayr and Clarke [
63], character 71.
321. Costal margin, articular surfaces for sternal ribs, location: on processus craniolateralis and body (1); on body only (2); on processus craniolateralis only (3).
322. Costal margin, linea intermuscularis dorsolateralis: absent (0); present (1).
323. Costal margin, linea intermuscularis dorsolateralis, mediolateral location of caudal terminus: medial only to trabecula caudolateralis (if present) (1); medial to trabecula cauolateralis and intermediana (if present) but lateral to mediana (2); medial to lateral margin of trabecula mediana (3). Noncomparable where linea intermuscularis dorsolateralis absent.
324. Costal margin, linea intermuscularis dorsolateralis, caudal extent: within rostral half of sternum only (1); extends to caudal margin of sternum or well within caudal half of sternum (2). Noncomparable where linea intermuscularis dorsolateralis absent.
325. Carina, cranial margin, lateral crest: present (1); lost (2). Noncomparable for taxa without a carina. Musser and Cracraft [
44], character 176; Livezey and Zusi [
95], character 1207.
326. Carina, cranial margin, sulcus carinae: absent (0); present (1). Not comparable for taxa without a carina. Musser and Cracraft [
44], character 177; Livezey and Zusi [
95], character 1213.
327. Carina, sulcus carinae or homologous site, pneumatic foramen: absent (0); present, variable size and shape (1). Not comparable for taxa without a carina. Musser and Cracraft [
44], character 178; Livezey and Zusi [
95], character 1218.
328. Carina, sulcus carinae or homologous site, crista complete along midline: present (1); lost (2). Noncomparable where carina absent.
329. Carina, apex, site relative to spina externa rostri or most proximal point of sternum: subequal to caudal location of spina externa rostri or homologous site (1); located caudally (2); located extremely cranially (3). Noncomparable where carina absent. Musser and Cracraft [
44], character 179; Livezey and Zusi [
95], character 1198.
330. Carina, apex, fused to furcula: no (1); yes (2). Noncomparable where carina absent. Musser and Cracraft [
44], character 180; Livezey and Zusi [
95], character 1200.
331. Carina, apex, facet for articulation with furcula: absent (0); present, as incisura or concavity (1). Noncomparable where carina absent. Musser and Cracraft [
44], character 180; Livezey and Zusi [
95], character 1200.
332. Carina, cranial margin, relative widths of dorsal and ventral portions: dorsal margin wider (1); ventral margin wider (2); width of both portions subequal (3). Noncomparable where carina absent. Musser and Cracraft [
44], character 181.
333. Carina, maximal depth ventral and normal to body of sternum, facies muscularis, relative to minimal width of body of sternum (exclusive of processes laterales, if present) across points on costal margin directly lateral to that of maximal depth of carina: height is equal to or greater than width of body, not including processus craniolateralis (1); height is less than width of body (2). Noncomparable where carina absent. Musser and Cracraft [
44], character 182; Livezey and Zusi [
95], character 1199.
334. Carina, apex, shape: rounded (1); acuminate (2); trapezoidal (3); bifurcated (4). Noncomparable where carina absent.
335. Carina, recurvature of cranial margin: absent (0); present (1). Noncomparable where carina absent.
336. Carina, apex and ventral keel, reinforced with thickened jugum: no (1); yes (2). Noncomparable where carina absent.
337. Carina, linea intermuscularis ventromedialis, status: present (1); lost (2). Noncomparable where carina absent.
338. Carina, lateral aspect, exceptionally concave: no (1); yes (2). Noncomparable where carina absent.
339. Carina, caudal margin, continues to caudal margin of trabecula mediana: yes (1); no (2). Noncomparable where carina absent.
340. Sternum, caudal margin: with 4 or more notches/fenestrae (1); with 2 notches/fenestrae (2); without notches/fenestrae (3); with 3 notches/fenestrae (4). Musser and Cracraft [
44], character 183; Livezey and Zusi [
95], character 1182; Mayr and Clarke [
63], character 73.
341. Incisura and fenestra caudolateralis (if present), cranial extent (length of fenestra measured using body of sternum, not trabecula lateralis): elongate—length of incisura and fenestra greater than 2/3 craniocaudal length of corpus sterni, approaching terminis caudalis of processus costales sterni (1); intermediate—length of incisura and fenestra between 1/3 and 2/3 of craniocaudal length of corpus sterni (2); abbreviate—length of incisura and fenestra less than 1/3 craniocaudal length of corpus sterni (3). Noncomparable where incisura (fenestra) and/or trabecula absent. Musser and Cracraft [
44], character 184; Livezey and Zusi [
95], character 1183.
342. Processus caudolateralis (if present), orientation relative to body of sternum as reflected (in part) by angle defined by incisura caudolateralis, cranial vertex of angle: angle undefined, processus is parallel to costal margin and “vertex” is ellipsoidal (1); laterally splayed, approximately 45 degrees (2); angle acute, less than 15 degrees (3). Noncomparable where incisura (fenestra) and/or trabecula absent. Musser and Cracraft [
44], character 185; Livezey and Zusi [
95], character 1184.
343. Processus caudolateralis (if present), space between process and body of sternum: typical, wide (1); extremely narrow so that almost no space is present between sternum body and trabecula caudolateralis (2). Noncomparable where incisura (fenestra) and/or trabecula absent.
344. Trabecula lateralis, status: absent (0); present (1).
345. Trabecula caudolateralis (if present), shape of caudal terminal margin: rounded or (sub)rectangular (1); (sub)acuminate (2); cruciate, with transverse pila (3). Noncomparable where presence of trabecula uncertain or known to be absent. Musser and Cracraft [
44], character 186; Livezey and Zusi [
95], character 1185.
346. Trabecula caudolateralis (if present), orientation of caudal terminal margin: caudal, does not deviate from trabecula lateralis axis (1); medial (2); lateral (3). Noncomparable where presence of trabecula uncertain or known to be absent.
347. Trabecula mediana, thickened, raised and bifurcated crista that connects to the ventral margin of the carina and extends to the caudolateral margin of trabecula mediana: absent (0); present (1). Noncomparable where carina absent.
348. Dorsal lip, labrum interna sterni, pair of rostrally oriented projections: absent (0); present (1). Musser and Cracraft [
44], character 161.
349. Dorsal lip, labrum interna sterni, pair of rostrally oriented projections (if present), shape: rounded (1); acuminate (2). Noncomparable where processes absent. Musser and Cracraft [
44], character 187.
350. Dorsal lip, labrum interna sterni, pair of rostrally oriented projections (if present), raised dorsally with accompanying raised jugae along visceral margin of sternum: no (1); yes (2). Noncomparable where processes absent.
351. Dorsal lip, labrum interna sterni, pair of rostrally oriented projections (if present and raised dorsally), sulcus present along midline: absent (0); present (1). Noncomparable if not raised along visceral face.
352. Dorsal lip, labrum interna sterni, pair of rostrally oriented projections (if present), mediolateral position: gap present between 2 processes (1); 2 processes contact each other (2). Noncomparable where processes absent.
353. Dorsal lip, labrum interna sterni, pair of rostrally oriented projections (if present), mediolateral width: wide (1); narrow (2). Noncomparable where processes absent.
354. Incisura and fenestra intermediana, relative cranial extent: rudimentary, incisura is a minor concavity (1); moderately elongate, length of incisura and fenestra less than 1/3 craniocaudal length of corpus sterni (2). Noncomparable where incisura (fenestra) and/or trabecula absent. Musser and Cracraft [
44], character 188; Livezey and Zusi [
95], character 1188.
355. Trabecula mediana, margin and caudal terminus, tapering: untapered or weakly tapered (1); distinctly tapered (2).
356. Trabecula mediana, caudal terminus, shape: subrectangular (1); rounded or acuminate (2); cruciate with rounded transverse pila extending laterally beyond mediana (3); cruciate with elongate, acuminate transverse pila extending laterally beyond mediana (4). Musser and Cracraft [
44], character 189; Livezey and Zusi [
95], character 1191.
357. Trabecula mediana or intermediana, fused to trabecula lateralis: no (1); yes (2). Noncomparable where trabeculae absent.
358. Trabecula mediana, fused to trabecula intermediana: no (1); yes (2). Noncomparable where trabeculae absent.
359. Trabecula mediana, mediolateral width: mediolaterally narrow, essentially confined to midline of sternum (1); mediolaterally wide (2).
360. Trabecula caudolateralis and mediana, relative caudal extents: mediana > caudolateralis (1); caudolateralis > mediana (2). Noncomparable if no notches or fenestrae present. Musser and Cracraft [
44], character 190; Livezey and Zusi [
95], character 1192.
361. Trabecula intermediana, relative caudal extent: subequal in length to mediana (1); subequal in length to caudolateralis (2); between lengths of craniolateralis and mediana (3). Noncomparable if trabecula intermediana absent. Musser and Cracraft [
44], character 190; Livezey and Zusi [
95], character 1192.
362. Trabecula intermediana, shape of terminus: round or subrectangular (1); acuminate (2); cruciate (3). Noncomparable if trabecula intermediana absent.
363. Trabecula intermediana, orientation of caudal terminal end: caudal, does not deviate from trabecula intermediana axis (1); medial (2); lateral (3). Noncomparable if trabecula intermediana absent.
364. Trabecula intermediana, orientation relative to body of sternum as reflected (in part) by angle defined by incisura caudolateralis, cranial vertex of angle: angle undefined, processus is parallel to trabecula mediana and “vertex’’ is ellipsoidal (1); laterally splayed, approximately 45 degrees (2). Noncomparable if trabecula intermediana absent.
365. Sternum, tracheal convolution: absent (0); present (1).
Ribs
366. Thoracic ribs, medial face, pneumatic foramina between capitulae: present (1); lost (2). Musser and Cracraft [
44], character 191; Livezey [
113], character 132.
367. Thoracic ribs, dorsal portion, craniocaudally thickened: yes (1); no (2).
368. Uncinate processes on thoracic ribs: absent (0); present (1).
369. Uncinate processes on thoracic ribs, fusion to ribs: fused (1); unfused (2). Noncomparable where absent.
370. Uncinate processes on thoracic ribs, shape of terminus: subrectangular (1); rounded (2). Noncomparable where absent.
371. Uncinate processes on thoracic ribs, angle of articulation: oriented caudally (1); oriented dorsocaudally (2). Noncomparable where absent.
372. Uncinate processes, angulus basalaris processi: present (1); absent on all processes (2). Noncomparable where absent.
373. Uncinate processes on thoracic ribs, length: truncate (1); elongate (2). Noncomparable where absent.
Shoulder girdle: Coracoid
374. Coracoid, recessus infra-acrocoracoideus: shallow (1); extremely deep (2).
375. Coracoid, insertion of ligamentum acrocoracoprocoracoideum strongly ventromedially protruding: no, essentially coplanar with craniocaudal axis of shaft of coracoid (1); yes (2). Musser and Cracraft [
44], character 193; Livezey and Zusi [
95], character 1267.
376. Coracoid, facies articularis clavicularis overhanging sulcus supracoracoideus or homologous site: no (1); yes (2). Mayr and Clarke [
63], character 64.
377. Coracoid, facies articularis clavicularis, if overhanging sulcus supracoracoideus, exceptionally mediocaudally elongate and hamate (ventral aspect): no (1); yes (2). Noncomparable where not overhanging.
378. Coracoid, facies articularis clavicularis, if overhanging sulcus supracoracoideus, (medial aspect): subhorizontal (1); diagonal, with most proximal portion being located along dorsal margin (2). Noncomparable where not overhanging.
379. Coracoid processus acrocoracoideus, impressio ligamenti acrocoracohumeralis: deep (1); shallow (2). Musser and Cracraft [
44], character 195; Livezey and Zusi [
95], character 1276.
380. Omalis of coracoid, fused to furcula: no (1); yes (2).
381. Processus procoracoideus: present (1); lost (2). Musser and Cracraft [
44], character 197; Livezey and Zusi [
95], character 1283.
382. Processus procoracoideus, medial length: truncate (1); elongate (2). Noncomparable where processus procoracoideus absent. Musser and Cracraft [
44], character 197; Livezey and Zusi [
95], character 1283.
383. Processus procoracoideus, fused to head of coracoid: no (1); yes (2).
384. Processus procoracoideus, ankyolosed to furcula: no (1); yes (2).
385. Processus procoracoideus, terminus, shape: no spatulation or expansion present (1); spatulation and expansion of terminus present (2); tapering present (3). Noncomparable where processus procoracoideus absent or fused to head of coracoid.
386. Processus procoracoideus, terminus, caudally projecting, acuminate flange: absent (0); present (1). Noncomparable where processus procoracoideus absent or fused to head of coracoid.
387. Processus procoracoideus, tuberculum apicalis procoracoidei, ventral curvature: absent or slight (1); present (2). Noncomparable where processus procoracoideus absent.
388. Processus procoracoideus, extending distally as a craniocaudally elongate, sharp, crest along midline of shaft (crista procoracoidei of Livezey [
113]): no (1); yes (2). Noncomparable where processus procoracoideus absent.
389. Processus procoracoideus, if extending as a craniocaudally elongate crista, additional foramen located cranially: no (1); yes (2). Noncomparable where processus procoracoideus absent or craniocaudally truncate.
390. Processus procoracoideus, if extending as a craniocaudally elongate crista, extreme medioventral projection of crista, even along distal margin: no (1); yes (2). Noncomparable where processus procoracoideus absent or craniocaudally truncate.
391. Scapular cotyla, form: shallow (1); deep, cuplike (2). Bertelli et al. [
14], character 37.
392. Scapular cotyla, large pneumatic foramen or foramina: absent (0); present (1).
393. Coracoid shaft, foramen nervi supracoracoidei: absent (0); present (1). Mayr and Clarke [
63], character 63.
394. Processus procoracoideus, pneumatic foramen directly below facies articularis scapularis which does not penetrate shaft: absent (0); present (1). Musser and Cracraft [
44], character 198; Livezey and Zusi [
95], character 1286; Mayr and Clarke [
63], character 66.
395. Coracoid shaft, general form sensu length relative to width of facies articularis sternalis (not including additional projections): elongate—length between 3 and 4× width (1); typically proportioned—length between 2 and 3× width (2); truncate (less than 2× width) (3). Musser and Cracraft [
44], character 199; Livezey and Zusi [
95], character 1292.
396. Sternal coracoid, impressio musculi sternocoracoidei on dorsal surface of extremitas sternalis: shallow (1); deep (2). Musser and Cracraft [
44], character 200; Livezey and Zusi [
95], character 1294; Mayr and Clarke [
63], character 67.
397. Sternal coracoid, impressio musculi sternocoracoidei, sulcus m. sternocoracoideus: absent, area smooth (0); present, defined ridges present (1).
398. Sternal coracoid, dorsal surface, impressio m. sternocoracoidei, pneumatic foramen, status: absent (0); present (1). Musser and Cracraft [
44], character 201; Mayr and Clarke [
63], character 67.
399. Sternal coracoid, crista articularis sternalis, cranial recurvature: absent (0); present (1).
400. Sternal coracoid, crista articularis sternalis, ventral recurvature: absent (0); present (1).
401. Sternal coracoid, medial angle: rounded (1); acuminate (2).
402. Sternal coracoid, medial angle, additional cranially oriented projection along cranial margin: absent (0); present (1). Noncomparable where crista extends along length of coracoid.
403. Sternal coracoid, medial angle, additional cranially oriented projection along cranial margin (if present), shape of apex: rounded (1); acuminate (2). Noncomparable where processus procoracoideus absent or craniocaudally truncate. Musser and Cracraft [
44], character 202; Livezey [
113], character 194.
404. Sternal coracoid, medial angle, additional cranially oriented projection along cranial margin (if present), craniocaudal length: craniocaudally truncate (1); craniocaudally elongate (2). Noncomparable where processus procoracoideus absent or craniocaudally truncate.
405. Sternal coracoid, processus lateralis, length: truncate (1); elongate (2).
406. Sternal coracoid, processus lateralis, shape of terminus: rounded or subrectangular (1); acuminate (2).
407. Sternal coracoid, processus lateralis, recurvature of cranial margin: absent or negligible (0); present (1).
408. Sternal coracoid, processus lateralis, recurvature of caudal margin: absent or negligible (0); present (1).
409. Sternal coracoid, processus lateralis, apex, orientation: lateral (1); rostral (2); caudal (3). Musser and Cracraft [
44], character 203; Livezey and Zusi [
95], character 1303.
410. Sternal coracoid, processus lateralis, apex, craniocaudal location: well cranial to lateral angle (1); just above lateral angle or subequal (2).
411. Sternal coracoid, lateral angle, mediolateral position relative to shaft of coracoid: medial to lateral margin of shaft (1); lateral to lateral margin of shaft (2).
412. Sternal coracoid, ventral aspect, distolateral corpus (just medial to lateral angle), deeply concave: no (1); yes (2).
413. Sternal portion of coracoid, ventral aspect, distomedial corpus (just lateral to medial angle), deeply concave: no (1); yes (2).
414. Sternal coracoid, facies articularis sternalis, labrum externa, general cranial extent of cranial margin (and correlated cranial expanse) relative to those of labrum interna: former approximately equal to latter, producing facies articularis of dorsoventrally equal expanse (1); former distinctly caudal to latter, producing internally (dorsally) angled facies articularis (2); former significantly cranial to the latter, producing externally (ventrally) angled facies articularis (3).
415. Sternal coracoid, facies articularis sternalis, labrum externa, labrum along medial angle ventrocranially angled: no (1); yes (2). Musser and Cracraft [
44], character 205; Livezey and Zusi [
95], character 1314.
Shoulder girdle: Furcula
416. General shape: laterally splayed, “u-shaped” (1); mediolaterally compressed, “v-shaped” (2).
417. Apophysis, status: absent (0); present (1).
418. Apophysis if present, caudal prominence: diminutive, barely raised (1); extremely elongate (2); intermediate (3). Noncomparable where apophysis absent.
419. Apophysis (if present), continues ventrally beyond symphysis and is dorsoventrally elongate: no (1); yes (2). Noncomparable where apophysis absent.
420. Apophysis (if present), mediolaterally wide: no (1); yes (2). Noncomparable where apophysis absent.
421. Processus interclavicularis dorsalis: absent (0); present (1).
422. Scapus clavicle, mediolateral width: extremely thin (1); intermediate (2); extremely thick (3).
423. Scapus clavicle, ventral curvature: absent (0); present (1).
424. Scapus clavicle, pneumatic foramen on lateral face: absent (0); present (1).
425. Scapus clavicle, craniocaudal length: truncate (1); elongate (2).
426. Scapus clavicle, direction of compression: craniocaudal (1); mediolateral (2).
427. Symphysis, orientation of compression: craniocaudally flattened (1); slightly dorsoventrally flattened (2); completely dorsoventrally flattened (3).
428. Processus acrocoracoideus, length: truncate (1); elongate (2).
429. Processus acromialis, length: truncate (1); elongate (2).
430. Processus acromialis, dorsocaudal orientation: dorsal (1); slightly caudal (2); caudal, at a 90-degree angle to scapus clavicle (3).
431. Processus acromialis, mediolateral orientation: does not deviate from axis of scapus clavicle (1); lateral (2); medial (3).
432. Scapus clavicle, lateral recurvature: absent (0); present (1).
Shoulder Girdle: Scapula
433. Collum, medial face, pointed, ventrally oriented projection: absent (0); present (1). Musser and Cracraft [
44], character 206; see also Musser et al. [
44], character 41 for further discussion.
434. Collum, medial face, large pneumatic foramen: present (1); lost (2). Mayr [
120], character 1248.
435. Collum, pronounced ventromedial bowing: absent (0); present (1).
436. Scapus, monotonic ventral curvature general to scapus, regardless of deviation of terminal end: straight to moderate, body and distal margin of scapula is slightly to moderately convex (1); pronounced, body and distal margin of scapula conspicuously convex (2). Musser and Cracraft [
44], character 207; Livezey and Zusi [
95], character 1260.
437. Scapus, lateral face, concavitas longitudinalis: present, concave throughout (1); lost, essentially planar throughout or shallow concavitas limited to cranial and medial portion (2). Musser and Cracraft [
44], character 208; Livezey and Zusi [
95], character 1257.
438. Scapus, ventrolateral face, tubercle of variable size located cranially, often accompanied by pitted crest trailing distally: present (1); lost (2). Musser and Cracraft [
44], character 209.
439. Dorsal angle: prominent, deviates from body of scapula (1); small or lost, does not interrupt curvature of body of scapula (2).
440. Dorsal angle of scapula, apex, if discernible: cranial to midpoint of shaft (1); at midpoint of shaft (2); caudal to midpoint of shaft (3).
441. Terminal margin, shape: rounded or subrectangular (1); acuminate (2).
442. Terminal margin, narrowing and/or expansion (regardless of prominence of dorsal angle): none (1); spatulation and expansion of terminus present (2); tapering present (3). Musser and Cracraft [
44], character 210; Livezey and Zusi [
95], character 1264.
443. Terminal margin, ventral deviation from scapus: no (1); yes (2).
444. Terminal margin, tip, dorsal orientation of terminus: no (1); yes (2).
Forelimb: Humerus
445. Tuberculum ventrale and crista m. scapulohumeralis caudalis, general form and caudal prominence: smoothed and often rounded, not very prominent (1); extremely prominent and laterally flattened, projects well beyond rest of bone caudally (2). Musser and Cracraft [
44], character 211; Livezey and Zusi [
95], character 1364.
446. Tuberculum ventrale, proximal elevation regardless of fossa pneumotricipitalis exposure: located inferior to tuberculum dorsale, projects cranially (1); subequal in elevation to that of tuberculum dorsale, projects cranially (2); elevated proximally to (above) tuberculum dorsale, projects cranially or cranioproximally (3). Musser and Cracraft [
44], character 213.
447. Tuberculum dorsale, proximodistally elongate: no (1); yes, tuberculum dorsale at least 1.5 times as long as it is wide (2). Musser et al. [
44], character 65.
448. Incisura capitis, depth: extremely deep and prominent (1); shallow (2). Musser and Cracraft [
44], character 215; Livezey and Zusi [
95], character 1358.
449. Incisura capitis, distal closure: not closed by additional structures distally (1); enclosed by distal projection of caput humeri (2); closed by transverse ridge (3). Musser and Cracraft [
44], character 221; Bertelli et al. [
14], character 54.
450. Fossa pneumotricipitalis ventralis: pneumatic (1); apneumatic (2). Musser and Cracraft [
44], character 216; Livezey and Zusi [
95], character 1414–1415; Mayr and Clarke [
63], character 77.
451. Fossa pneumotricipitalis (fossa pneumotricipitalis dorsalis): absent (0); present (1).
452. Fossa pneumotricipitalis dorsalis (if present), depth: shallow (1); deep and concave (2). Noncomparable where absent.
453. Proximal portion of humerus, caudal surface, fossa pneumotricipitalis, crus dorsale fossae: dorsoventrally narrow (1); dorsoventrally broad (2). Musser and Cracraft [
44], character 219; Livezey [
113], character 202.
454. Head, craniocaudally flattened: no, globose (1); yes (2). Musser and Cracraft [
44], character 220.
455. Head, distally reaching apex near ventral tubercle: present (1); lost (2).
456. Head, orientation: horizontal or dorsoventral (1); diagonal (2)
457. Impressio coracobrachialis: absent or shallow (1); deep (2). Musser and Cracraft [
44], character 222; Livezey and Zusi [
95], character 1428.
458. Sulcus ligamentosus transversus, dorsoventral length: abbreviate (1); elongate, at least half dorsoventral width of humeral head (2). Musser and Cracraft [
44], character 223; Livezey and Zusi [
95], character 1431.
459. Sulcus ligamentosus transversus, depth: shallow (1); deep (2). Musser and Cracraft [
44], character 223; Livezey and Zusi [
95], character 1431.
460. Bicipital crest, proximodistal length relative to that of deltopectoral crest: less than 1/2 length of deltopectoral crest (1); over 1/2 length of deltopectoral crest (2). Musser and Cracraft [
44], character 231; Livezey and Zusi [
95], character 1383.
461. Bicipital crest, terminus on body of humerus, ventral margin: abruptly discontinued proximally on corpus humeri, ventral margin (1); gradually continued by shallow, low but distinct jugum along corpus humeri, ventral margin (2) Musser and Cracraft [
44], character 227; Livezey and Zusi [
95], character 1413.
462. Head of humerus, separation from crista deltopectoralis and tuberculum dorsale (“externus”): pronounced, caput offset and well distinguished from tuberculum dorsale (1); diminished, caput low and poorly distinguished adjacent features (2). Musser and Cracraft [
44], character 228; Livezey and Zusi [
95], character 1354.
463. Crista deltopectoralis, orientation relative to head of humerus: cranial (1); craniolateral (2). Musser and Cracraft [
44], character 229; Livezey and Zusi [
95], character 1374.
464. Crista deltopectoralis, shape: rounded (1); trapezoidal (2).
465. Crista deltopectoralis, comparative proximodistal length relative to that of body of humerus: great, well developed and extending at least 1/3 length of corpus humeri (1); small, diminutive, extending less than 1/3 length of corpus humeri (2). Musser and Cracraft [
44], character 230; Livezey and Zusi [
95], character 1382.
466. Proximal humerus, long and narrow accessory scar for m. supracoracoideus: absent (0); present (1). Musser et al. [
44], character 61.
467. Proximal section of shaft with triangular cross section: absent (0); present (1). Musser and Cracraft [
44], character 232; Bertelli et al. [
14], character 56.
468. Caudal surface of proximal shaft with centrally positioned, raised muscle attachment scar for m. latissimus dorsi, pars caudalis: absent (0); present (1). Mayr (2013), character 20.
469. Fossa olecrani, depth: limited depth (1); present, markedly deep (2). Musser and Cracraft [
44], character 236; Livezey and Zusi [
95], character 1482.
470. Processus supracondylaris dorsalis greatly elongated proximodistally: no (1); yes (2). Musser and Cracraft [
44], character 241; Bertelli et al. [
14], character 57; Livezey and Zusi [
95], character 1467.
471. Flexor process, proximodistally elongate: no (1); yes (2).
472. Sulcus tendinis m. scapulotricipitalis: absent (0); present, weakly defined, typically broad, shallow and truncate (1); present, conspicuously defined, usually narrow, deep and elongate (2). Not comparable for Spheniscidae. Musser and Cracraft [
44], character 237; Livezey and Zusi [
95], character 1488.
473. Fossa m. brachialis, dorsoventral position relative to median axis of humerus: ventral (1); medial (2). Musser and Cracraft [
44], character 239; Livezey and Zusi [
95], character 1460.
474. Impression of brachialis anticus (brachial depression): shallow and small, ovoid, or brachial depression nonexistent (1); deep and part of brachial depression, distal portion of humerus appears to have been “scooped” out (2). Musser and Cracraft [
44], character 240; Livezey and Zusi [
95], character 1456; Mayr and Clarke [
63], character 80.
475. Condylus dorsalis humeri, proximal extent relative to distal margin of fossae m. brachialis: condylus distal to distal terminus of fossa and typically separated from distal margin of fossae m. brachialis by smooth area of bone from latter (1); condylus (proximal margin) typically extending at least proximal to distal margin of fossae m. brachialis (2); condylus markedly proximal to fossa, including extreme proximal condylorum dorsalis and ventralis (3). Musser and Cracraft [
44], character 244; Livezey and Zusi [
95], character 1447.
476. Condylus dorsalis humeri, proximal margin: rounded (1); acuminate (2). Musser and Cracraft [
44], character 243; Livezey and Zusi [
95], character 1451.
Forelimb: Ulna
477. Ulna greatly exceeding humerus in length: no (1); yes (2). Mayr and Clarke [
63], character 82.
478. Processus cotylaris dorsalis, pronounced ventral orientation such that apex processi is approximately coplanar with dorsal face of ulna: absent, apex of process variably elevated dorsally to body of ulna (0); present (1). Not comparable for Spheniscidae. Musser and Cracraft [
44], character 245; Livezey and Zusi [
95], character 1492.
479. Processus cotylaris dorsalis, facies articularis relative to that of cotyla ventralis: less expansive (1); subequal (2). Not comparable for Spheniscidae. Musser and Cracraft [
44], character 246; Livezey and Zusi [
95], character 1496.
480. Processus cotylaris dorsalis, crista intercotylaris: crista rudimentary but evident despite more typically conformed cotylae (1); variably prominent, cotylae dorsalis and ventralis distinct (2). Musser and Cracraft [
44], character 247; Livezey and Zusi [
95], character 1497.
481. Impressio insertii m. brachialis: absent or shallow (1); deep (2). Musser and Cracraft [
44], character 248; Livezey and Zusi [
95], character 1502.
482. Incisura radialis: absent or indistinct (0); present and variably pronounced (1). Musser and Cracraft [
44], character 249; Livezey and Zusi [
95], character 1505.
483. Trochlea humeroulnaris: no articular facet visible or exhibits shallow articular facet (1); exhibits deep articular facet (2).
484. Impressio scapulotricipitalis, depth: shallow (1); deep (2).
485. Olecranon process, proximal prominence: prominent (1); limited, at about level of cotylae (2).
486. Distal ulna, dorsal aspect, tab-like distally oriented projection along ventromedial margin of labrum condyli dorsalis: absent (0); present (1).
487. Olecranon fossa, depth: shallow (1); deep (2).
488. Depressio radialis, depth: shallow (1); deep (2).
489. Sulcus tendinis, depth: shallow (1); extremely deep and pit-like (2).
Forelimb: Manual Phalanges
496. Manual digit II: phalanx 1, index process: absent or barely protruding (1); present, prominent (2).
497. Manual II:1: not perforated (1); perforated (2). Bertelli et al. [
14], character 60; Strauch (1978), character 49.
Pelvis
498. Iliac blades and synsacrum: fused (1); unfused (2). Bertelli et al. (2013), character 178.
499. Thoracic vertebrae, if fused within preacetabular synsacrum and iliac blades, degree of fusion of transverse processes of thoracic vertebrae to iliac blades: transverse processes of 3 caudalmost thoracic vertebrae incompletely fused to preacetabular iliac blades (1); transverse processes of 1–2 caudalmost thoracic vertebrae incompletely fused to preacetabular iliac blades (2); transverse processes of all thoracic vertebrae completely fused to preacetabular iliac blades (3); transverse processes of all thoracic vertebrae incompletely fused to preacetabular iliac blades (4) Note: Here, incomplete fusion is defined by a lack of fusion across entire or almost entire ventral suture. Noncomparable where iliac blades completely unfused to synsacrum. Musser and Cracraft [
44], character 281.
500. Preacetabular and postacetabular ilium, dorsal face, relative craniocaudal lengths, indexed by ratio of length of former divided by length of latter: preacetabular ilium greater in craniocaudal length than postacetabular ilium, ratio significantly greater than unity (1); subequal (2); postacetabular ilium greater in craniocaudal length than preacetabular ilium, ratio less than unity (3). Musser and Cracraft [
44], character 255; Livezey and Zusi [
95], character 1890.
501. Preacetabular ilium, dorsal face, dorsomedial margin, dorsal iliac crest, dorsomedial synostosis forming carina iliaca dorsales: present (1); lost (2). Musser and Cracraft [
44], character 256; Livezey and Zusi [
95], character 1814; Mayr and Clarke [
63], character 792.
502. Preacetabular ilium and synsacrum, dorsal margin of separated preacetabular ilia, if not fused into carina iliaca dorsales: subequal to height of crista spinosa synsacri and postacetabular synsacrum (1); ventrally positioned so that dorsal edge of preacetabular ilia are well ventral to crista spinosa synsacri (2). Noncomparable where carina iliaca dorsales present.
503. Carina iliaca dorsales (if present) extent of dorsal fossae created on either side of crista spinosa synsacra: moderate, approximately half or less the length of the carina (1); extremely elongate, extending proximally over half of the carina, typically extending proximolaterally (2); essentially absent (3). Noncomparable where carina iliaca dorsales absent.
504. Preacetabular synsacrum, dorsal prominence of crista spinosa synsacra or carina iliaca dorsales relative to postacetabular synsacrum: subequal in dorsoventral height (1); dorsally prominent and elevated above level of postacetabular synsacrum (2). Musser and Cracraft [
44], character 257; Livezey and Zusi [
95], character 1819.
505. Preacteabular ilium, lateral face, angle respective to transverse plane of synsacrum: oblique or subvertical (1); subhorizontal (2). Musser and Cracraft [
44], character 258; Livezey and Zusi [
95], character 1823.
506. Preacetabular ilium, shape of anterior terminus: subrectangular, does not narrow cranially (1); rounded (sometimes exhibits incomplete ossification) (2); trapezoidal, narrows cranially (3); triangular, narrows cranially toward apex (4).
507. Preacetabular ilium, anterior terminus, extends anteriorly well beyond proximal margin of synsacrum: no (1); yes (2).
508. Preacetabular tubercles, status: present (1); lost (2). Musser and Cracraft [
44], character 259; Livezey and Zusi [
95], character 1810; Mayr and Clarke [
63], character 93.
509. Preacetabular tubercles, length: elongate (1); diminutive, close to body of pelvis (2). Noncomparable where preacetabulear tubercles absent. Musser and Cracraft [
44], character 259; Livezey and Zusi [
95], character 1810; Mayr and Clarke [
63], character 93.
510. Preacetabular tubercles, shape of termini: subrectangular (1); acuminate (2). Noncomparable where preacetabulear tubercles absent. Musser and Cracraft [
44], character 259; Livezey and Zusi [
95], character 1810.
511. Acetabular ilium, dorsal face, interacetabular width relative to synsacral length: great, exceeds 1/2 synsacral length (1); moderate, 1/2 to 1/3 synsacral length (2); small, approximately 1/4 synsacral length (3); extremely small, approximately 1/6 synsacral length (4). Musser and Cracraft [
44], character 260; Livezey and Zusi [
95], character 1845.
512. Acetabular and postacetabular dorsal synsacrum, osseous sheet covering fenestrae intertransversae synsacrales: yes (1); lost or incomplete along medial or lateral margins (2).
513. Antitrochanter, lateral face: raised dorsally so that it is more visible and robust (1); depressed ventrally so that it is less visible and somewhat obscures the acetabulum (2). Musser and Cracraft [
44], character 263.
514. Antitrochanter, craniolaterally oriented semicircular prominence on cranial margin of antitrochanter that thickens ventrally: absent (0); present (1). Musser and Cracraft [
44], character 264.
515. Ilioischiadic foramen: absent (0); oblong (1); essentially circular (2). Musser and Cracraft [
44], character 265; Livezey and Zusi [
95], character 1791.
516. Obturator foramen, closed caudally: no (1); yes (2). Musser and Cracraft [
44], character 268; Livezey and Zusi [
95], character 1783.
517. Obturator foramen, shape: circular, often with wider dorsoventral margin (1); elongate, ovoid (2). Musser and Cracraft [
44], character 268; Livezey and Zusi [
95], character 1783.
518. Obturator foramen, axis of orientation: dorsal cranially and ventral caudally (1); craniocaudal or horizontal (2); ventral cranially and dorsal caudally (3). Musser and Cracraft [
44], character 269.
519. Vertex craniolateralis ilii, dorsally protrudes above antitrochanteric sulcus: no (1); yes (2).
520. Vertex craniolateralis ilii, if dorsally protruding above antitrochanteric sulcus, shape: crest-like (1); elongate and tab-like, typically laterally directed (2). Noncomparable where not dorsally protruding.
521. Vertex caudolateralis ilii: tab-like laterally directed (1); a simple crest (2); lost (3); a ventrally directed flange (4). Noncomparable for Paleognathae. Musser and Cracraft [
44], character 272; Livezey [
113], character 286.
522. Postacetabular ilium and ischium, lateral face, fusion: unfused posteriorly (0); fused posteriorly, ilioischiatic fenestra is closed (1). Musser and Cracraft [
44], character 273; Cracraft and Clarke [
115], character 27.
523. Postacetabular ilium and ischium, lateral face, synchondrosis ilioischiadica: extremely smooth, barely visible or absent (1); line-like and distinctly etched craniocaudally or caudoventrally (2). Noncomparable for Paleognathae. Musser and Cracraft [
44], character 275.
524. Postacetabular ilium and ischiium, lateral face, deep incisura in caudal margin: absent (0); present (1). Noncomparable for Paleognathae. Musser and Cracraft [
44], character 276; Bertelli et al. [
14], character 65; Livezey and Zusi [
95], character 1787.
525. Caudal extent of caudal terminus of postacetabular ilium relative to that of ischium: ilium distinctly cranial to ischium (1); ischium distinctly cranial to ilium (2); subequal (3). Noncomparable for Paleognathae. Musser and Cracraft [
44], character 278; Livezey and Zusi [
95], character 1892.
526. Preacetabular synsacrum, ventral face, large spine protruding from cranial portion: absent (0); present (1). Musser and Cracraft [
44], character 279.
527. Acetabular synsacrum, ankylosis of acetabular transverse process (often synostotic) to acetabular area beyond lateral margin of synsacrum: absent, transverse process does not ankylose to acetabular area (0); present, ankyloses to dorsal margin of acetabulum, often within dorsocaudal margin of the acetabulum (1); present, ankyloses to craniodorsal margin of ilioishciadic foramen with variable separation between acetabulum and ankylosis, accompanied by sessile concavity between ankylosis and margin of acetabulum (2); present, ankyloses to ventral site between caudal acetabular margin and cranial margin of ilioischiadic foramen, ankylosed to pila that extends from ankyloses between the ilioischiadic foramen toward the dorsal margin of the obturator foramen (3). Noncomparable for Paleognathae. Musser and Cracraft [
44], character 280.
528. Preacetabular and acetabular synsacrum, ventral face, ventral sulcus of synsacrum: present (1); lost or barely discernible (2). Musser and Cracraft [
44], character 282.
529. Postacetabular ischium, ventral face, foramen in oblique iliac crest: present (1); lost (2). Musser and Cracraft [
44], character 283.
530. Postacetabular portion of synsacrum, dorsoventral position: postacetabular portion of synsacrum depressed dorsally (1); postacetabular portion of synsacrum raised ventrally (2). Musser and Cracraft [
44], character 284; Livezey and Zusi [
95], character 1793.
531. Fossa renalis, depth: shallow or lost (1); deep (2). Musser and Cracraft [
44], character 284; Livezey and Zusi [
95], character 1793.
532. Lamina of ischium anteriorly extensive so as to create pons for recessus caudalis fossae: yes (1); no (2). Noncomparable for Paleognathae. Musser and Cracraft [
44], character 286; Livezey and Zusi [
95], character 1802.
533. Ischial lamina creating pons for recessus caudalis fossae (if present), visible through ilioischiadic foramen in lateral aspect: no (1); yes (2). Noncomparable where recessus caudalis fossae absent.
534. Recessus caudalis fossae (if present): shallow (1); deep (2). Noncomparable where absent. Musser and Cracraft [
44], character 286; Livezey and Zusi [
95], character 1802; Mayr and Clarke [
63], character 95.
535. Deep ovoid fossa along caudomedial length of postacetabular ilium and ischium forming recessus caudalis fossae (if present): absent (0); present (1). Noncomparable where absent. Musser and Cracraft [
44], character 287; Livezey and Zusi [
95], character 7427.
536. Caudal margin of postacetabular ilium and in relation to caudal portion of synsacrum: caudal portion of synsacrum separating from and continuing caudally past caudal margin of ilium and splaying laterally towards apex (1); caudal portion of synsacrum fused to caudal margin of ilium completely (2); caudal portion of synsacrum separating from and continuing caudally past caudal margin of ilium and tapered toward apex (3). Musser and Cracraft [
44], character 289.
537. Caudalmost synsacral vertebrae, fused to costal processes of closest proximal vertebra: no (1); yes (2).
538. Caudalmost synsacral vertebrae, if fused to costal processes of closest proximal vertebra, number of fused vertebrae present: 2 (1); 3 or more (2). Noncomparable where unfused to costal processes.
539. Iliosynsacral incisura, shape: semicircular (1); subrectangular (2).
540. Processus marginis caudalis, shape of terminus: subrectangular (1); acuminate (2); rounded, tab-like (3). Musser and Cracraft [
44], character 291; Livezey and Zusi [
95], character 1808.
541. Processus marginis caudalis, termini invaginated on caudodorsal surface so that termini are medially oriented: no (1); yes (2). Musser and Cracraft [
44], character 292.
542. Processus terminalis ischii, shape: acuminate (1); subrectangular (2); rounded (3). Musser et al. [
44], character 82.
543. Processus terminalis ischii, ankylosed to pubis: no (1); yes (2).
544. Spatium ischiopubicum, dorsoventral width: wide (1); narrow, ischium and pubis nearly in contact (2).
545. Scapus pubis, exclusive of marked departures in apex: recurved, dorsal margin variably concave (1); essentially straight or slightly sigmoid, approximately aligned with major axis of apex of pubis (2); decurved, dorsal margin convex (3). Musser and Cracraft [
44], character 294; Livezey and Zusi [
95], character 1928.
546. Scapus and apex of pubis, length of extension beyond ischium: pubis not extending beyond caudal margin of ischium or, if it does, extended portion makes up less than one third of entire length of pubis (1); portion extending beyond caudal portion of ischium makes up at least one fourth that of entire length of pubis or more (2). Musser and Cracraft [
44], character 295; Livezey and Zusi [
95], character 2006.
547. Apex pubis, dorsoventral orientation: projecting dorsally (1); projecting ventrally (2). Musser and Cracraft [
44], character 296; Livezey and Zusi [
95], character 1945.
548. Apex pubis, dorsomedial form: dorsoventrally spatulate and cruciate (1); spatulation negligible, apex pubis subparallel along ventral and dorsal margins, terminal end subrectangular or acuminate (2). Note: Not to be confused with dorsal angle (can be seen in Rallidae) that is a point of attachment between pubis and ischium. Musser and Cracraft [
44], character 297; Livezey and Zusi [
95], character 1940.
549. Apex pubis, mediolateral curvature: absent or slight (1); present, prominent departure from craniocaudal axis, apex of pubis medial to caudal ischium (2); present, apex of pubis curves lateral to caudal ischium (3).
Ossified Tendons
550. Intratendinous ossification in hindlimbs: absent (0); present (1). Musser and Cracraft [
44], character 298; Bertelli et al. [
14], character 80.
551. Intratendinous ossification in forelimbs: absent (0); present (1).
Femur
552. Crista trochanteris, markedly projected cranially: yes (1); no (2). Musser and Cracraft [
44], character 299; Mayr and Clarke [
63], character 97.
553. Crista trochanteris, caudal face distal to facies articularis antitrochantericus: thickening developed (1); lacking distinct, distal thickening (2). Musser and Cracraft [
44], character 300; Livezey [
113], character 301.
554. Fossa trochantericus: lost or shallow (1); present, deep and extends across entire width of facies articularis antitrochantericus (2). Musser and Cracraft [
44], character 301; Livezey and Zusi [
95], character 1978.
555. Collum femoris, facies articularis antitrochanterica, caudal margin: located at 90-degree angle to femoral body (1); projecting proximally (2). Musser and Cracraft [
44], character 303; Livezey and Zusi [
95], character 1975.
556. Proximal femur, pneumatic foramen on craniolateral side: absent (0); present (1). Mayr and Clarke [
63], character 98.
557. Sulcus patellaris, cranial aspect, ovate accessory subfossa in proximal region: absent (0); present (1). Musser and Cracraft [
44], character 305; Livezey and Zusi [
95], character 2039.
558. Fossa poplitea: shallow, weakly delimited (1); deep (2). Musser and Cracraft [
44], character 306; Livezey and Zusi [
95], character 2047.
559. Fossa poplitea, pneumatic foramen: small or absent (1); present, large (2). Musser and Cracraft [
44], character 307; Livezey and Zusi [
95], character 2048.
560. Impressio ligamentum collateralis lateralis, depth: shallow (1); deep (2).
561. Epicondylus medialis, prominence of tubercle: prominently elevated, typically tubercular (1); not prominently elevated, grading essentially smoothly (2). Musser and Cracraft [
44], character 311; Livezey [
113], character 308.
562. Condylus lateralis, crista tibiofibularis, distal extent relative to condylus medialis: subequal to condylus medialis (1); distinctly and significantly distal to condylus medialis (2). Musser and Cracraft [
44], character 312; Livezey and Zusi [
95], character 2012.
563. Shaft, cranial recurvature, status: absent (0); present (1).
564. Condylus lateralis, caudal aspect, lateromedial width relative to that of condylus medialis: subequal (1); lateral condyle significantly wider (2); lateral condyle significantly narrower (3). Musser and Cracraft [
44], character 314; Livezey and Zusi [
95], character 2027.
565. Condylus lateralis, relative proximodistal position of crista tibiofibularis and trochlea fibularis: crista tibiofibularis crest located more proximally than trochlea fibularis (1); subequal in proximal position (2); trochlea fibularis more proximal (3). Musser and Cracraft [
44], character 315; Livezey and Zusi [
95], character 2010.
566. Condylus lateralis, relative caudal prominence of trochlea fibularis and crista tibiofibularis: caudal prominence subequal (1); crista tibiofibularis more prominent than trochlea fibularis (2).
567. Condylus medialis, caudal aspect, general form: spherical, distal portion greatly rounded, extremely convex (1); triangular, distal portion somewhat rounded (2). Musser and Cracraft [
44], character 316; Livezey and Zusi [
95], character 2030.
568. Condylus medialis, medial aspect, cranial face of body, proximal terminus: abrupt, subperpendicular or acuminate (1); comparatively gradual (2). Musser and Cracraft [
44], character 317; Livezey and Zusi [
95], character 2032.
569. Crista supracondylaris medialis, medial aspect, form relative to condylus medialis and facies caudalis corporis: absent or rudimentary, condylus not extended by crista, caused by one distinct (often abrupt) angle or incision above the medial condyle, rarely also a second is present (comparatively proximal), interrupting an otherwise gradual curving crest (1); present and prominent, medial condyle continued by crest without interruption by angle or incision above the medial condyle (2). Musser and Cracraft [
44], character 318; Livezey and Zusi [
95], character 2042.
570. Fovea tendineus m. tibialis cranialis, depth: shallow (1); deep (2).
571. Fovea tendineus m. tibialis cranialis, distal aspect, location: located cranially (1); located caudally (2). Musser and Cracraft [
44], character 320; Livezey and Zusi [
95], character 7428.
572. Femur, length compared to that of tibiotarsus: 1/2 length of tibiotarsus (1); over 1/2 length of tibiotarsus (2); less than 1/2 length of tibiotarsus (3).
Tibiotarsus and Fibula
573. Cnemial fossa retropatellaris: shallow (1); deep (2). Musser and Cracraft [
44], character 321; Livezey and Zusi [
95], character 2112.
574. Cnemial fossa retropatellaris, shape: circular (1); elongate and ovoid (2).
575. Cnemial fossa retropatellaris or homologous site, recessus and/or pneumatic foramina (pores): no (1); yes (2). Musser and Cracraft [
44], character 322; Livezey and Zusi [
95], character 2113.
576. Lateral fossa retropatellaris: shallow (1); deep (2).
577. Lateral fossa retropatellaris or homologous site, recessus and/or pneumatic foramina (pores): no (1); yes (2).
578. Incisura in caudal aspect of interarticular area delimiting facies articularis medialis: present (1); lost (2). Musser and Cracraft [
44], character 323; Livezey and Zusi [
95], character 7429.
579. Pneumatic foramen underneath jugum between facies articularis lateralis and area interarticularis (typically found distal to incisura, if present): present, 1 (1); present, 2 or more (2); lost (3). Musser and Cracraft [
44], character 324.
580. Fossae retropatellares, angle relative to facies articularis medialis and lateralis: on same plane as articular area (1); proximocranially oriented between 45 and 80 degrees (2); proximocranially oriented at approximately 90 degrees so that fossae are perpendicular to articular areas (3).
581. Crista cnemialis and crista patellaris, apex, proximodistal height relative to head of tibiotarsus: at same height of fossae retropatellares or only slightly elevated above fossae (1); well projected proximally to fossae (2); extremely proximally projected, at least 3 times the length of the fossae above the fossae (3).
582. Crista cnemialis lateralis apex, proximodistal height relative to head of tibiotarsus: essentially at level of fossae retropatellares (1); proximal to fossae retropatellares (2); extremely proximally projected, at least 3 times the length of the fossae above the fossae (3); extremely distally projected (4).
583. Crista cnemialis lateralis and cranialis, distal apices (distal angles of cristae, not attachment locations on shaft of tibiotarsus), relative proximodistal heights: subequal (1); apex of crista cnemialis lateralis located proximally to that of crista cnemialis cranialis (2); apex of crista cnemialis cranialis proximal to that of crista cnemialis lateralis (3).
584. Crista cnemialis cranialis, pronounced lateral curvature (especially proximally): present (1); lost (2). Musser and Cracraft [
44], character 329; Livezey and Zusi [
95], character 2082.
585. Crista cnemialis lateralis, shape: hamate but intermediate, but hook rounded and does not protrude beyond jugum of lateral crista (1); more prominent and hamate, acuminate hook protrudes distolaterally beyond lateral crest jugum (2); rounded, tuberculate (3).
586. Crista cnemialis cranialis, especially cranially projected: no, but is still well projected (1); yes (2); no, extremely reduced (3).
587. Crista cnemialis cranialis, distal apex, shape: rounded (1); acuminate (2).
588. Crista cnemialis cranialis, apex of crista cnemialis cranialis and crista patellaris or apex of crista cnemialis cranialis only, shape: apex lost (0); rounded (1); acuminate (2).
589. Crista cnemialis cranialis, concavity of lateral and medial surfaces: both surfaces concave (1); medial surface convex, lateral surface concave (2); both surfaces convex (3); medial surface concave, lateral surface convex (4). Musser and Cracraft [
44], character 328; Livezey and Zusi [
95], character 2078.
590. Crista cnemialis lateralis, concavity of lateral and medial surfaces: both surfaces concave (1); medial surface convex, lateral surface concave (2); both surfaces convex (3); medial surface concave, lateral surface convex (4). Musser and Cracraft [
44], character 328; Livezey and Zusi [
95], character 2078.
591. Crista cnemialis lateralis, proximal crista, width compared to crista patellaris: extremely wide (1); narrow, subequal in width to crista patellaris (2).
592. Lateral fossa retropatellaris, extremely reduced: no (1); yes (2).
593. Facies articularis medialis: convex (1); concave (2).
594. Crista between fossae retropatellares: absent (0); present (1).
595. Facies articularis lateralis, concavity: absent or small (1); present, large (2).
596. Interarticular area: convex (1); concave (2).
597. Crista cnemialis lateralis, lateral prominence relative to articulated fibula: not lateral to fibular head (1); lateral to fibular head (2). Musser and Cracraft [
44], character 331; Livezey and Zusi [
95], character 2094.
598. Cristae cnemiales cranialis and lateralis, comparative distal extents on shaft of tibiotarsus: crista cnemialis cranialis terminating distinctly distal to crista cnemialis lateralis (1); cristae subequal in distal extent (typically truncated), both lacking jugae (2). Musser and Cracraft [
44], character 332; Livezey and Zusi [
95], character 2099.
599. Crista cnemialis cranialis, recurvature of distal margin: absent (0); present (1).
600. Fibula, head, form and position relative to head of tibiotarsus: elongate and extends caudally past interarticular area (1); essentially circular, from proximal view does not extend past interarticular area (2). Musser and Cracraft [
44], character 333.
601. Fibular crest, length: over 1/4 length of tibiotarsus (1); about 1/4 length of tibiotarsus (2); less than 1/4 length of tibiotarsus (3).
602. Fibular crest, lateral prominence: little to none (1); prominently projected (2).
603. Fibular crest, if projected laterally, relative lateral prominence of proximal and distal ends: distal end more laterally projected (1); subequal (2). Not comparable where fibular crest not projected laterally.
604. Fibular crest, proximodistal location: just distal to head of tibiotarsus (1); close to midshaft of tibiotarsus (2). Musser and Cracraft [
44], character 330; Livezey and Zusi [
95], character 2085.
605. Foramen interosseum proximale: sublinear and incisurate, relatively narrow (1); approximately ovate, relatively spacious (2). Musser and Cracraft [
44], character 335; Livezey and Zusi [
95], character 2128.
606. Foramen interosseum distale, length as compared to length of the tibiotarsus: more than 1/4 length of the tibiotarsus (1); less than 1/4 length of tibiotarsus (2); exactly 1/4 length of tibiotarsus (3). Musser and Cracraft [
44], character 336; Livezey and Zusi [
95], character 2129.
607. Proximal shaft of tibiotarsus, cranial aspect: rounded (1); craniocaudally flattened and mediolaterally broad (2).
608. Epicondylus medialis: pronounced tubercle (1); lost or diminutive (2). Musser and Cracraft [
44], character 337.
609. Epicondylus lateralis: lost or diminutive (1); pronounced tubercle (2).
610. Trochlea cartilaginis tibialis, lateral and medial margins: splayed laterally (1); medially compressed (2); only medial margin splayed laterally (3). Musser and Cracraft [
44], character 338.
611. Trochlea cartilaginis tibialis, height: craniocaudally short (1); craniocaudally elongate (2). Musser and Cracraft [
44], character 339.
612. Trochlea cartilaginis tibialis, rectangular, medially oriented projection on medial margin: absent (0); present (1). Musser and Cracraft [
44], character 340.
613. Condylus lateralis, depressio epicondylaris lateralis: shallow or absent (1); deep (2). Musser and Cracraft [
44], character 341.
614. Condylus lateralis, depressio epicondylaris lateralis, accessory depression located along caudal margin of condylus lateralis: absent (0); present (1). Musser and Cracraft [
44], character 341.
615. Condylus lateralis, depressio epicondylaris lateralis, accessory depression (if present), depth: shallow (1); deep (2). Noncomparable where depressio epicondylaris lateralis absent.
616. Condylus lateralis, proximocranial rim, tubercle: absent (0); present (1).
617. Condylus medialis, depressio epicondylaris medialis: shallow or absent (1); deep (2).
618. Condylus medialis, depressio epicondylaris medialis, accessory depression located along caudal margin of condylus medialis: absent (0); present (1).
619. Condylus medialis, depressio epicondylaris medialis, accessory depression (if present), depth: shallow (1); deep (2). Noncomparable where depressio epicondylaris absent.
620. Condylus medialis and condylus lateralis, relative positions: pulled in medially (1); spread laterally (2).
621. Medial and lateral condyles, incisura intercondylaris, distal aspect, foveae (sulcus) transcondylares medialis and lateralis: present (1); lost (2). Musser and Cracraft [
44], character 344; Livezey and Zusi [
95], character 2155.
622. Tuberositas retinaculi extensorius lateralis, ossified retinaculum extensorum where medial portion is almost completely ossified or is completely ossified (retinaculum m. fibularis): absent (0); present (1).
623. Tuberositas retinaculi extensorius lateralis, if ossified retinaculum absent, comparative prominence of lateral and medial cristae: subequal (1); medial crista more prominent (2); lateral crista more prominent (3).
624. Tubercle laterodistal to pons supratendinous: absent (0); present (1). Mayr and Clarke [
63], character 101.
625. Tubercle laterodistal to pons supratendinous (if present), prominence: diminuitive (1); prominent (2). Noncomparable where tubercle absent. Mayr and Clarke [
63], character 101.
626. Crista extending proximally from tubercle laterodistal to pons supratendinous or homologous site: absent (0); present (1).
627. Crista extending proximally from tubercle laterodistal to pons supratendinous or homologous site (if present), length: proximodistally truncate (1); proximodistally elongate (2). Noncomparable where crista absent.
628. Crista extending proximally from tubercle laterodistal to pons supratendinous or homologous site (if present), foramen at base: absent (0); present (1). Noncomparable where crest absent.
629. Tuberositas retinaculi extensorius medialis: absent (0); present (1).
630. Medial and lateral condyles, relative cranial extensions (compare looking at distal surface): subequal (1); medial condyle extended more cranially (2).
631. Medial and lateral condyles, relative mediolateral thickness (compare looking at distal surface): subequal (1); medial condyle mediolaterally thinner (2); lateral condyle thinner (3).
632. Tuberositas retinaculi extensorius medialis (if present), form: tab-like, prominent (1); small and diminutive crest (2). Noncomparable where absent.
633. Tuberositas retinaculi extensorius medialis (if present), craniomedial orientation: oriented medially (1); oriented cranially (2). Noncomparable where absent.
634. Tuberositas retinaculi extensorius medialis (if present), proximodistal location: just above pons supratendinous (1); proximally located (2). Noncomparable where absent.
635. Tuberositas retinaculi extensorius lateralis, proximodistal location: just above pons supratendinous (1); proximally located (2); extends distal to pons supratendinous (3).
636. Tuberositas retinaculi extensorius lateralis, extreme lateral extension: no (1); yes (2).
637. Pons supratendinous, location of distal opening: above medial condyle (1); just lateral to medial condyle but within medial margin of tibiotarsus (2); centered along midline (3).
638. Canalis extensorius, depth: shallow (1); deep (2); extremely deep and cavernous (3).
639. Medial and lateral condyles, relative proximal extents: lateral condyle located more proximally than medial condyle (1); subequal (2); medial condyle located more proximally than lateral condyle (3).
640. Medial condyle, distal rim distinctly notched: no (1); yes (2). Mayr and Clarke [
63], character 102.
Tarsometatarsus
641. Medial cotyle, medial margin, exceptionally projected proximally and crista-like: no (1); yes (2).
642. Medial cotyle, plantar margin, exceptionally projected proximally and crista-like: no (1); yes, moderately projected to roughly the same level of the proximal margin of the hypotarsal eminence (2); yes, prominently projected proximal to hypotarsal eminence (3).
643. Medial cotyle, proximodistal orientation of dorsal margin: cotyla perpendicular to shaft of tarsometatarsus, proximal surface of cotyla not visible in dorsal aspect (1); dorsal margin of cotyla positioned slightly more distally than that dorsal half of cotyla somewhat visible in dorsal aspect (2); dorsal margin of cotyla positioned much more distally than that of plantar margin so that cotyla is largely visible in dorsal aspect (3).
644. Lateral cotyle, dorsal margin, crista-like margin and/or subtending angle present: yes (1); no, dorsal margin of cotyle essentially absent and cotyla is coplanar with and almost indistinguishable from cotyla corpus (2).
645. Lateral cotyle, depth: deep, concave (1); flat or only slightly concave (2).
646. Lateral cotyle, proximodistal orientation of dorsal margin: dorsal margin of cotyla positioned slightly more distally than that dorsal half of cotyla somewhat visible in dorsal aspect (1); cotyla perpendicular to shaft of tarsometatarsus, proximal surface of cotyla not visible in dorsal aspect (2); dorsal margin of cotyla positioned much more distally than that of plantar margin so that cotyla is largely visible in dorsal aspect (3).
647. Hypotarsal eminence, proximally prominent: no (1); yes (2).
648. Lateral and medial cotylae (dorsal perspective), relative proximal elevation: cotyla lateralis distinctly distal to cotyla medialis (1); cotyla lateralis subequal to cotyla medialis (2); cotyla medialis distinctly distal to cotyla lateralis (3). Musser and Cracraft [
44], character 346; Livezey and Zusi [
95], character 2250.
649. Fossa parahypotarsalis lateralis, depth: shallow or barely visible (1); deep (2). Musser and Cracraft [
44], character 347; Livezey and Zusi [
95], character 2258.
650. Fossa parahypotarsalis medialis, depth: shallow or barely visible (1); deep (2). Musser and Cracraft [
44], character 348; Livezey and Zusi [
95], character 2257.
651. Hypotarsus, crista(e) medialis hypotarsi and crista(e) lateralis hypotarsi, relative plantar prominence: crista(e) medialis hypotarsi more plantarly prominent than crista(e) lateralis hypotarsi (1); crista(e) subequal in plantar prominence (2); crista(e) medialis hypotarsi less plantarly prominent than crista(e) lateralis hypotarsi (3). Musser and Cracraft [
44], character 349; Livezey [
113], character 336.
652. Crista medialis hypotarsi and crista lateralis hypotarsi, relative distal extents and magnitude: distal-most terminus(i) of medial crest(s) much more distally extensive, lateral crista(e) proximodistally truncate, about 1/2–2/3 proximodistal length of the medial crista (1); subequal (2); distal-most terminus(i) of lateral crest(s) more distally extensive, medial crista(e) proximodistally truncate, approximately 1/2 proximodistal length of the lateral crista, and reduced to a mediolaterally thin, osseous sheet (3); distal-most terminus(i) of lateral crest(s) more distally extensive, medial crista(e) proximodistally truncate, reduced to at least approximately 1/3 or less proximodistal length, thin lamina (4).
653. Hypotarsus, tendon of m. flexor digitorum longus (FDL) enclosed by a canal: no, sulcus present for m. (1); m. enclosed in a canal (2). Noncomparable where muscle absent or sulcus/canal for muscle absent/indiscernible, or where homology is uncertain. Musser and Cracraft [
44], character 350; Livezey and Zusi [
95], characters 2284–2286; Mayr and Clarke [
63], character 103.
654. Hypotarsus, canal for tendon of m. flexor digitorum longus (FDL, if present), centrally located: no (1); yes (2). Noncomparable where not enclosed in canal.
655. Hypotarsus, tendon of m. flexor perforatus digiti II (FPII) enclosed in a canal: no, sulcus present for m. (1); m. enclosed in a canal (2). Noncomparable where muscle absent or sulcus/canal for muscle absent/indiscernible, or where homology is uncertain. Musser and Cracraft [
44], character 352; Bertelli et al. [
14], character 75.
656. Hypotarsus, tendon of m. flexor perforans et perforatus digiti II (FPPII) enclosed in a canal: no, sulcus present for m. (1); m. enclosed in a canal (2). Noncomparable where muscle absent or sulcus/canal for muscle absent/indiscernible, or where homology is uncertain.
657. Hypotarsus, tendon of m. flexor hallicis longus (FHL) enclosed in a canal: no, sulcus present for m. (1); m. enclosed in a canal (2). Noncomparable where muscle absent or sulcus/canal for muscle absent/indiscernible, or where homology is uncertain.
658. Fossa infracotylaris dorsalis, depth: shallow (1); deep (2). Musser and Cracraft [
44], character 354; Livezey and Zusi [
95], character 2259; Mayr and Clarke [
63], character 104.
659. Proximal tarsometatarsus, dorsal margin of lateral cotyla, additional, subtriangular, distal-reaching tuberositas present that may be concave laterally: absent (0); present (1).
660. Proximal tarsometatarsus, dorsal aspect (medial portion), arcus extensiorius (ossified retinaculum): not present, remains of retinaculum are only impressio retinaculi extensorii (1); present (2). Mayr [
120], character 35.
661. Proximal portion of tarsometatarsus, dorsal aspect, foramina vascularia proximalia: essentially equal in height (1); lateral foramina distinctly distal to medial foramina (2); medial foramina distal to lateral foramina (3). Musser and Cracraft [
44], character 353; Livezey and Zusi [
95], character 2264.
662. Sulcus extensorius, proximal portion medial to dorsal foramina vascularia proximalia, depth: essentially absent (1); present, shallow (2); present, deep (3).
663. Sulcus extensorius, proximal portion lateral to dorsal foramina vascularia proximalia, depth: essentially absent (1); present, shallow (2); present, deep (3).
664. Dorsal foramina vascularia proximalia, enclosed in ovoid depression that is plantar to plane of sulcus extensorius: yes (1); no, foramina coplanar with sulcus extensorius (2); yes, but only lateral foramen is plantar to sulcus extensorius (3). Musser and Cracraft [
44], character 354; Livezey and Zusi [
95], character 2259.
665. Proximal portion of tarsometatarsus, dorsal aspect, foramina vascularia proximalia, proximodistally elongate: no (1); yes (2).
666. Proximal portion of tarsometatarsus, plantar aspect, foramina vascularia proximalia, proximodistally elongate: no (1); yes (2); only one is proximodistally elongate, typically medial foramen (3).
667. Proximal portion of tarsometatarsus, dorsal aspect, additional foramen or pair of circular foramina proximal to foramina vascularia proximalia that do not open to plantar face: absent (0); present, one foramen (1); present, pair of foramina (2).
668. Proximal portion of tarsometatarsus, plantar aspect, foramina vascularia proximalia, number: a pair is present (1); a pair is present with an additional foramen located on the lateral side (2); only one foramen is present (3); a pair is present with at least an additional foramen located on the medial side (4). Musser and Cracraft [
44], character 355.
669. Proximal portion of tarsometatarsus, plantar aspect, foramina vascularia proximalia: lateral foramina distinctly distal to medial foramina (1); lateral and medial foramina about equal in height (2). Noncomparable where only one foramen is present. Musser and Cracraft [
44], character 356. Ksepka and Clarke [
121], character 78.
670. Sulcus extensorius, depth along shaft of tarsometatarsus: shallow or absent (1); deep sulcus depth is roughly half that of tarsometatarsus (2); cavernous, only planter lamina of tarsometatarsus present (3). Musser and Cracraft [
44], character 357; Livezey and Zusi [
95], character 2305.
671. Sulcus extensorius, medial and lateral cristae, relative dorsal prominence: medial crista more dorsally prominent than lateral crista (1); subequal (2); lateral crista more dorsally prominent than medial crista (3).
672. Tuberositas m. tibialis cranialis, dorsal prominence: prominent and raised dorsal to sulcus extensorius (1); diminutive, barely raised dorsally (2).
673. Tuberositas m. tibialis cranialis, number: 1 (1); 2 (2). Noncomparable for taxa with scoring (1) for character (canal) due to fusion of tuberosities.
674. Tuberosities m. tibialis cranialis, relative proximodistal lengths: subequal in proximodistal length (1); medial tuberosity much more proximodistally elongate (2). Noncomparable where one tuberositas present or where tuberosities fused.
675. Tuberosities m. tibialis cranialis, subtending ossified bridge and canal opening distally along midline of tarsometatarsus: absent (0); present (1).
676. Processus calcaris: absent or diminutive (1); present (2).
677. Fossa supratrochlearis plantaris: absent or indistinct (0); present, distinctly concave (1). Musser and Cracraft [
44], character 363; Livezey and Zusi [
95], character 2329.
678. Trochlea metatarsi II, plantarly prominent eminentia (medio) plantaris (ala): absent (0); present (1).
679. Trochlea metatarsi II, eminentia (medio) plantaris (ala): rounded (1); subrectangular (2). Noncomparable in absence of ala. Musser and Cracraft [
44], character 364; Livezey and Zusi [
95], character 2352.
680. Trochlea metatarsi II, eminentia (medio) plantaris (ala, if present): projects plantarly (1); projects plantomedially (2); projects laterally (3). Noncomparable in absence of ala.
681. Trochlea metatarsi III, proximodistal length as compared to that of body of tarsometatarsus: length from proximal to distal end is less than 1/5 length of entire tarsometatarsus from top of eminentia intercotylaris to distal end of trochlea metatarsi III (1); length from proximal to distal end is 1/5 or greater length of entire tarsometatarsus (2). Musser and Cracraft [
44], character 365.
682. Trochlea metatarsi IV, foveae ligamentorum collateralium, status and form sensu depth and width relative to that of associated trochlea: present, moderate depression (1); small, shallow, almost absent (2). Musser and Cracraft [
44], character 366; Livezey and Zusi [
95], character 2349.
683. Trochleae metatarsalia II-IV, distal view, relative dorsal elevations: II < III ≥ IV and II < IV (1); II < III > IV and II = IV (2); II = III = IV (3). Musser and Cracraft [
44], character 367; Livezey and Zusi [
95], character 2363.
684. Trochleae metatarsalia II-IV, relative distal extents: II < III ≥ IV and II < IV (1); II < III > IV and II subequal to IV (2); II > IV, III ≥ IV (3).
685. Distal portion of tarsometatarsus, trochlea metatarsi II, facies articularis phalangealis, sulcus trochlearis (narrow groove between lateral and medial rims of trochlea): remains distinct on facies dorsalis (1); obsolete on facies dorsalis, terminating medially toward fovea ligamentorum collaterallium at distal apex of trochlea (2). Musser and Cracraft [
44], character 368; Livezey [
113], character 354.
686. Trochlea metatarsi II, plantarly deflected: no (1); yes (2). Mayr and Clarke [
63], character 108.
687. Trochlea metatarsi IV, plantar ala: not prominent, plantar extent significantly less than that of trochlea metatarsi II (1); conspicuously prominent, plantar extent subequal to that of trochlea metatarsi II (2). Note: not to be confused with flange on plantar surface of trochlea metatarsi II, common in many genera.
688. Pedal digit 4: phalanges 3 and 4, relative lengths: phalanx 4 longer (1); both phalanges of equal length (2); phalanx 4 shorter (3). Bertelli et al. [
14], character 76.
689. Three anterior toes notably elongated: absent (0); present (1). Bertelli et al. [
14], character 77.
690. Pedal I:1, pronounced bowing: absent (0); present (1). Musser et al. [
44], character 97.
691. Pedal I:1, length as compared to III:1: not as follows (1); I:1 about half the length of III:1 (2). Mayr and Clarke [
63], character 110; Mayr [
47], character 32.
692. Pedal IV:1, medially protruding phalange at proximal margin: absent (0); present (1). Mayr [
47], character 35.
693. Proximal end of pedal II:2 extending little beyond proximal end of III:2: no (1); yes (2). Mayr [
47], character 33.
694. Olfactory bulb, size: absent (0); present (1).
695. Olfactory bulb, position: ventral (0); dorsal (1).
696. Olfactory bulb, conformation: clearly bifurcated (0); single tract (1).
697. Telencephalon, shape in dorsal view: rounded (0); anteriorly tapered (1).
698. Wulst, dorsal expansion in anterior view: not expanded (0); distinctly enlarged (1).
699. Wulst, anteroposterior development in dorsal view: extends from olfactory bulb to cerebellum (0); positioned posteriorly (1); anteriorly positioned (2).
700. Wulst, lateral expansion of anterior wulst in dorsal view: not expanded laterally (0); expanded or swollen laterally beyond lateral margin of the telencephalon (1).
701. Junction between telencephalon and cerebellum in dorsal view: v-shaped (0); curved (1).
702. Posterior telencephalon, shape in dorsal view: rounded (0); pointed (1).
703. Contact between telencephalon and dorsal optic lobe in lateral view: straight (0); curved (1).
704. Relative size of optic lobe in lateral view (relative to rhombencephalon): small (0); large (1).
705. Optic lobe in dorsal view: visible (0); not visible (1).
706. Occipital sinus: not visible (0); clearly demarcated on dorsal cerebellum (1).
707. Dorsal projection of cerebellum in lateral view: <50% of posterior margin of telencephalon in contact with cerebellum (0); >50% of posterior margin of telencephalon in contact with cerebellum (1).
708. Relative length of telencephalon and cerebellum in lateral view: short cerebellum (0); long cerebellum (1).
709. Cerebellum, width in dorsal view: narrow (0); moderately wide (1); wide (2).
710. Cerebellum, fissures: distinct (0); indistinct (1).
711. Cerebellum, floccular lobes, mediolateral length: reduced (0); short (1); elongate (2).
712. Cerebellum, floccular lobes, fenestration: absent (0); present (1).
713. Cerebellum, floccular lobes, shape in posterior view: triangular with tapered lateral margin (0); rectangular with thickened dorsal and ventral margins (1).
714. Rhombencephalon, sulcus: conspicuous (0); inconspicuous (1).
715. Pituitary gland, contact with optic tract: separated (0); contacting (1).
716. Carotid artery, entrance into pituitary gland fossa: single (0); paired (1).
717. Brain and endosseous labyrinth, relative position and orientation: vertical long-axis (0); horizontal long-axis (1).
718. Semicircular canal, cross-sectional shape in lateral view: rounded (0); compressed (1).
719. Semicircular canals, number of common crus: two (asc-psc and hsc-psc) (0); three (asc-psc, hsc-psc, and asc-hsc) (1).
720. Endosseous labyrinth, cochlear curvature in lateral view: curved (0); straight (1).
721. Endosseous labyrinth, shape of distal cochlea in lateral view: tapered (0); swollen (1).
Table A1.
Optimized synapomorphies of selected groups recovered through constrained analyses employing the molecular backbone constraints based on the phylogenies of Kimball et al [
66], Reddy et al. [
65] and Prum et al. [
64]. Synapomorphies with a CI of 1.0 are noted with a subscript 1. Asterisks indicate unambiguous synapomorphies. Dashes indicate groups that were unresolved in strict consensus trees.
Table A1.
Optimized synapomorphies of selected groups recovered through constrained analyses employing the molecular backbone constraints based on the phylogenies of Kimball et al [
66], Reddy et al. [
65] and Prum et al. [
64]. Synapomorphies with a CI of 1.0 are noted with a subscript 1. Asterisks indicate unambiguous synapomorphies. Dashes indicate groups that were unresolved in strict consensus trees.
| Group Name | Kimball et al. [66] Constraint | Reddy et al. [65] Constraint | Prum et al. [64]
Constraint |
|---|
| Gruiformes + Charadriiformes | 1(1)*, 2(1), 27(1)*, 62(0), 74(1), 75(1)*, 92(2)*, 98(1)*, 125(1)*, 142(1), 154(1)*, 173(2)*1, 181(2), 188(1), 223(2)*, 229(2), 236(2), 292(1)*, 311(2)*, 326(1)*, 333(1), 334(2)*, 341(2)*, 345(2), 361(1), 369(2)*, 401(2), 406(2)*, 407(1), 436(2), 439(2), 443(2)*, 452(2), 453(1), 493(2), 494(2), 506(2), 518(1), 527(1), 546(1), 552(1)*, 565(2), 586(2)*, 606(1), 611(2), 623(3), 641(2)*, 673(2), 677(1), 679(2)*, 686(2), 693(1)*, 698(0), 699(2) | - | - |
| Charadriiformes (including S.mikkelseni and N. grandei) | 4(1), 16(2)*, 21(2)*, 46(1), 55(1), 68(1), 69(1), 71(2), 86(1)*, 91(2), 99(2)*, 108(0), 111(1), 129(1)*,163(0), 167(2), 180(2)*, 183(2)*, 206(2)*, 237(2), 253(2), 261(2)*, 270(0)*, 279(2)*, 299(1)*, 315(0), 317(0)*, 342(2)*, 346(2)*, 356(3), 359(1), 362(2), 366(2)*, 377(2)*, 393(0), 404(1)*, 449(3), 450(2)*, 451(1)*, 469(1), 470(2)*, 473(1), 489(2), 495(3), 498(2)*, 519(1), 527(3), 532(2)*, 535(1), 554(2), 555(2)*, 579(1), 595(2)*, 598(2)*, 610(2)*, 615(1), 643(2)*, 652(1)*, 671(2)*, 672(2), 691(2)*, 692(0), 719(1)*, 720(0)* | 11(3), 62(0), 71(2), 84(0), 90(2)*, 91(2), 92(2), 108(0), 120(1), 125(1), 129(1), 163(0), 173(2)*, 181(2), 183(2)*, 206(2)*, 223(2), 237(2), 238(1)*, 279(2), 299(1), 311(2), 317(0), 324(2), 326(1), 333(1), 334(2), 341(1), 345(2), 356(3), 359(1), 361(1), 366(2), 369(2)*, 377(2), 391(2), 393(0), 406(2), 407(1), 449(3), 450(2), 451(1), 453(1), 470(2), 479(2), 489(2), 493(2), 495(3), 498(2)*, 506(2), 521(1)*, 535(1), 552(1), 554(2), 584(1), 586(1)*, 595(2), 610(2), 641(2)*, 652(1), 668(3), 670(2)*, 672(2), 677(1)*, 679(2), 686(2), 692(0), 698(0), 707(0), 717(0), 719(1), 720(0), 721(1) | 1(1), 10(1)*, 15(2)*, 18(3)*, 20(1), 32(1), 44(1), 58(1)*, 61(2), 75(1), 86(1), 98(1)*, 125(1)*, 129(1), 144(1), 151(1), 155(1), 165(1), 173(2)*, 175(1), 176(1), 223(2), 248(2)*, 258(1)*, 262(1), 273(2)*, 279(2)*, 292(1)*, 324(2), 338(2), 340(1)*, 355(2)*, 356(3)*, 359(1)*, 377(2)*, 385(1), 400(1), 407(1), 437(1), 443(2), 448(1), 450(2)*, 451(1)*, 453(1)*, 454(2), 455(1), 468(1), 470(2)*, 477(2)*, 478(1)*, 487(2)*, 489(2)*, 497(2), 502(1), 517(1), 527(2)*, 540(1), 546(1)*, 547(1), 579(1), 583(1), 588(2), 592(2), 597(2), 598(2)*, 623(3), 632(2), 639(1), 651(1), 652(1), 662(3), 671(2), 679(2)*, 687(2), 719(1)*, 720(0) |
| S. mikkelseni + (Turnix + N. grandei) | 47(2), 87(1), 90(2)*, 103(1), 104(2), 120(2), 142(2), 188(2), 190(1), 229(1), 232(2), 292(2)*, 296(2), 297(2), 306(2), 319(1), 320(1), 341(1), 348(0), 375(1), 385(2), 395(1), 408(0), 411(1), 412(1), 416(2), 418(2), 419(2), 422(1), 425(2), 430(1), 436(1), 439(1), 442(2), 444(2), 445(1), 458(1), 464(1), 466(1), 472(2), 492(2), 521(1)*, 526(1), 539(2), 542(3), 549(1), 553(1)*, 558(2), 568(1), 579(3), 585(1), 589(2), 596(1), 600(2), 605(1), 621(2), 622(1), 626(0), 685(1), 717(0) | - | - |
| Turnix + N. grandei | 34(1)*, 91(1), 150(2), 155(2), 160(2), 192(2), 225(1), 245(2), 511(2), 513(1), 582(2), 586(1)* | - | - |
| Charadriiformes (excluding Scandiavis mikkelseni and Nahmavis grandei, excludes Turnix under Kimball et al. 2019 constraint) | 18(1)*, 78(2)*, 204(2)*, 216(2), 258(1)*, 272(0)*, 273(2)*, 280(2), 308(2)*, 356(4), 392(1), 396(1), 401(1), 413(2)*, 429(2)*, 454(2)*, 457(2), 467(1)*, 477(2)*, 478(1)*, 481(1), 487(2), 502(1)*, 510(2)*, 512(2), 518(2), 531(1), 543(2), 552(2), 570(1)*, 577(1), 584(2), 590(1)*, 591(2)*, 608(2)*, 632(2), 638(1), 688(3)* | 15(2), 18(1)*, 36(0), 40(1), 49(0), 78(2), 150(2), 151(1)*, 154(1)*, 225(1), 227(1)*, 242(2), 292(0), 565(2)*, 582(2), 611(2)* | - |
| Crown Charadriiformes (Most exclusive clade containing fossil humeri) | 21(1), 51(1), 52(1)*,87(3)*, 98(2)*, 108(1)*, 216(1), 222(0), 237(1), 248(2)*,280(1), 299(2), 323(2)*, 336(2)*, 341(3)*, 342(1), 345(1)*, 346(1), 355(2)*, 356(3), 392(0), 401(2), 418(3), 436(1), 449(1), 481(2), 494(1), 540(2)*, 555(1)*, 558(2)*, 568(1), 592(2), 609(2)*, 610(1)*, 611(1), 641(1), 643(1)*, 650(1), 653(1)*, 663(2), 667(0), 672(1), 680(2)* | 42(1)*, 43(1)*, 47(1), 50(2), 71(1), 90(1)*, 136(2), 146(1), 148(2)*, 223(2), 227(2)*, 280(1), 313(1)*, 339(2)*, 347(0), 369(1)*, 370(1), 416(1), 483(2)*, 507(2), 519(2)*, 565(2), 636(1)*, 693(2), 699(0), 716(1), 717(1)*, 721(0)
- | - |
| SMF av 619 + (Chionis alba + IGM 100/1435) | 20(2), 21(2), 24(4), 31(2), 32(1), 34(1), 43(1), 44(3), 50(3), 53(1), 56(2), 66(1), 69(3), 75(2), 77(1), 85(2), 86(2), 87(4), 99(1), 112(2), 116(1), 122(2), 124(2), 158(1), 163(1), 165(2), 177(1), 179(2), 183(1), 204(1), 215(2), 216(2), 223(1), 239(1), 242(2), 246(1), 249(3), 273(1), 282(3), 285(1), 299(1), 312(2), 313(1), 317(1), 329(2), 333(2), 334(3), 342(2), 371(1), 385(3), 406(1), 418(1), 427(2), 443(1), 455(2), 461(1), 469(2), 474(2)*, 479(1), 487(1), 494(3), 501(1), 504(2), 506(1), 507(2), 514(1), 530(2), 531(2), 572(2), 573(1), 575(2), 577(2), 581(1), 583(1), 584(1), 588(1), 589(2), 590(3), 592(1), 599(1), 601(2), 608(1), 615(1), 638(2), 652(2), 664(2), 672(2), 677(0), 683(2), 691(1) | - | 20(2), 24(4), 31(2), 32(1), 34(1), 50(3), 53(1), 56(2), 58(1), 66(1), 69(3), 75(2), 77(1), 78(2), 85(2), 86(2), 87(4), 99(1), 116(1), 140(1), 148(1), 158(1), 177(1), 179(2), 183(1), 204(1), 223(1), 242(2), 260(1), 273(1), 285(1), 299(1), 312(2), 317(1), 329(2), 333(2), 334(3), 342(2), 369(2), 370(2), 371(1), 379(1), 406(1), 418(1), 425(1), 427(2), 443(1), 455(2), 461(1), 469(2), 474(2)*, 479(1), 483(1), 493(2), 501(1), 504(2), 506(1), 514(1), 530(2), 531(2), 572(2), 577(2), 581(1), 583(1), 584(1), 589(2), 590(3), 592(1), 599(1), 601(2), 608(1), 615(1), 632(2), 638(2), 652(2), 671(2), 672(2), 677(0) |
| Chionis alba + IGM 100/1435 | 475(1)* | 20(2), 24(4), 31(2), 32(1), 34(1), 44(3), 50(3), 53(1), 56(2), 58(1), 66(1), 69(3), 75(2), 77(1), 78(2), 85(2), 86(2), 87(4), 99(1), 116(1), 140(1), 148(1), 158(1), 177(1), 179(2), 183(1), 204(1), 216(2), 223(1), 242(2), 260(1), 273(1), 285(1), 299(1), 312(2), 317(1), 329(2), 333(2), 334(3), 342(2), 369(2), 370(2), 371(1), 379(1), 406(1), 418(1), 425(1), 427(2), 443(1), 455(2), 461(1), 469(2)*, 474(2)*, 475(1)*, 479(1), 483(1), 493(2), 494(3), 501(1), 504(2), 506(1), 514(1), 530(2), 531(2), 577(2), 581(1), 583(1), 584(1), 589(2), 590(3), 592(1), 599(1), 601(2), 608(1), 615(1), 632(2), 638(2), 652(2), 671(2), 672(2), 677(0), 683(2), 691(1), | 475(1)* |
| Gruiformes (including extinct taxa) | 34(1), 42(2), 49(0), 67(2)*, 77(1), 124(2), 131(2), 138(1), 197(3), 228(2), 289(1), 345(3), 350(2), 378(2)*, 379(2), 388(2)*, 423(0)*, 426(1), 433(1), 468(1), 475(1), 504(2)*, 506(3), 511(2)*, 530(2)*, 534(2)*, 537(2)*, 542(2)*, 561(1), 599(1), 624(1)*, 625(2), 636(2)*, 644(2), 661(2)*, 674(2)* | 2(1), 4(2)*, 16(1)*, 27(1), 32(1), 46(2), 49(0), 67(2)*, 74(1)*, 77(1), 84(0), 85(1), 87(3), 92(2), 112(1), 125(1)*, 126(1), 142(1)*, 174(1)*, 188(1)*, 197(3)*, 206(1), 210(1), 236(2)*, 246(2)*, 268(2), 289(1), 291(2), 292(1), 311(2)*, 325(1), 326(1), 328(2), 334(2), 341(2), 345(3), 378(2)*, 388(2)*, 392(0), 396(2)*, 401(2), 406(2), 408(1), 423(0)*, 426(1), 430(1), 433(1), 436(2), 437(1)*, 439(2), 442(3), 443(2), 456(2)*, 472(1), 504(2)*, 506(3)*, 516(2), 518(1), 520(2), 527(1)*, 530(2)*, 531(2)*, 534(2)*, 537(2)*, 546(1)*, 551(1), 553(2)*, 558(1), 561(1), 603(1), 606(1)*, 636(2)*, 641(2)*, 644(2), 646(1), 670(2)*, 679(2)*, 686(2) | 27(1), 36(1), 49(0), 67(2)*, 74(1)*, 124(2), 125(1)*, 131(2), 154(1), 173(2)*, 185(2), 188(1)*, 223(2)*, 227(1), 228(2)*, 229(2), 238(1), 269(2), 292(1), 311(2)*, 324(2), 326(1), 369(2)*, 388(2)*, 406(2), 426(1), 433(1), 436(2), 439(2), 504(2)*, 506(3), 514(1), 518(1)*, 527(1)*, 530(2)*, 534(2)*, 537(2)*, 542(2), 550(1)*, 551(1), 552(1)*, 558(1), 561(1)*, 584(1), 599(1)*, 624(1)*, 633(1), 641(2)*, 644(2), 674(2)*, 679(2)*, 686(2)*, 693(1)*, 721(1) |
Kimball et al. (2019): Ralloidea + (Messelornithidae + Songzia)
Reddy et al. (2017): Messelornithidae + (Songzia + Ralloidea)
Prum et al. (2015) Messelornithidae + Ralloidea (including Songzia)
| 65(1), 90(2)*, 123(2), 158(1), 257(2)*, 272(0)*, 297(2)*, 318(2), 338(2), 349(2), 351(1), 353(2), 355(2)*, 375(1), 385(2), 400(0), 413(2), 421(1), 430(1), 446(3), 454(2), 467(1)*, 474(2), 485(2), 490(2), 494(3), 515(2), 521(4), 523(2), 569(1), 590(1), 596(1), 622(1)*, 623(2), 645(2), 652(3), 655(2), 660(2)*, 662(3), 664(2) | 1(1), 13(2), 65(1), 90(2), 123(2), 181(2), 237(2), 250(2), 254(1), 257(2)*, 266(0), 272(0)*, 297(2)*, 318(2), 323(1), 338(2), 339(1)*, 344(1), 351(1), 353(2), 356(2)*, 375(1), 413(2), 421(1), 446(3)*, 461(2), 467(1)*, 490(2), 493(2)*, 513(2), 515(2), 521(4), 523(2), 560(1), 570(1), 572(2), 575(1), 590(1), 611(2), 622(1)*, 623(2), 645(2), 652(3), 662(3), 664(2) | 1(1), 2(1)*, 30(1)*, 46(1), 55(1), 65(1), 90(2)*, 91(2), 123(2), 181(2), 195(2), 208(1)*, 215(2), 218(1)*, 237(2), 253(2), 257(2)*, 258(1), 272(0)*, 297(2)*, 318(2), 338(2), 349(2), 353(2), 355(2), 356(2)*, 371(1), 430(1), 446(3)*, 453(1)*, 454(2), 467(1)*, 490(2), 493(2)*, 505(2), 510(2), 521(3), 523(2), 540(2), 545(1), 560(1), 565(2), 570(1), 572(2), 579(3), 611(2), 622(1), 623(2), 645(2), 652(3), 655(2)*, 656(2), 660(2), 662(3), 685(1) |
| Messelornithidae + S. acutunguis | 10(1), 13(2)*, 55(1), 62(1), 180(2), 225(1), 261(2), 279(2), 313(1)*, 334(1), 340(1)*, 344(1)*, 366(2), 407(0), 436(1), 441(2), 498(2), 500(3), 507(2), 512(2), 524(1), 540(2), 554(2), 555(2), 608(2), 609(2), 650(1), 671(1) | - | - |
| Crown Ralloidea (including S. acutunguis under latter two constraints) | 64(2)*, 66(1)*, 91(2)*, 138(2), 293(2), 308(2), 347(1), 360(2)*, 416(2), 426(2), 450(2)*, 561(2), 613(2)*, 651(2), 677(0) | 34(1)*, 62(0), 64(2), 138(2), 293(2), 308(2), 329(2), 360(2), 361(1), 362(3), 407(1), 422(1)*, 426(2), 450(2), 541(2), 550(0)*, 551(0), 561(2), 613(2), 651(2), 671(3) | 34(1), 62(0)*, 64(2)*, 66(1)*, 138(2), 159(2), 308(2)*, 334(3)*, 347(1)*, 360(2)*, 362(3), 396(1), 407(1)*, 425(2), 426(2), 450(2)*, 551(0), 613(2)*, 620(1)*, 649(1), 651(2)*, 654(1) |
| Messelornithidae | 34(2), 361(3), 371(1), 542(3), 543(2), 550(1)*, 551(1)*, 691(2) | 10(1)*, 55(1)*, 66(2), 180(2)*, 225(1)*, 261(2)*, 262(1)*, 279(2)*, 334(1), 366(2), 371(1), 436(1), 441(2), 498(2)*, 500(3), 507(2)*, 512(2), 524(1), 540(2), 542(3), 543(2), 554(2), 555(2)*, 608(2), 609(2), 650(1)*, 677(1), 691(2) | 10(1)*, 13(2), 68(2), 127(1), 180(2)*, 225(1)*, 261(2)*, 279(2)*, 313(1), 340(1)*, 344(1)*, 361(3), 366(2), 436(1), 441(2), 498(2)*, 500(3), 507(2)*, 512(2), 524(1), 542(3), 554(2), 555(2)*, 608(2), 609(2), 650(1)*, 671(1)*, 677(1), 691(2) |




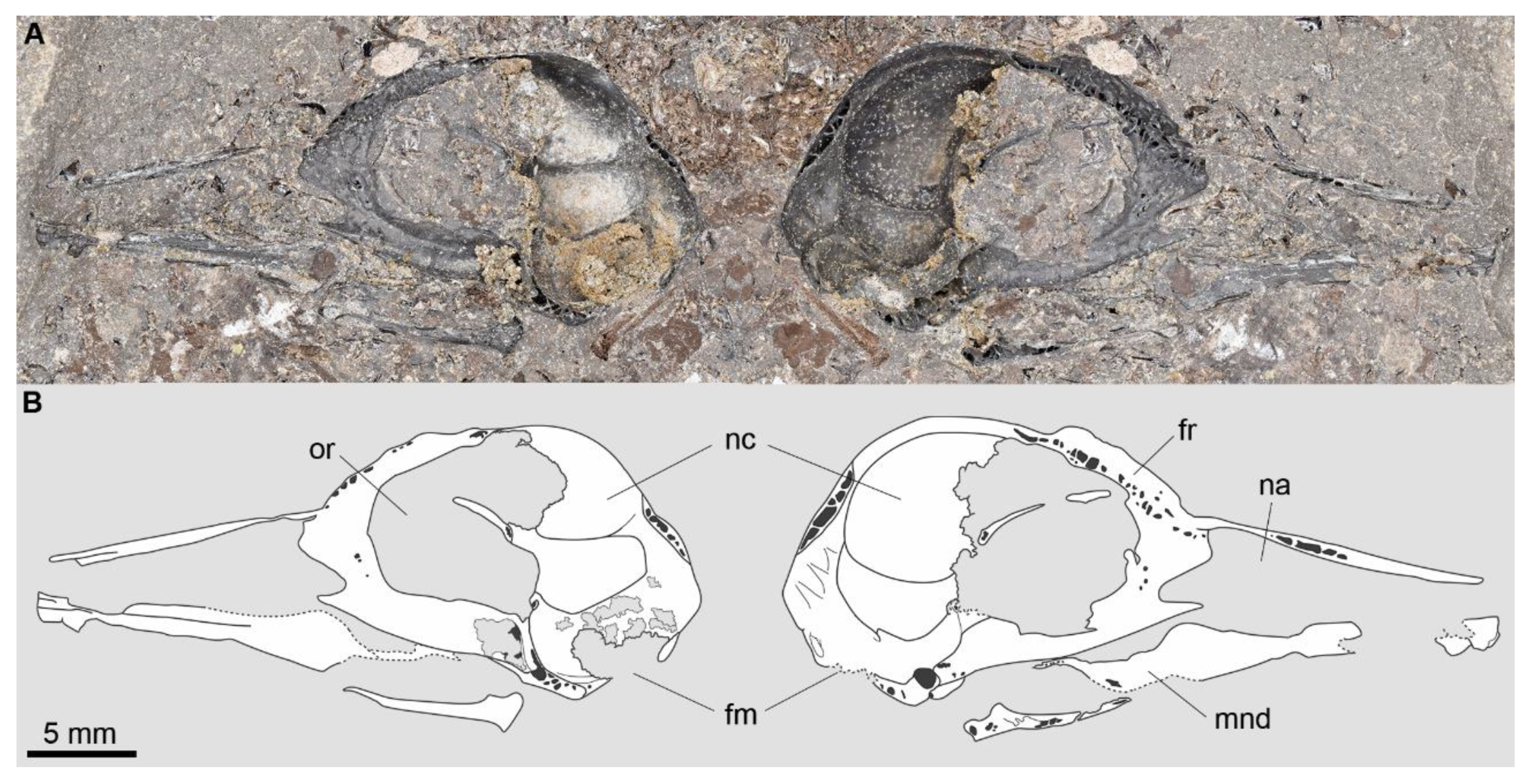{kind=link}
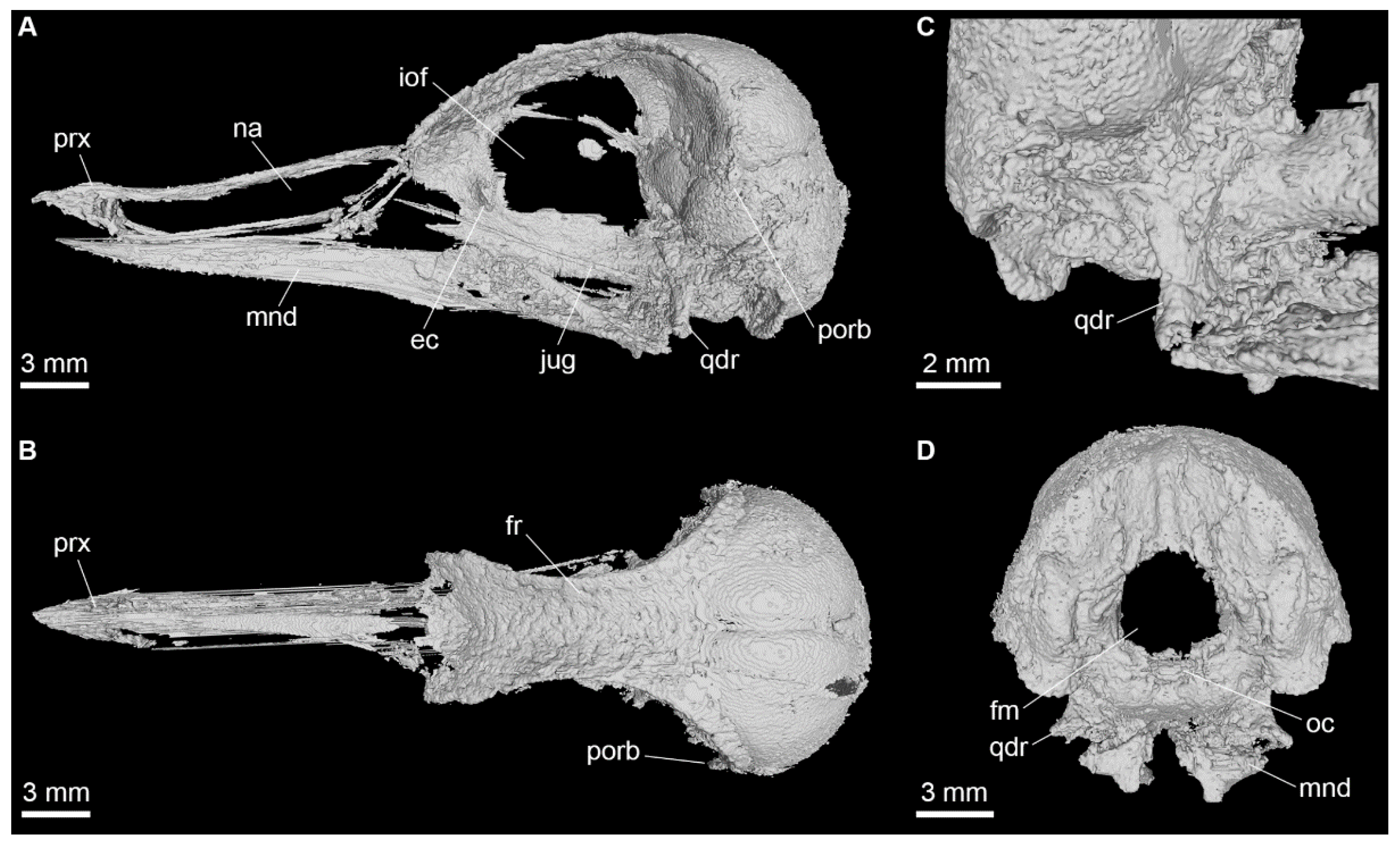{kind=link}
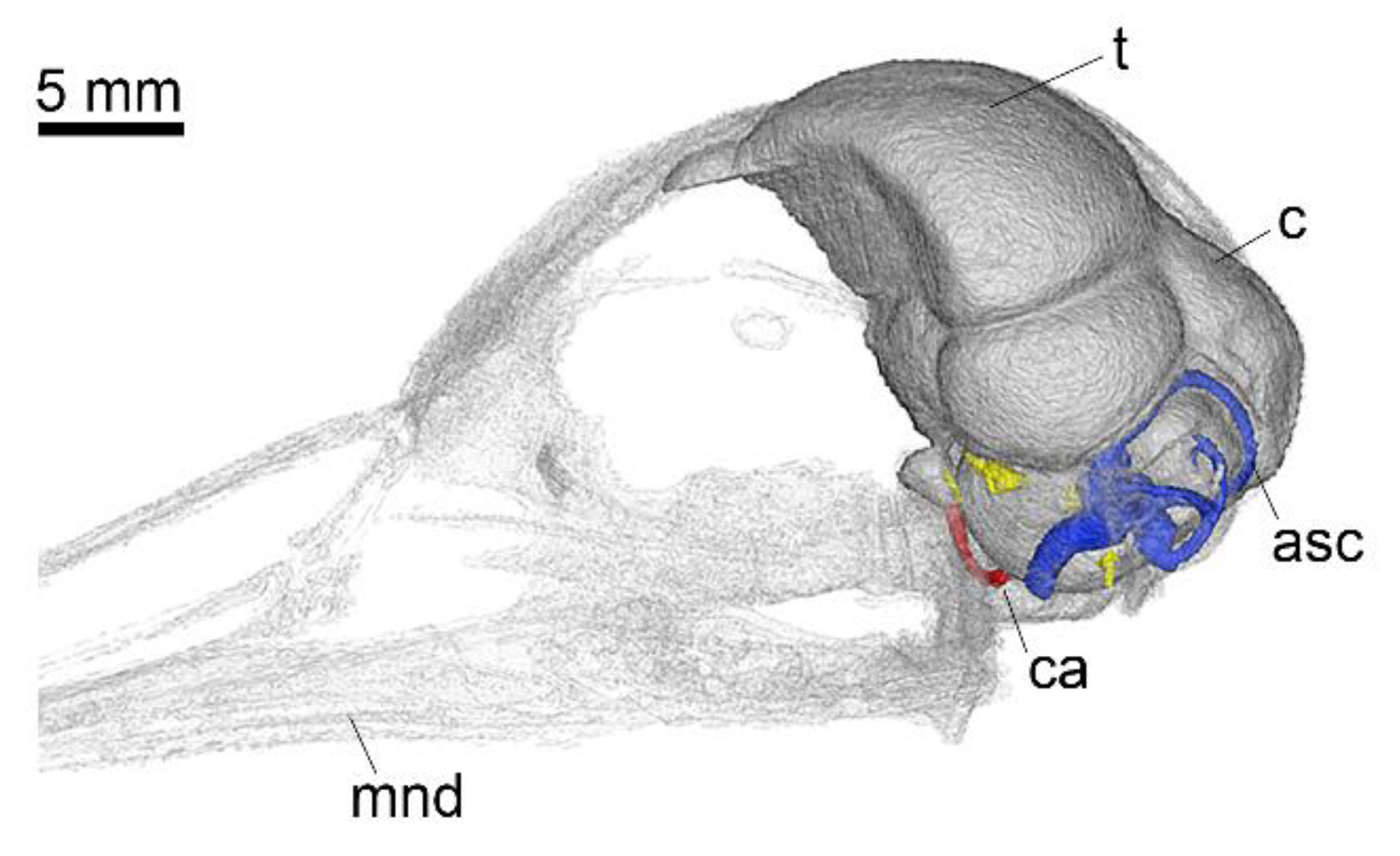{kind=link}
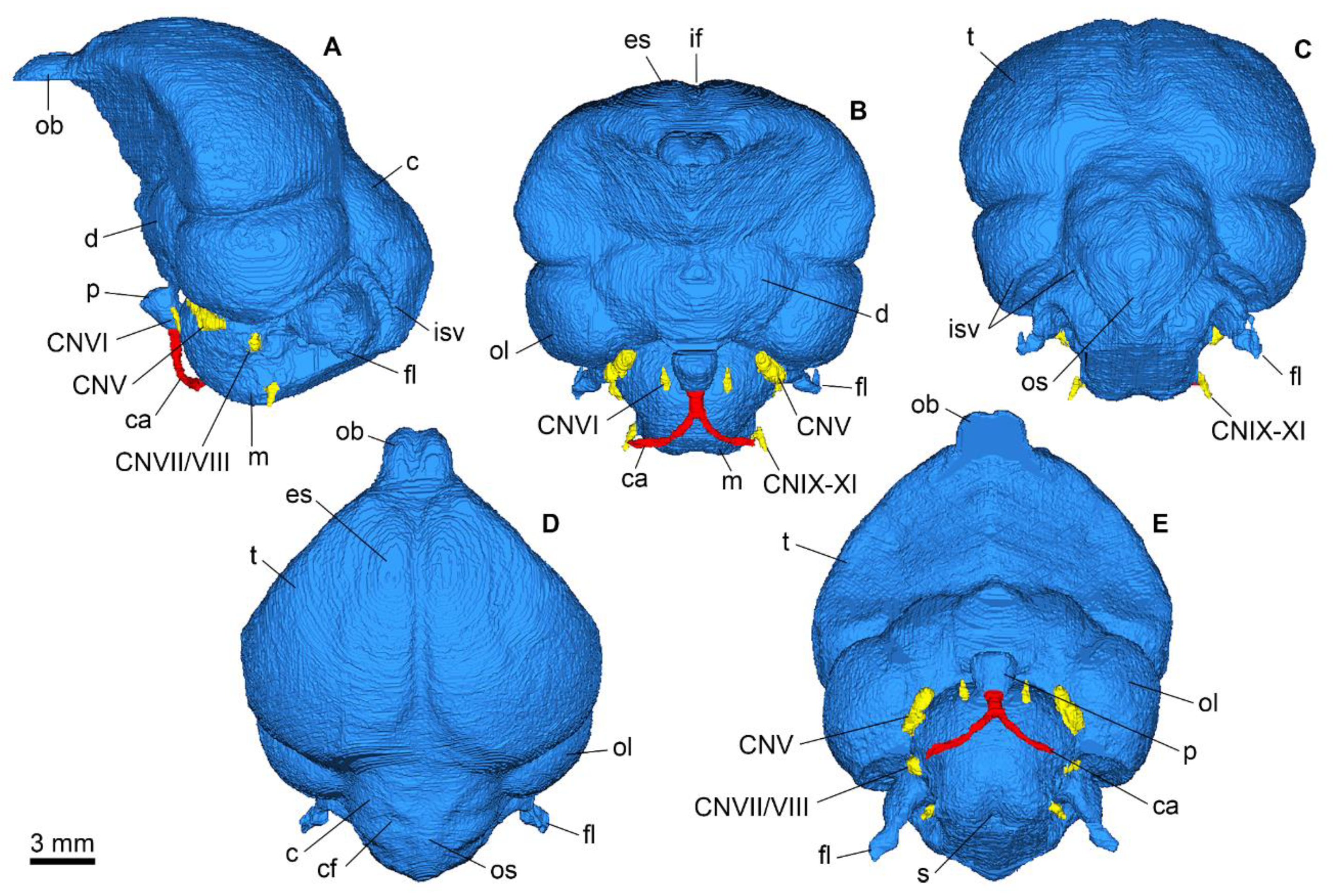{kind=link}
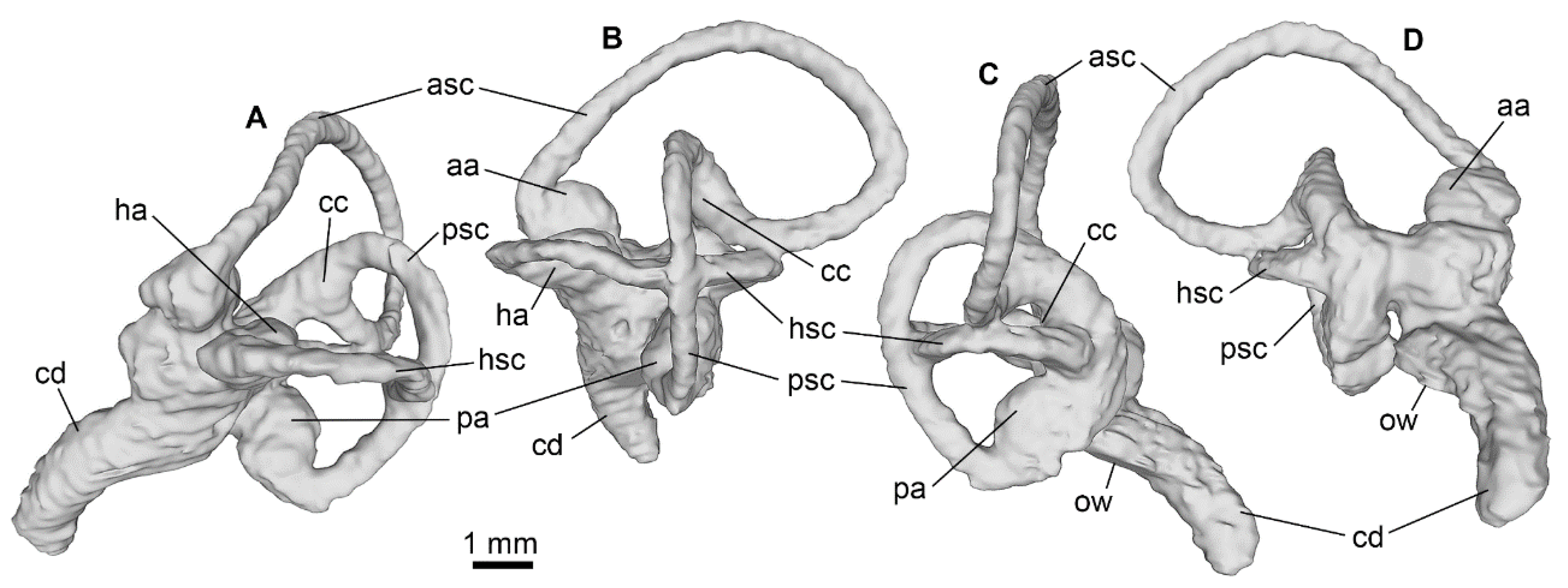{kind=link}
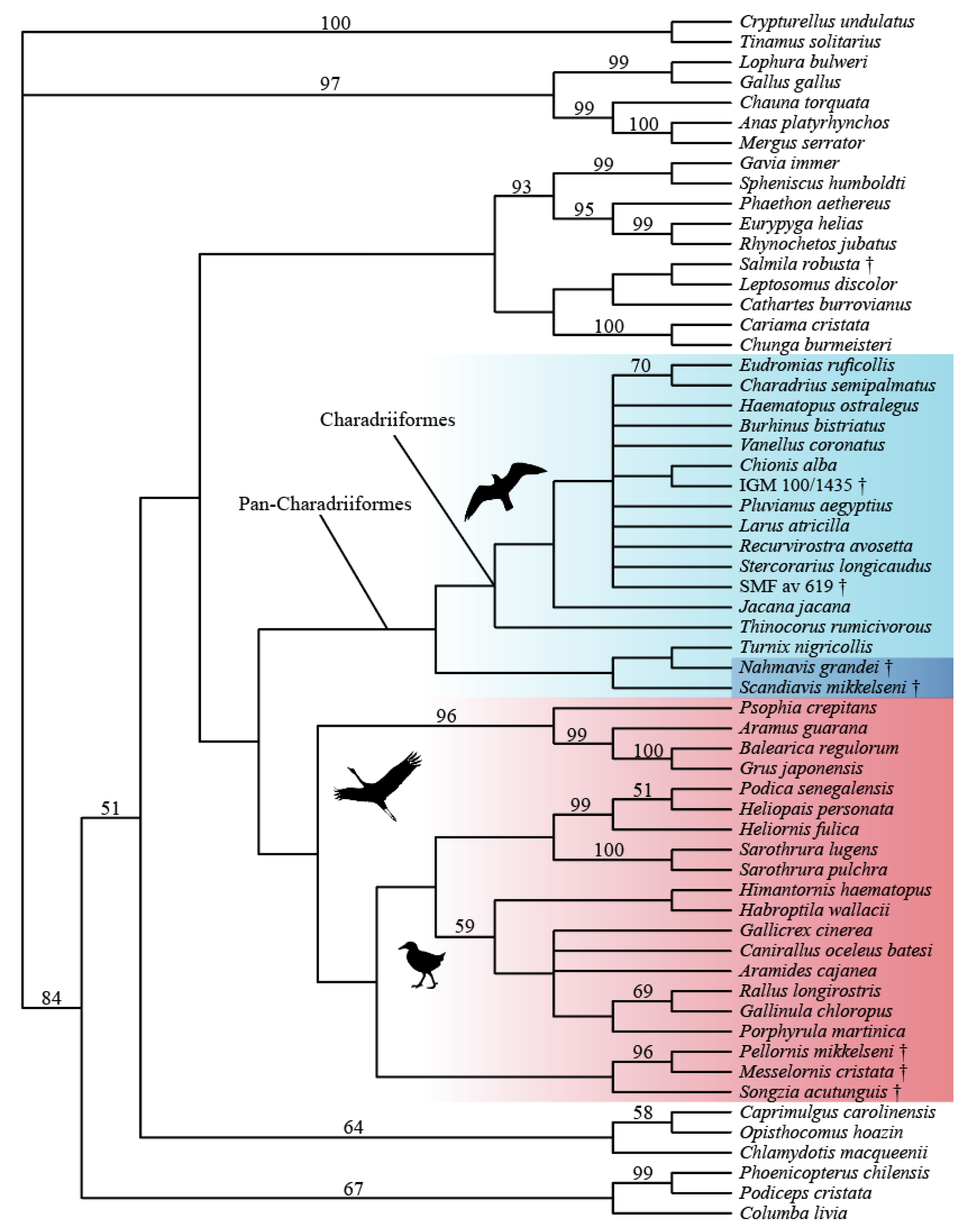{kind=link}
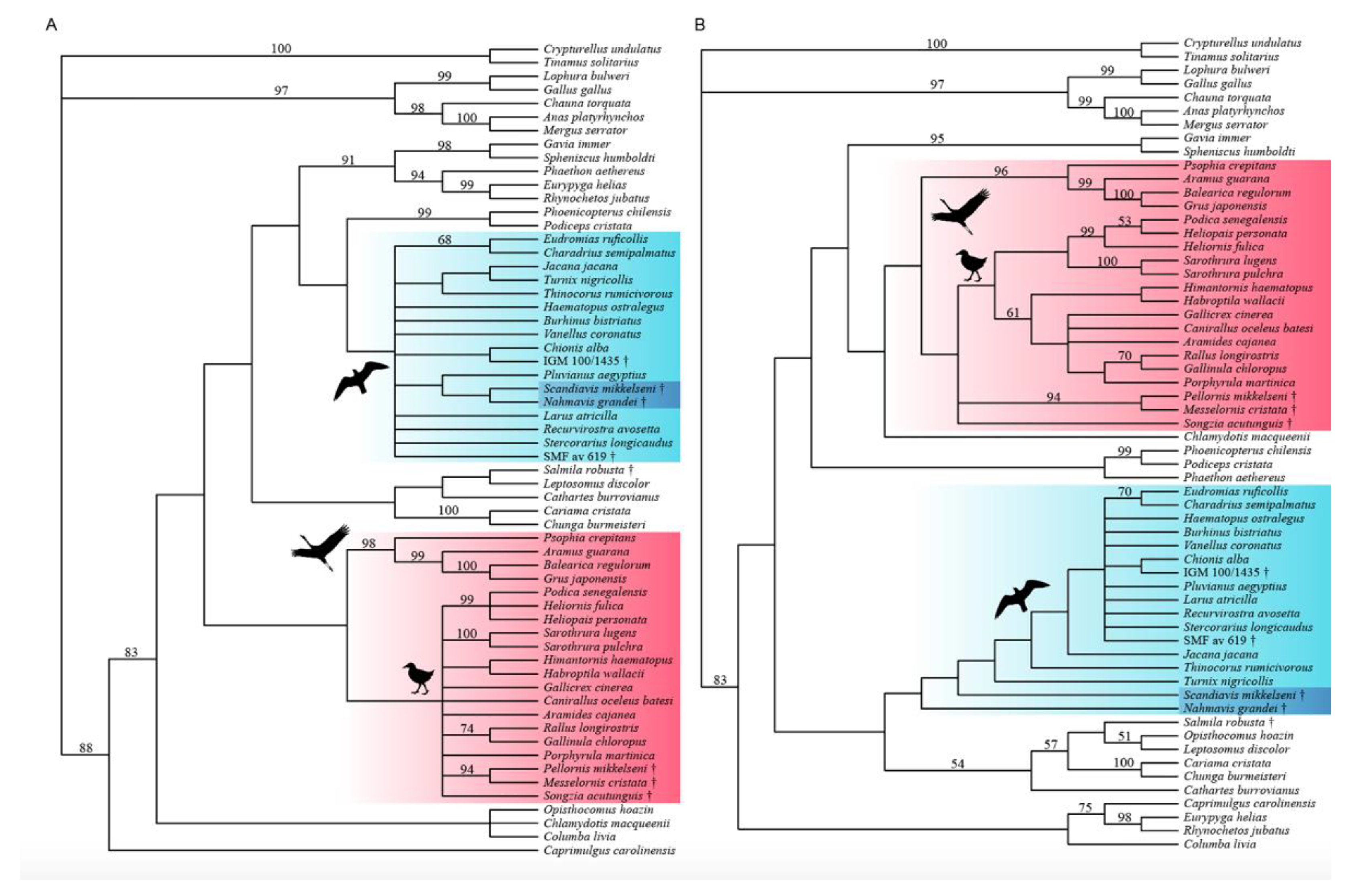{kind=link}