
Taxonomic Revision of Eastern Part of Western Palaearctic Cordulegaster Using Molecular Phylogeny and Morphology, with the Description of Two New Species (Odonata: Anisoptera: Cordulegastridae) †

 , ,
, , 
Abstract
:1. Introduction
1.1. General Remarks on the Genus Cordulegaster
1.2. Threats to Western Palaearctic Cordulegaster
1.3. History of the Taxonomy of Western Palaearctic Cordulegaster
1.4. Geological and Climatic Events Influencing Diversification of Western Palaearctic Cordulegaster
2. Material and Methods
2.1. Material
2.2. General Methods
2.3. Molecular Genetic Analysis
2.4. DNA Extraction, PCR Methods, and Sequencing
2.5. Phylogenetic Methods
MAFFT (Multiple Alignment with Fast Fourier Transform)
2.6. jModelTest
2.7. Tree Construction with MrBayes and PAUP
2.8. StarBeast
2.9. BPP
- Running species delimitation:Settings were Speciesdelimitation = 1 0 2, Speciestree = 1, speciesmodelprior = 1, burnin = 16,000, samplefreq = 2, nsample = 500,000.
- Running Species tree Estimation:Settings were Speciesdelimitation = 0, Speciestree = 1, speciesmodelprior = 1, burnin = 16,000, samplefreq = 2, nsample = 500,000.
2.10. Timetree
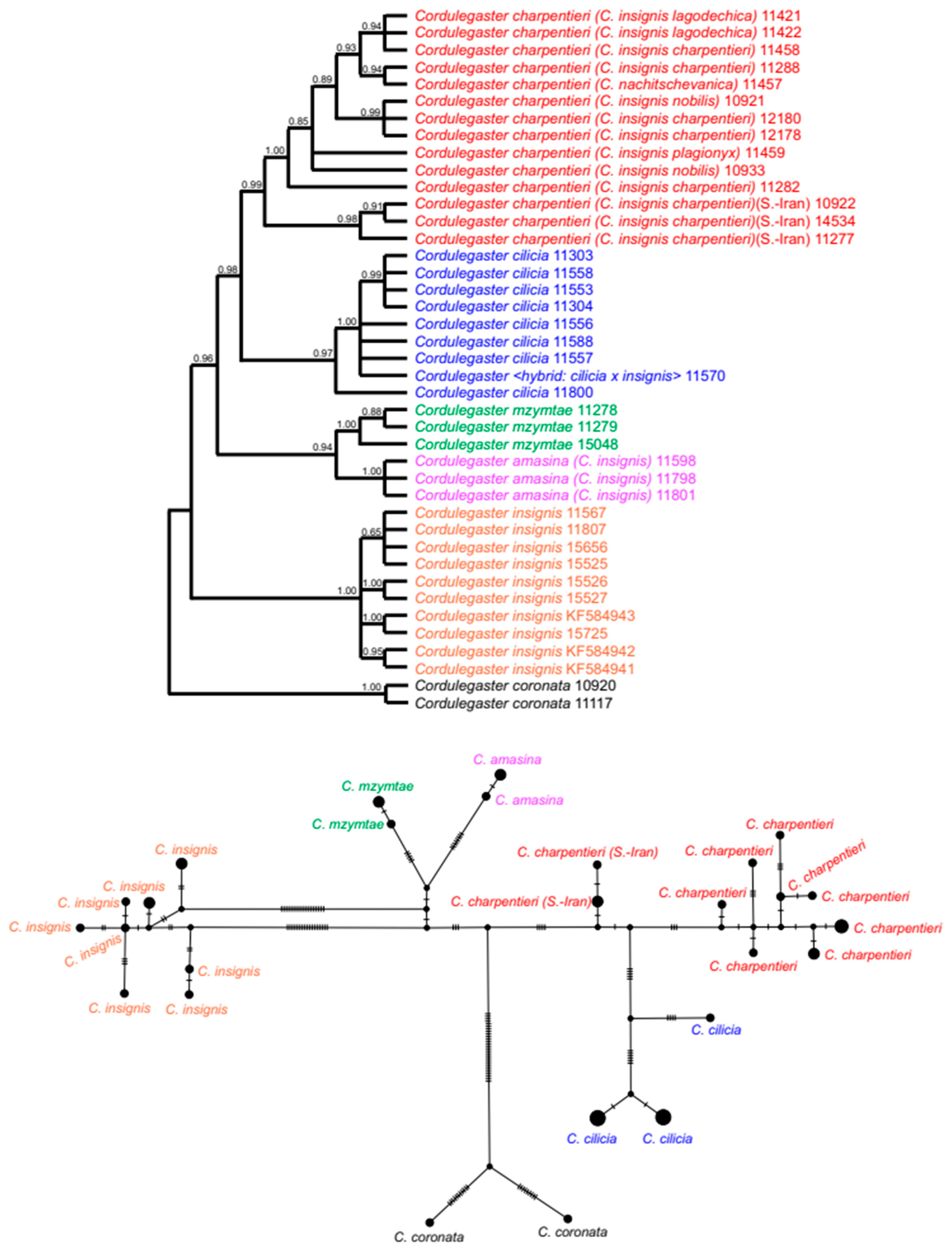
2.11. Haplotype Analysis
2.12. Evolutionary Distance Analysis
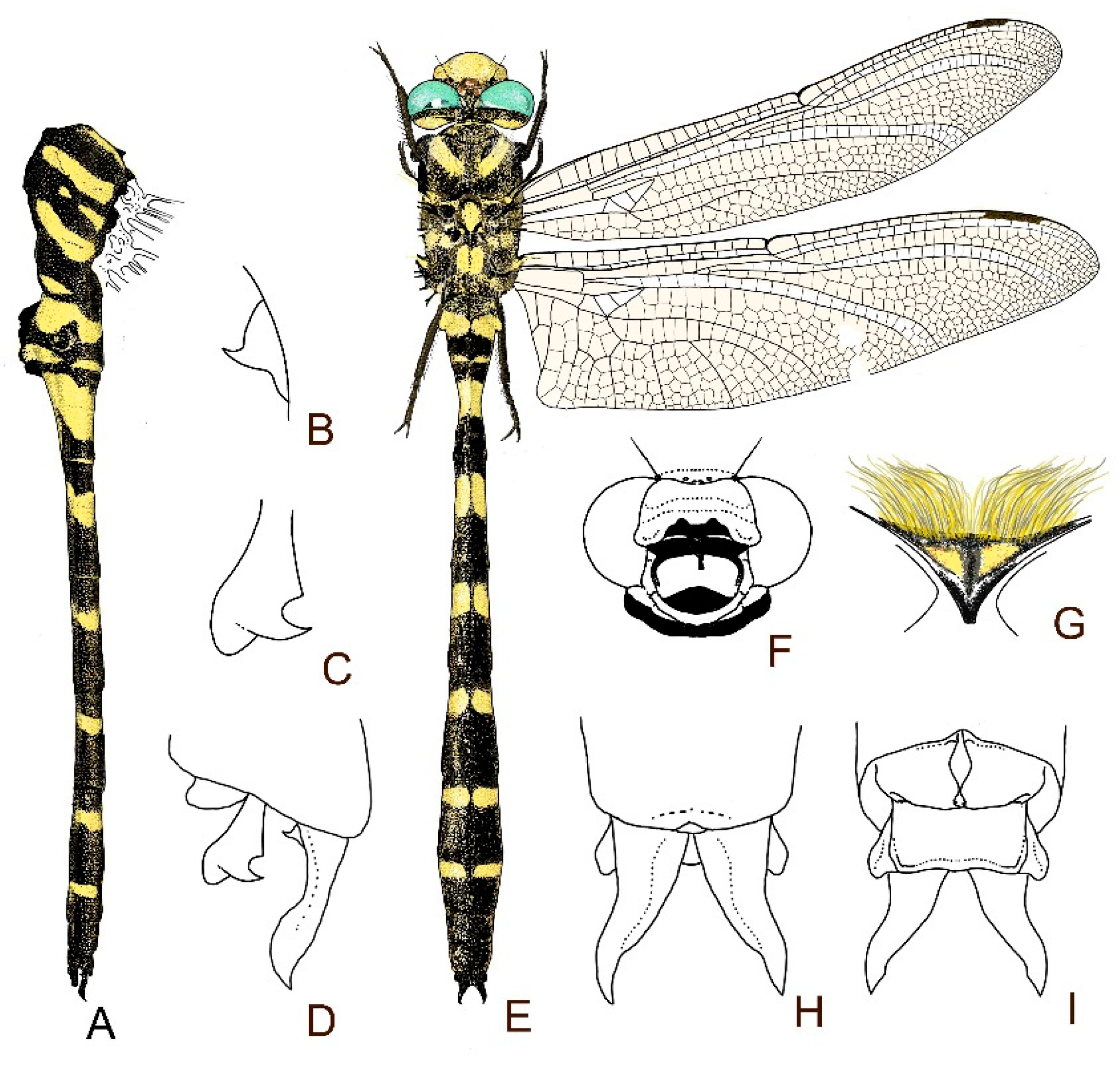
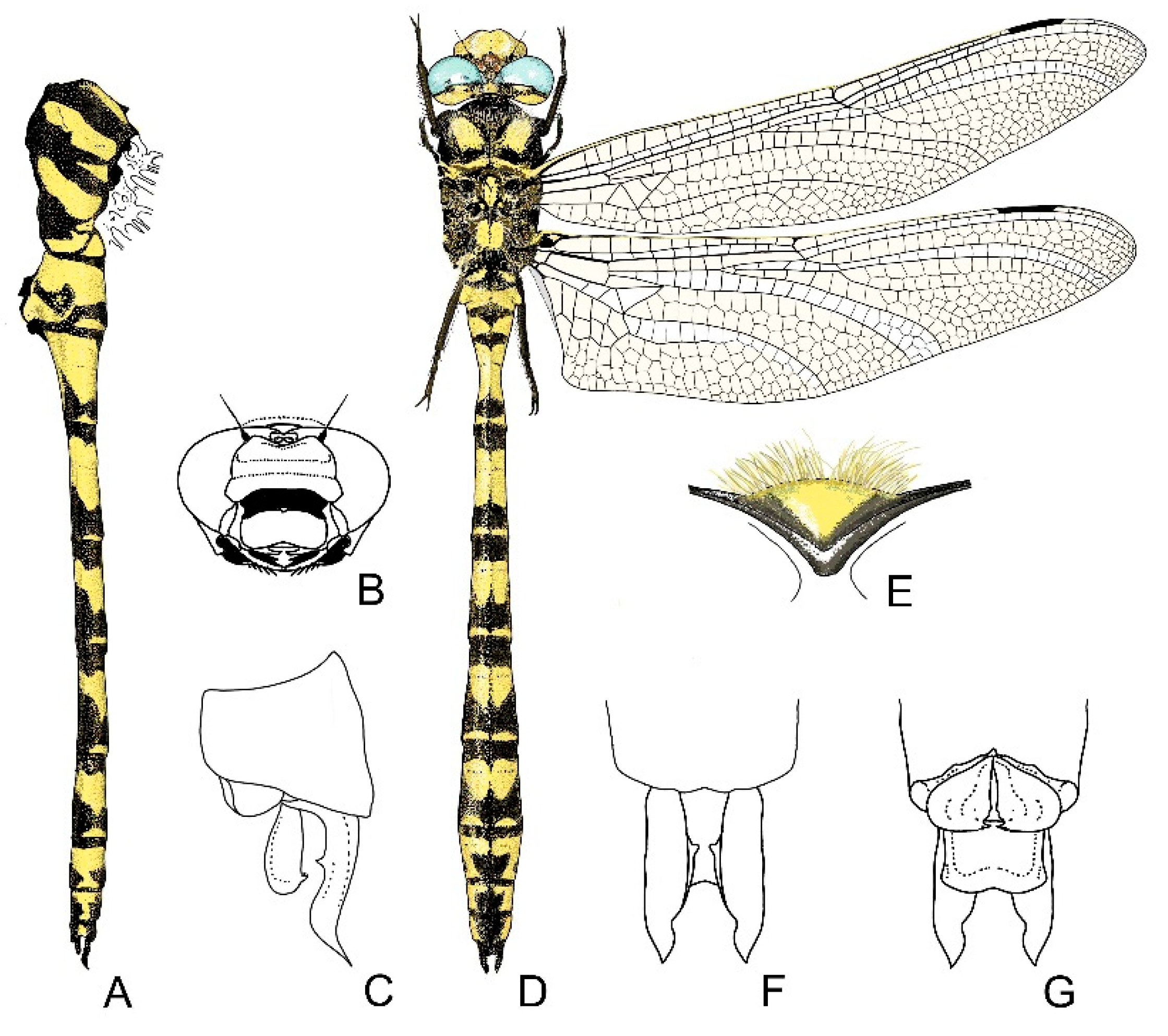
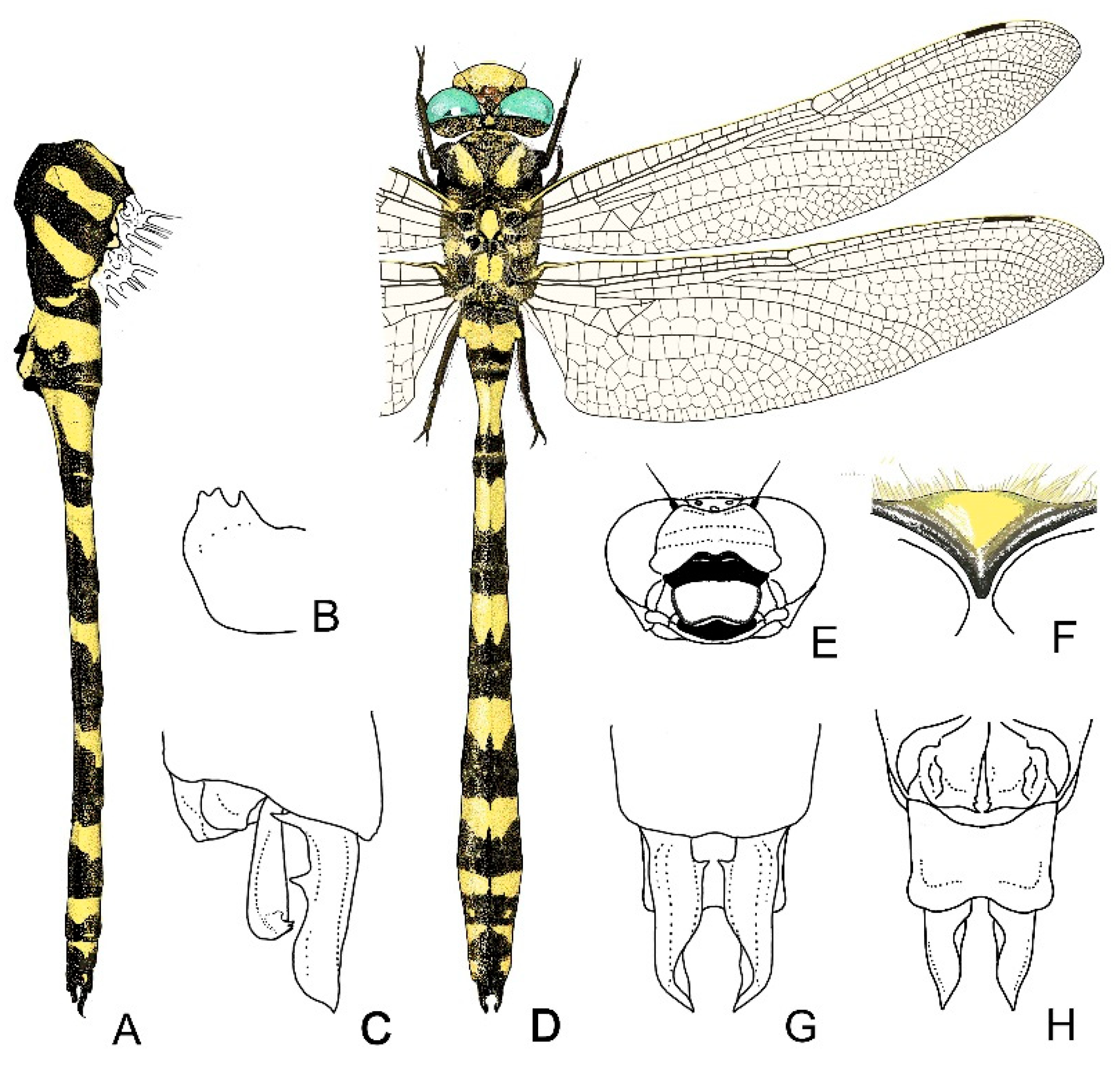
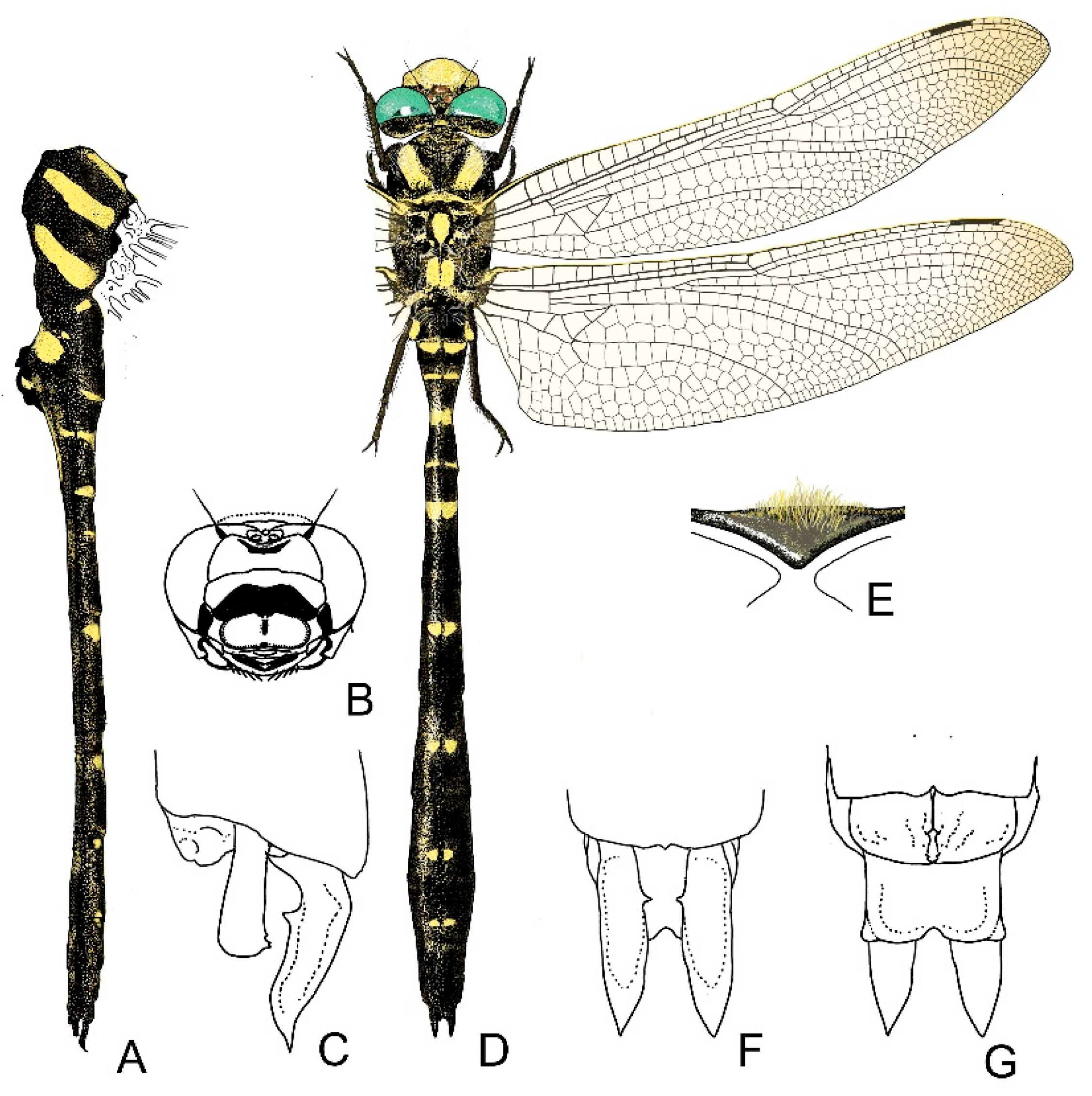
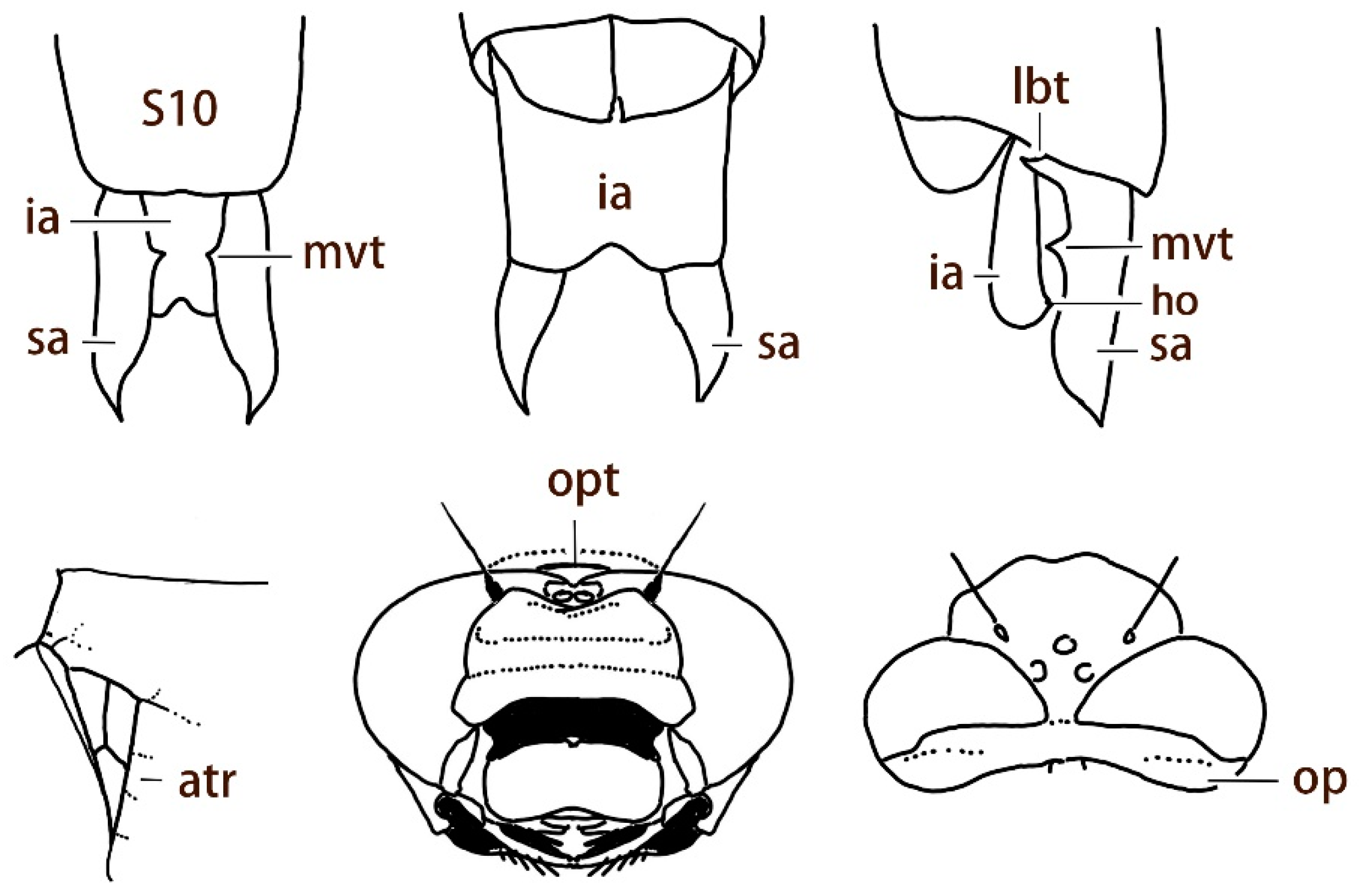
2.13. Morphological Analysis and Systematics
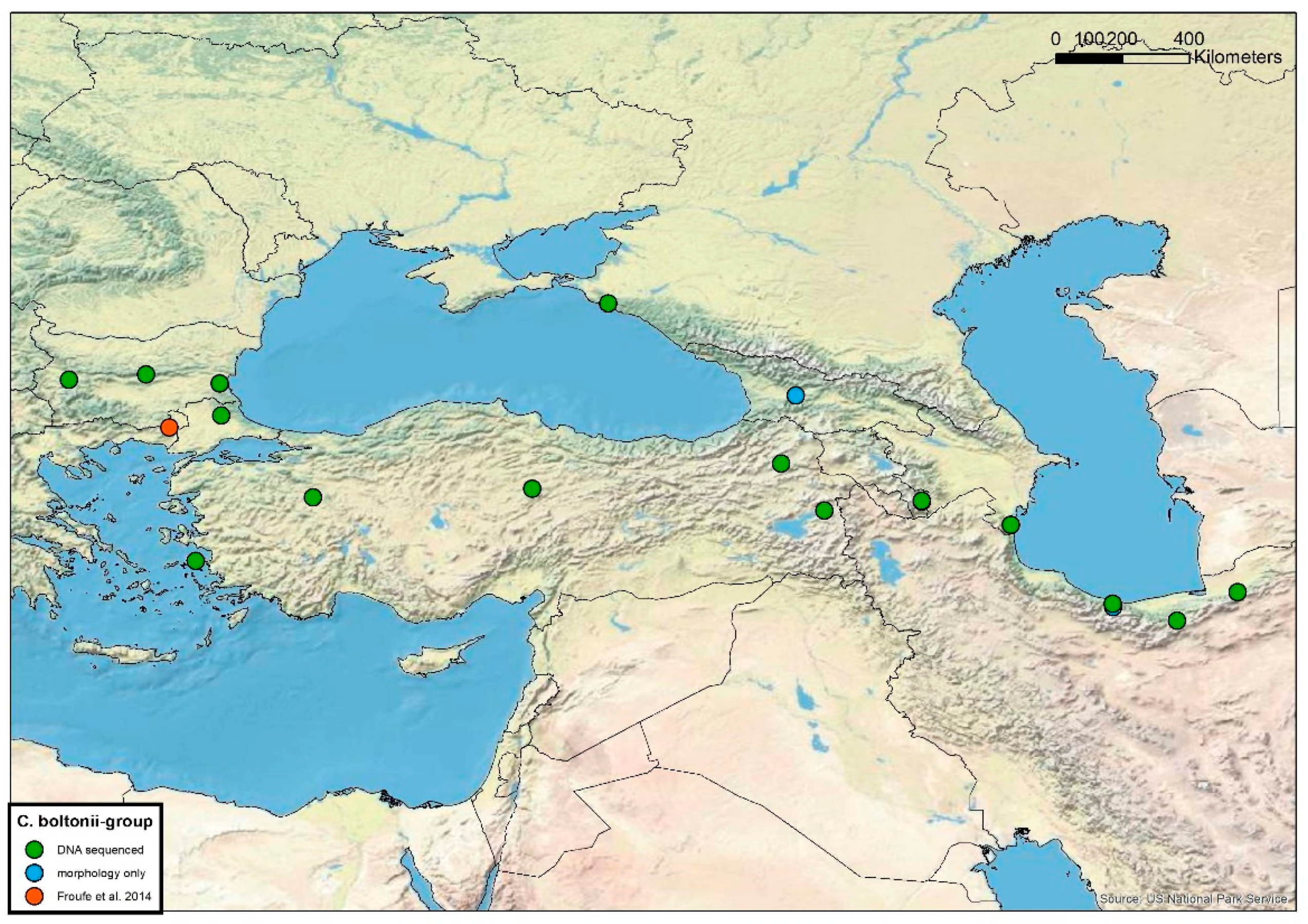
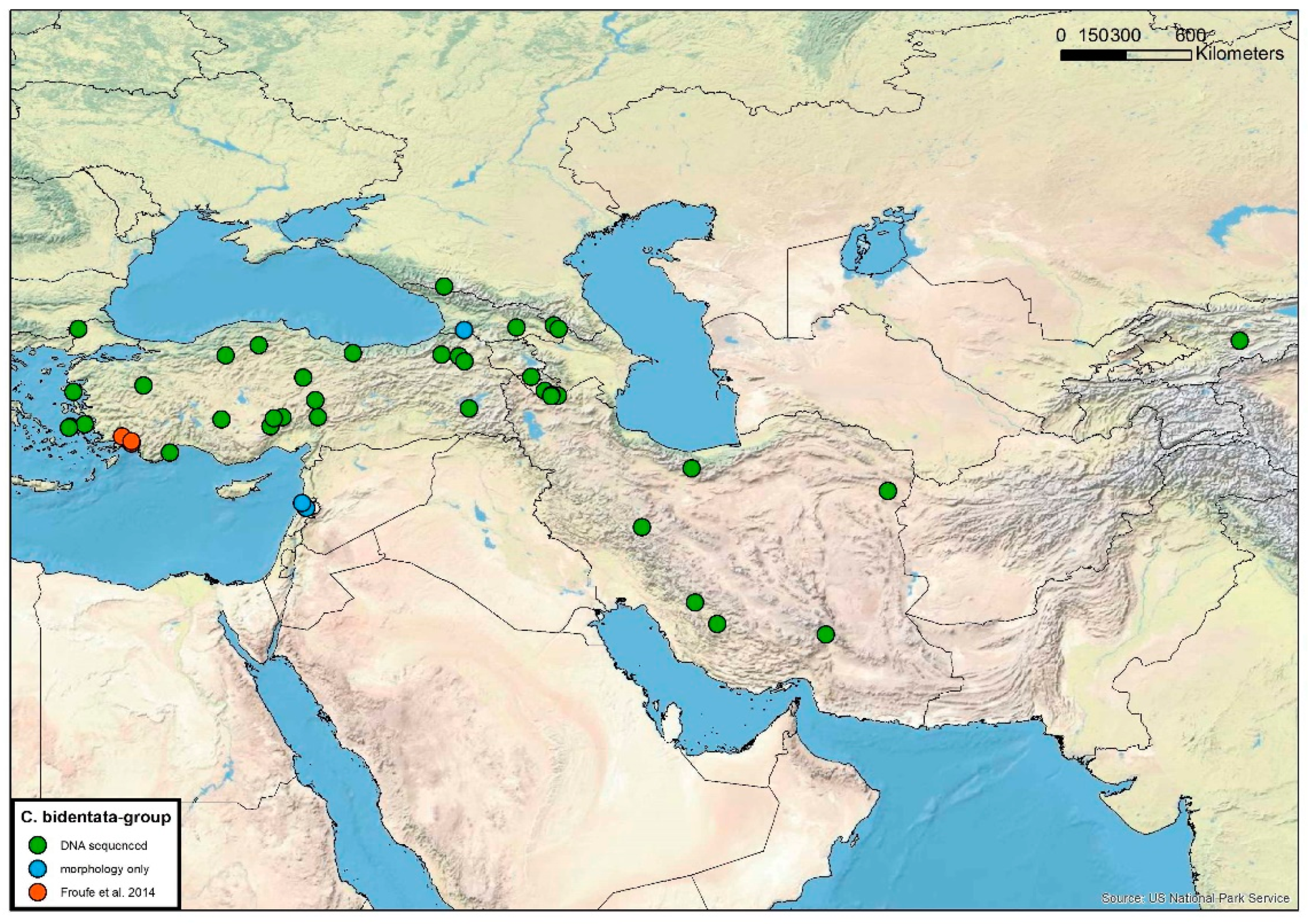
2.14. Distribution Maps
3. Results
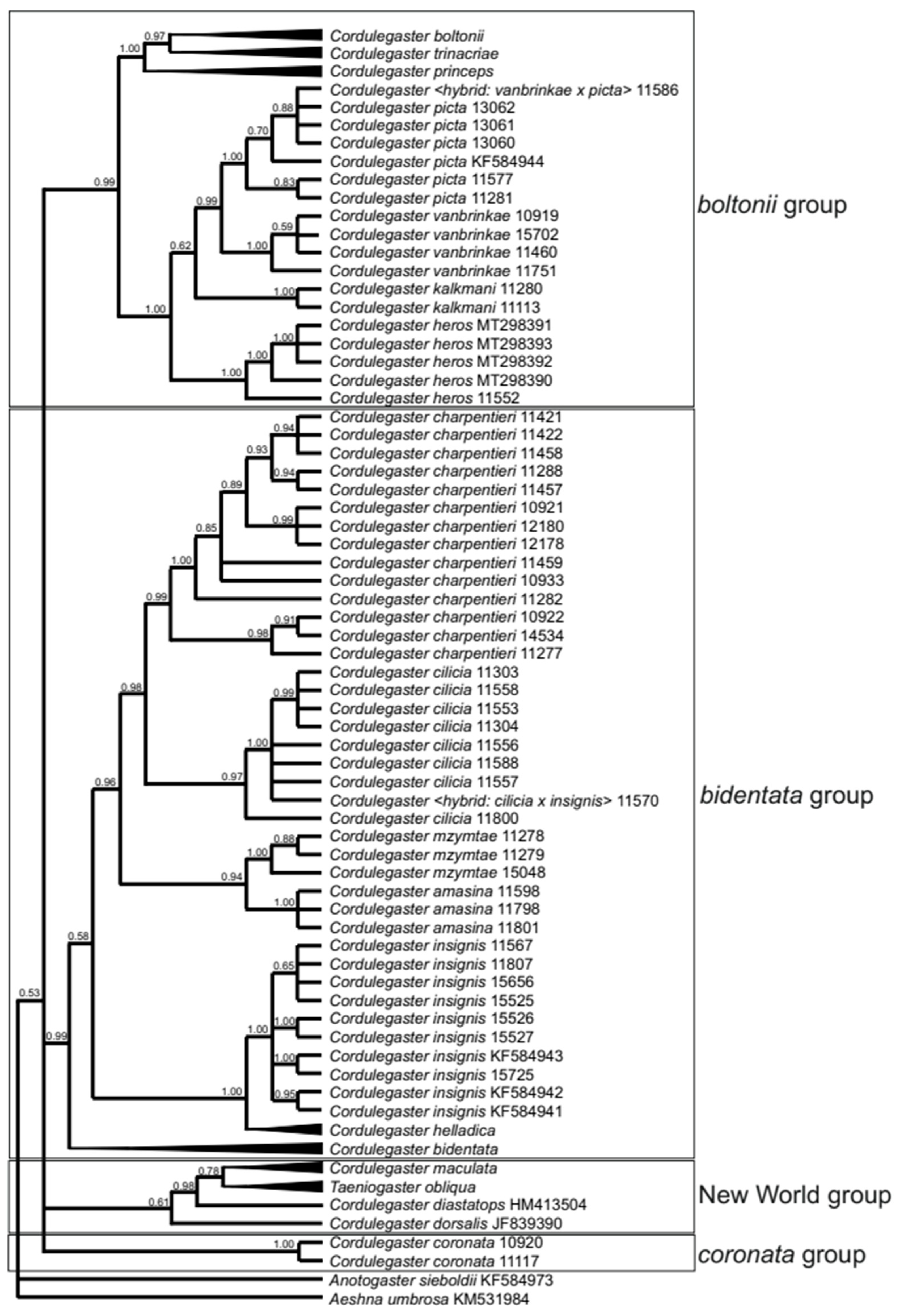
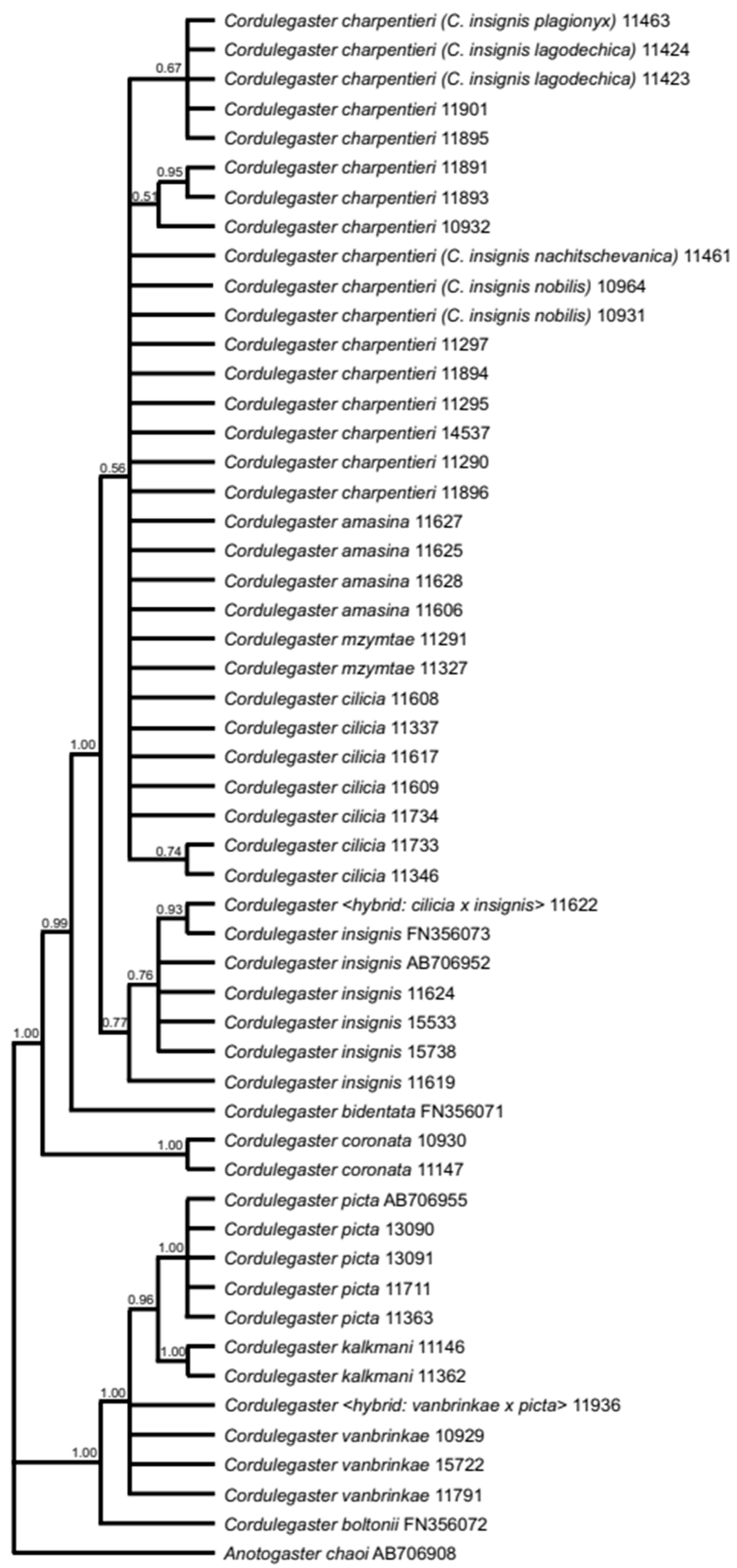
3.1. Molecular Genetic Analysis of the COI and ITS Genes
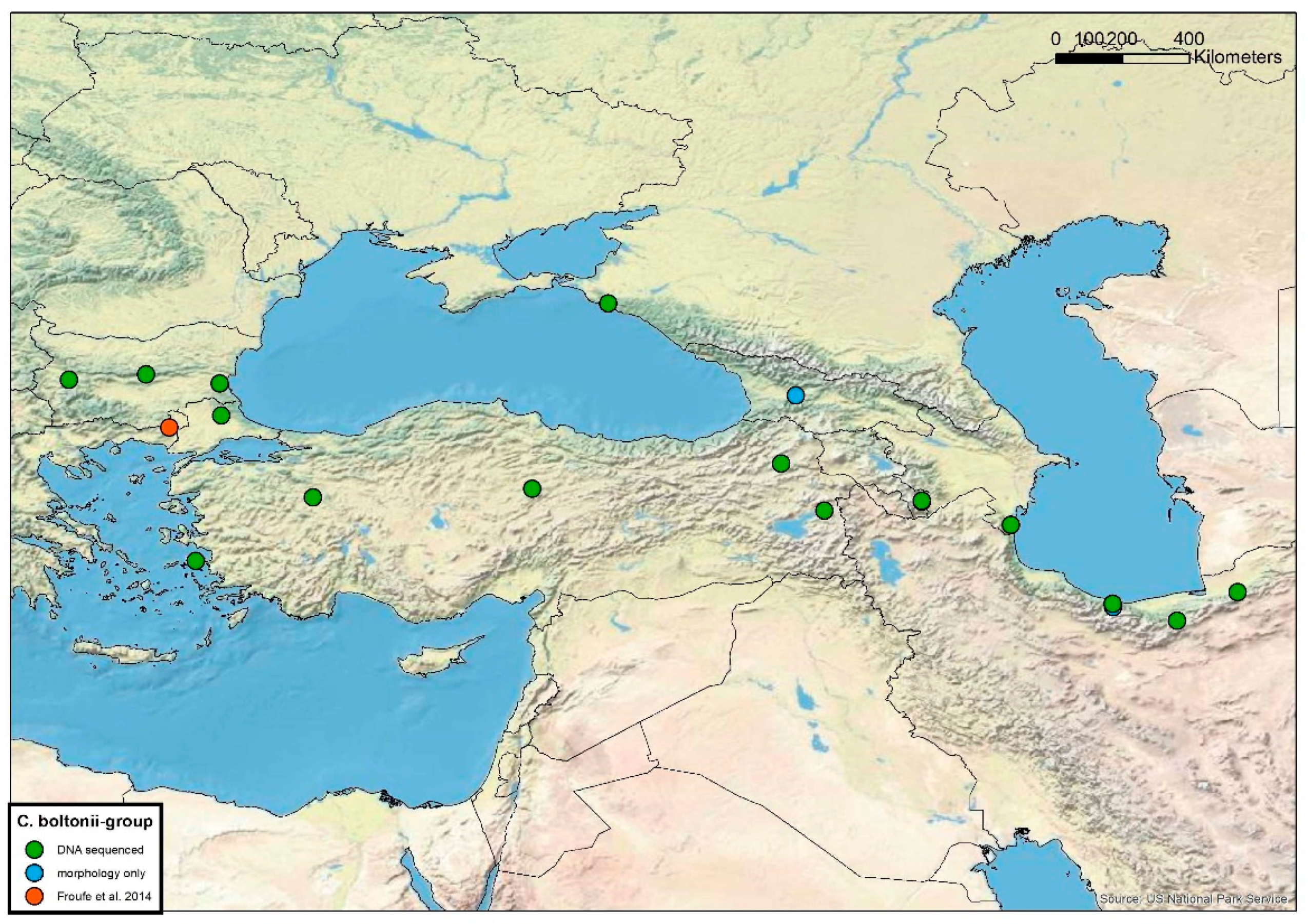
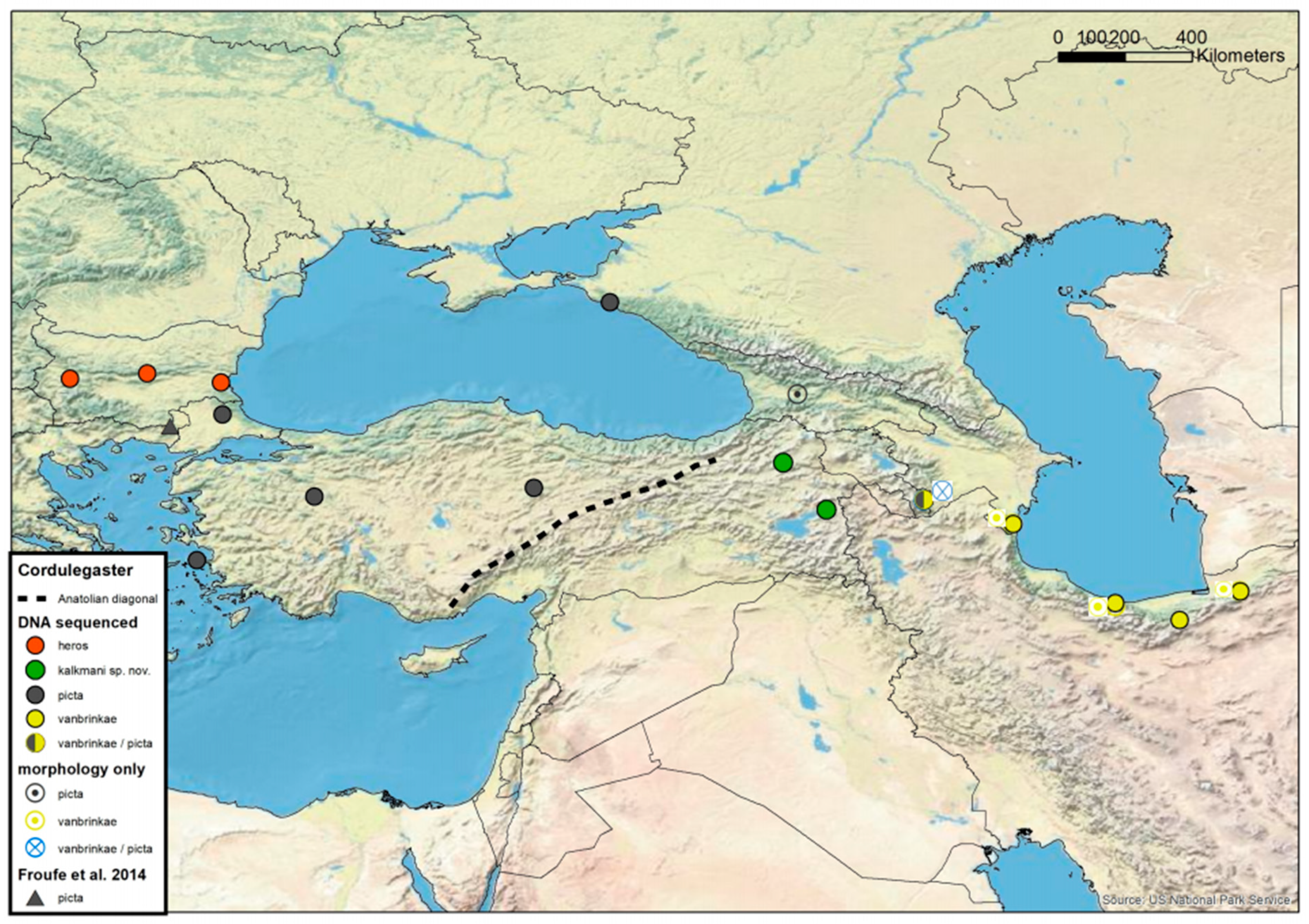
3.2. Cordulegaster boltonii-Group
3.3. Possible Hybridisation in the Cordulegaster boltonii-Group
3.4. Cordulegaster coronata-Group
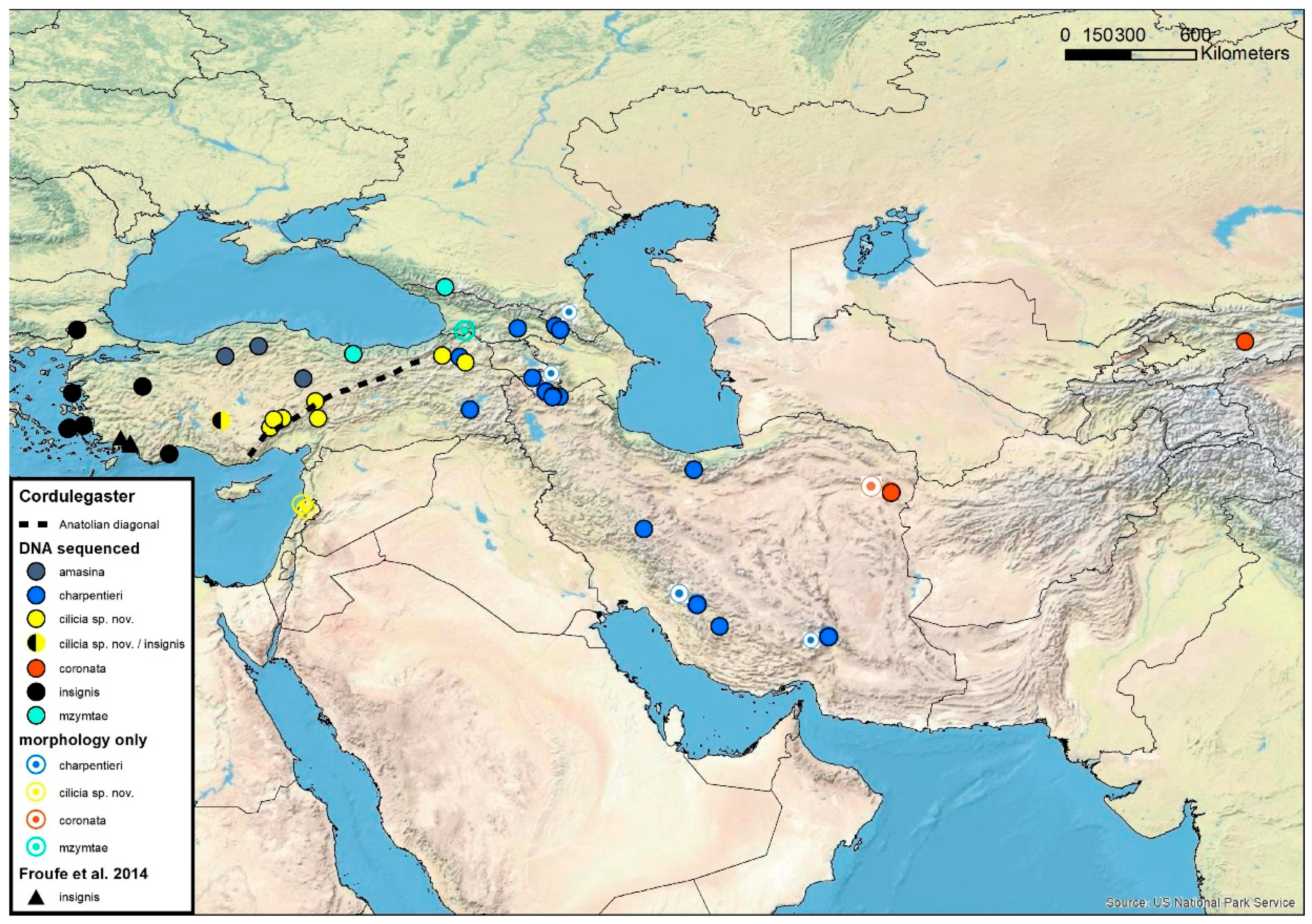
3.5. Cordulegaster bidentata-Group
3.6. Cordulegaster charpentieri-Complex
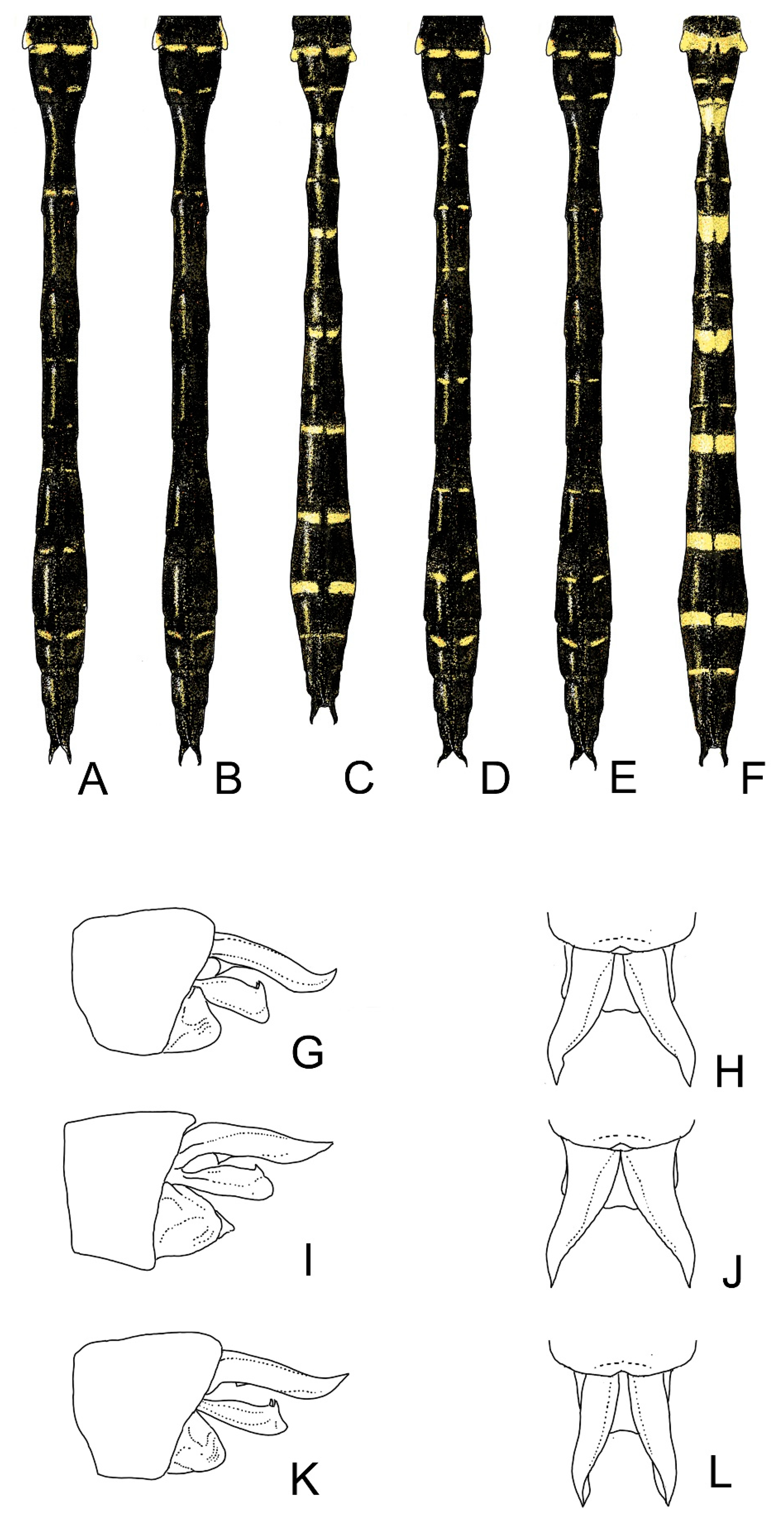
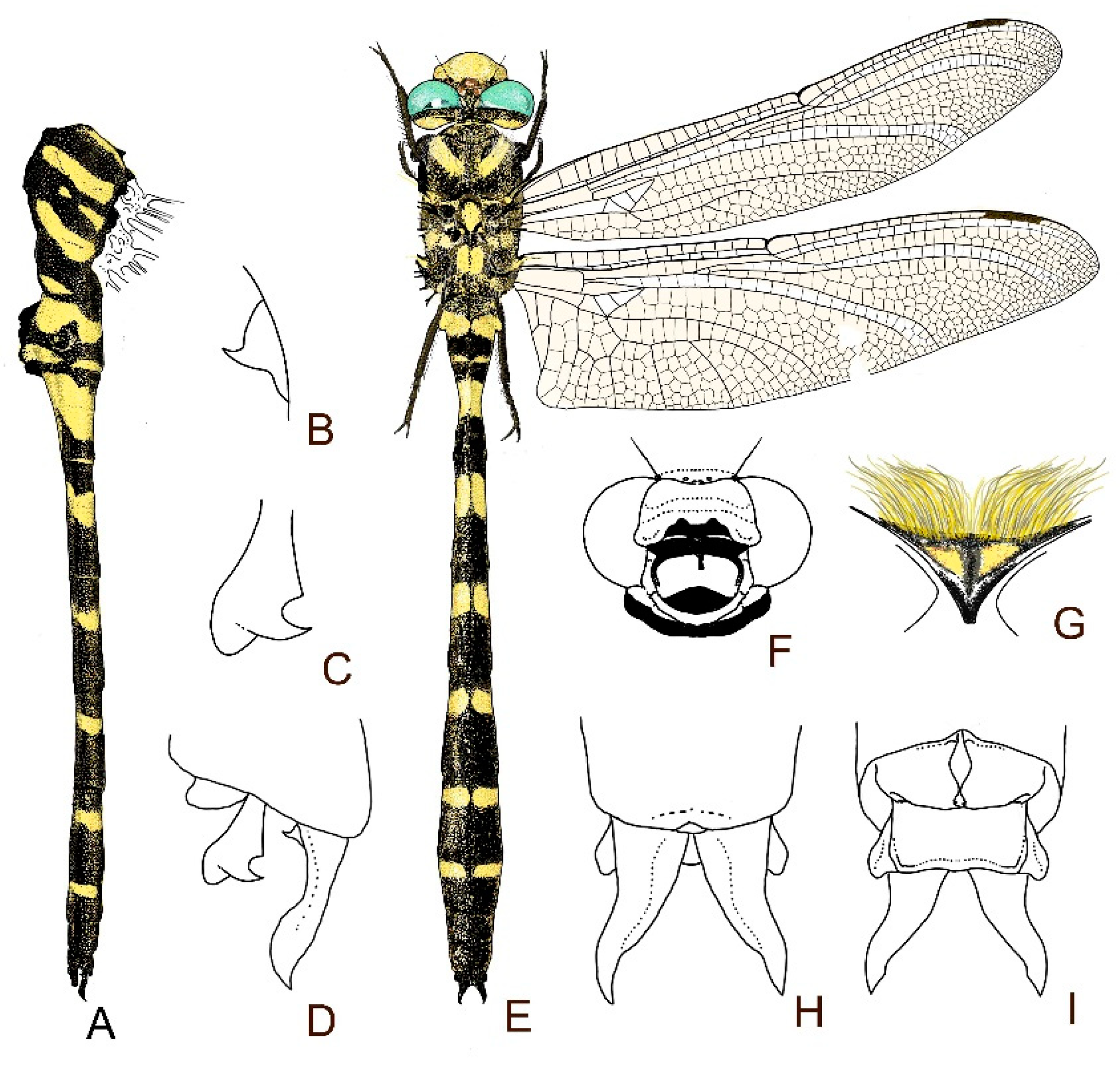
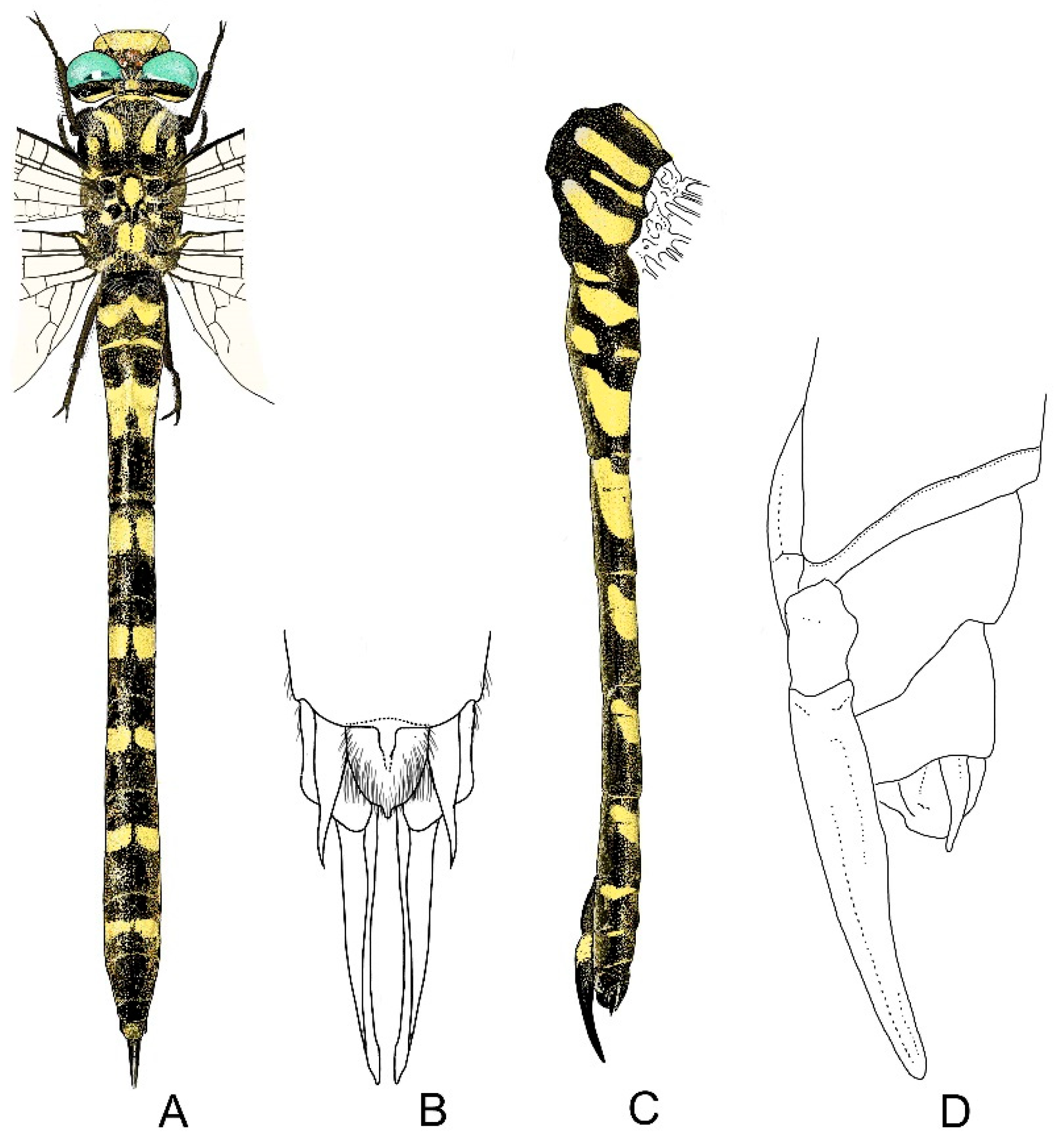
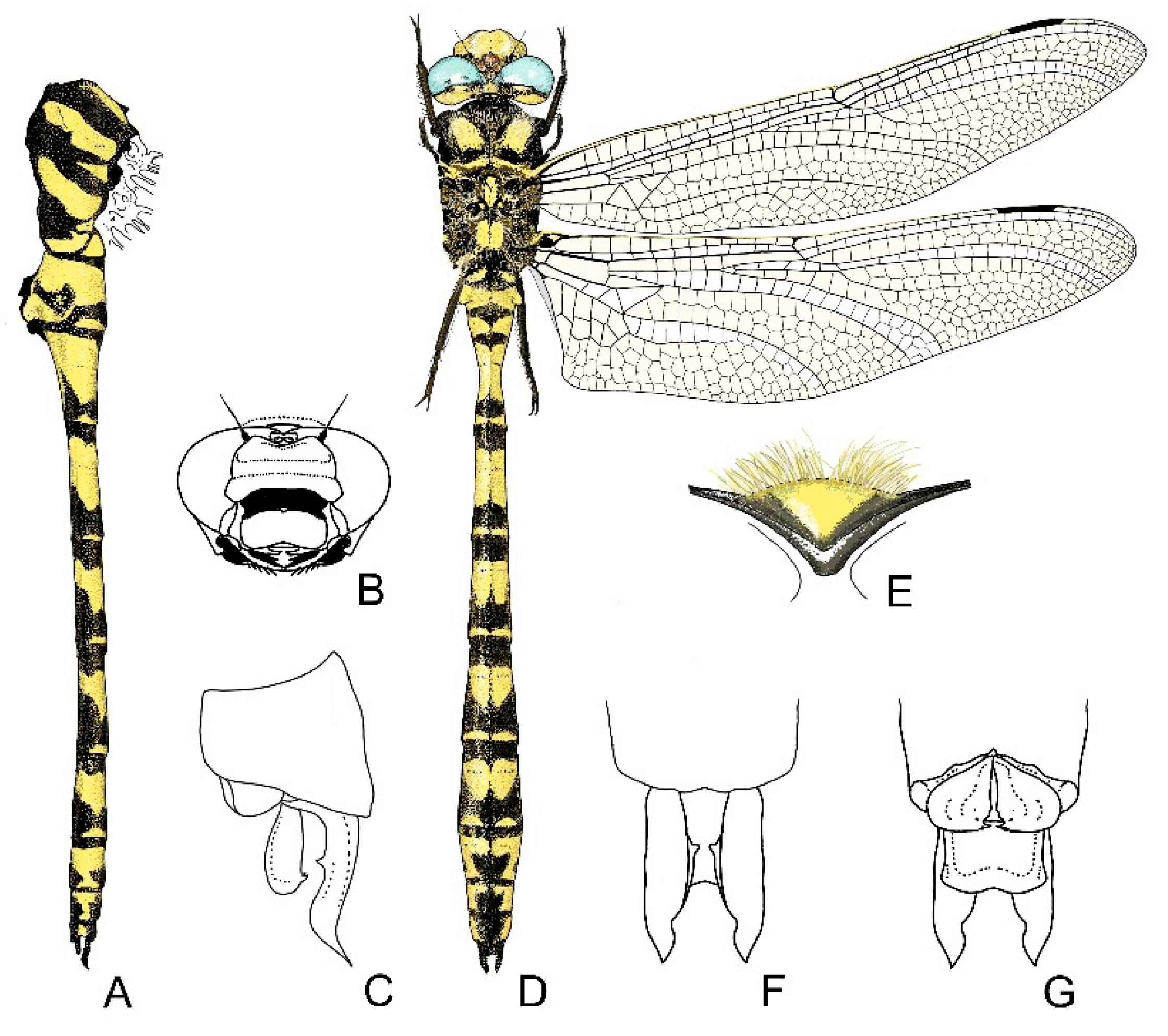
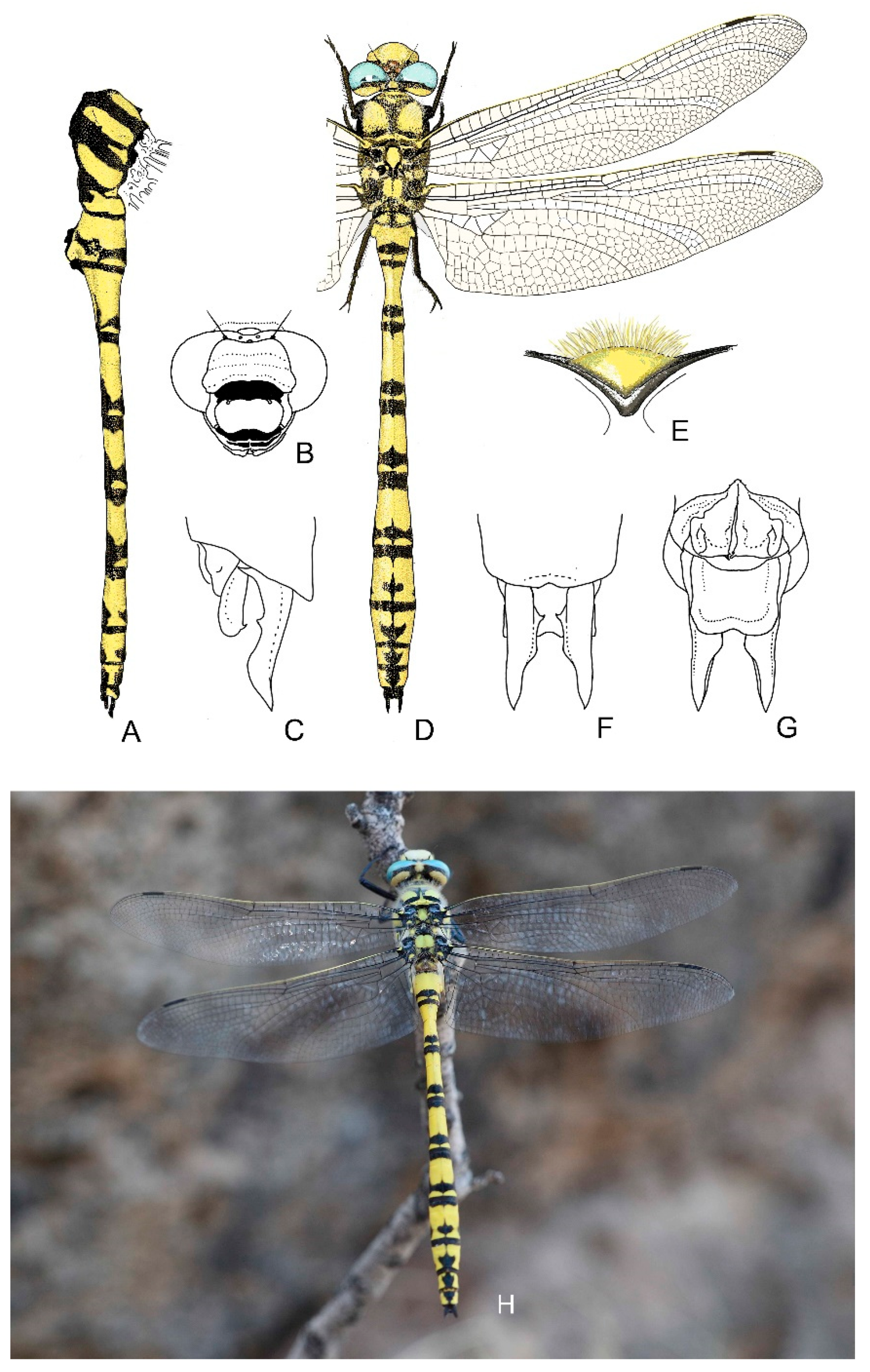
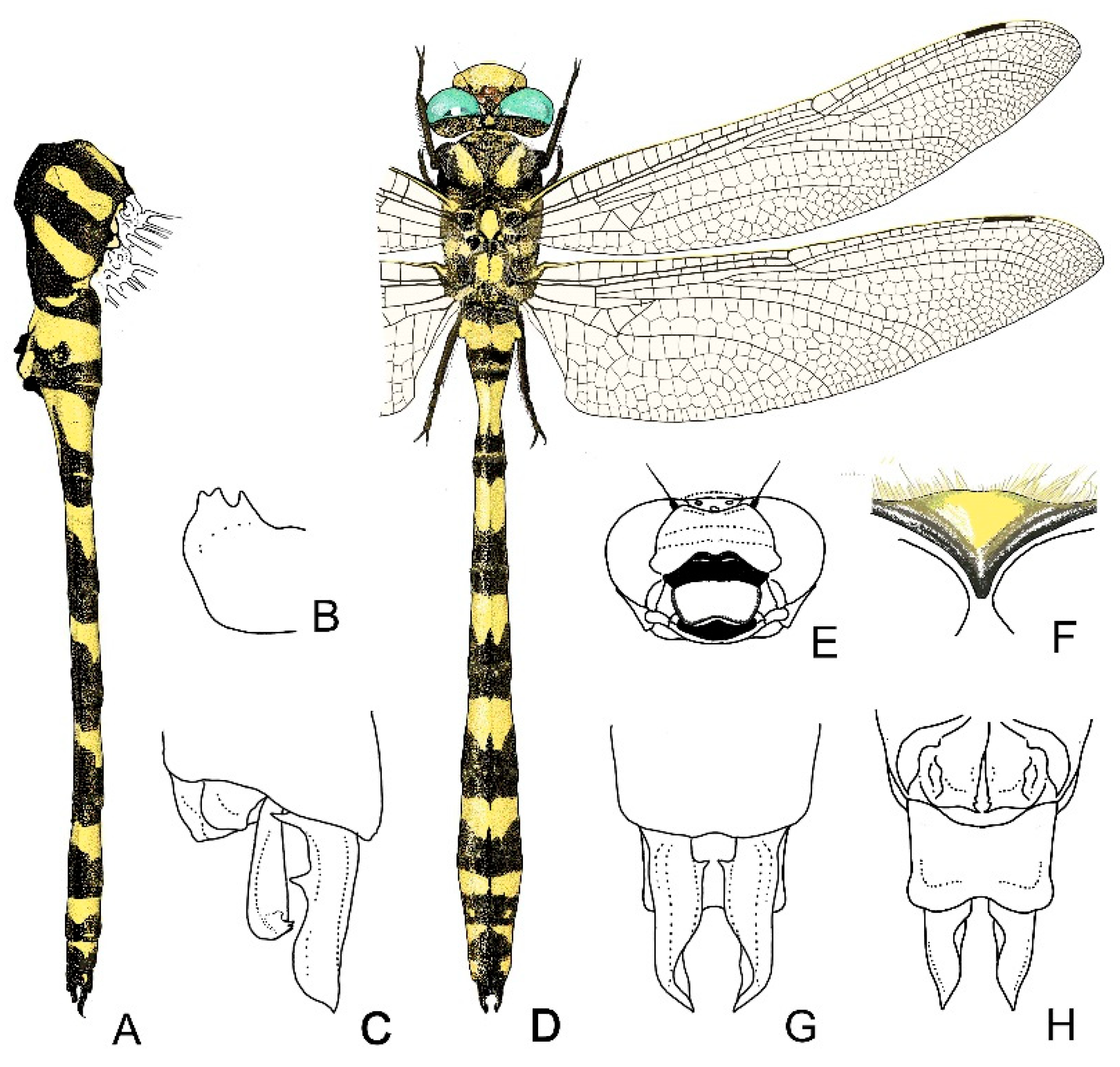
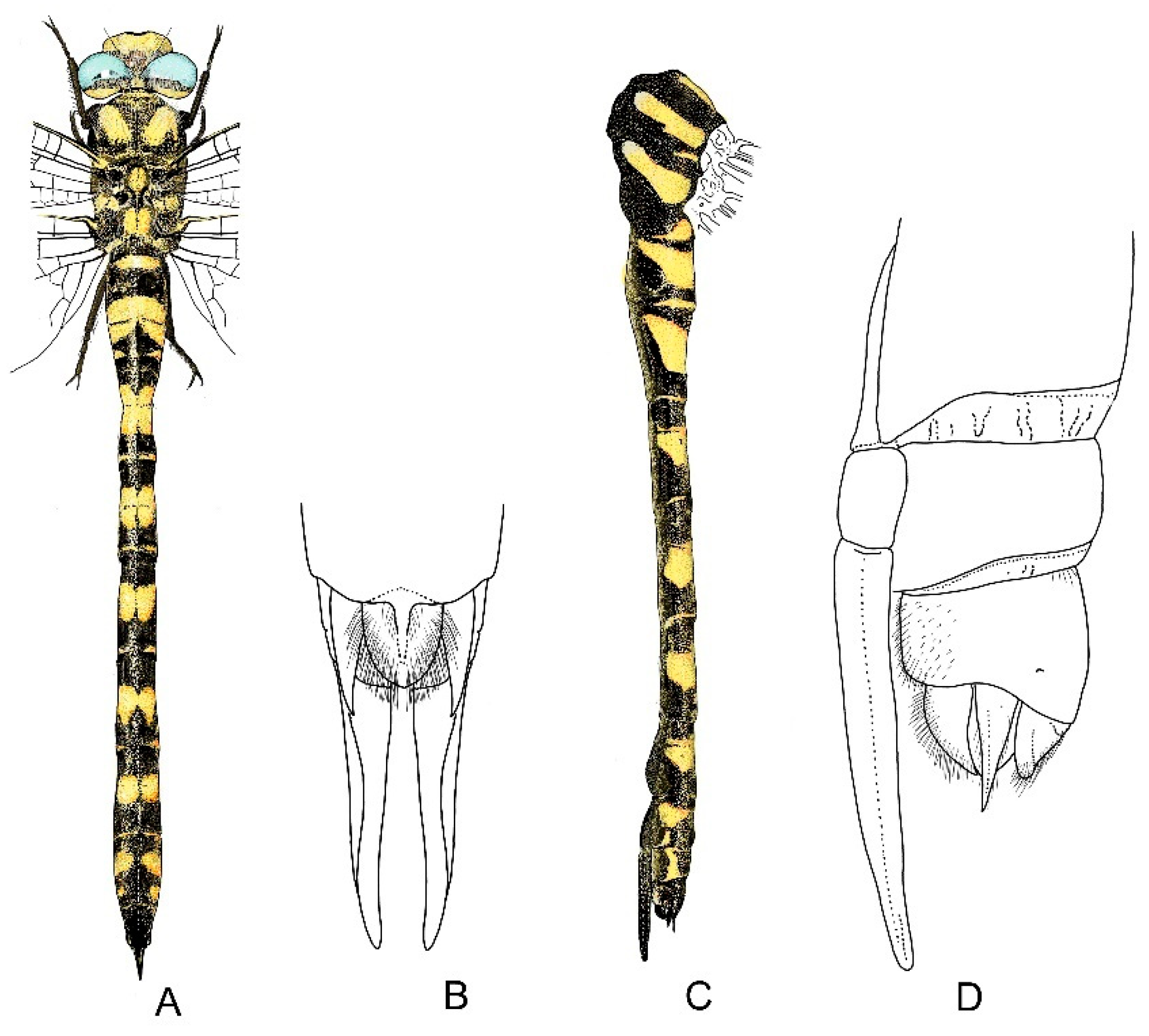
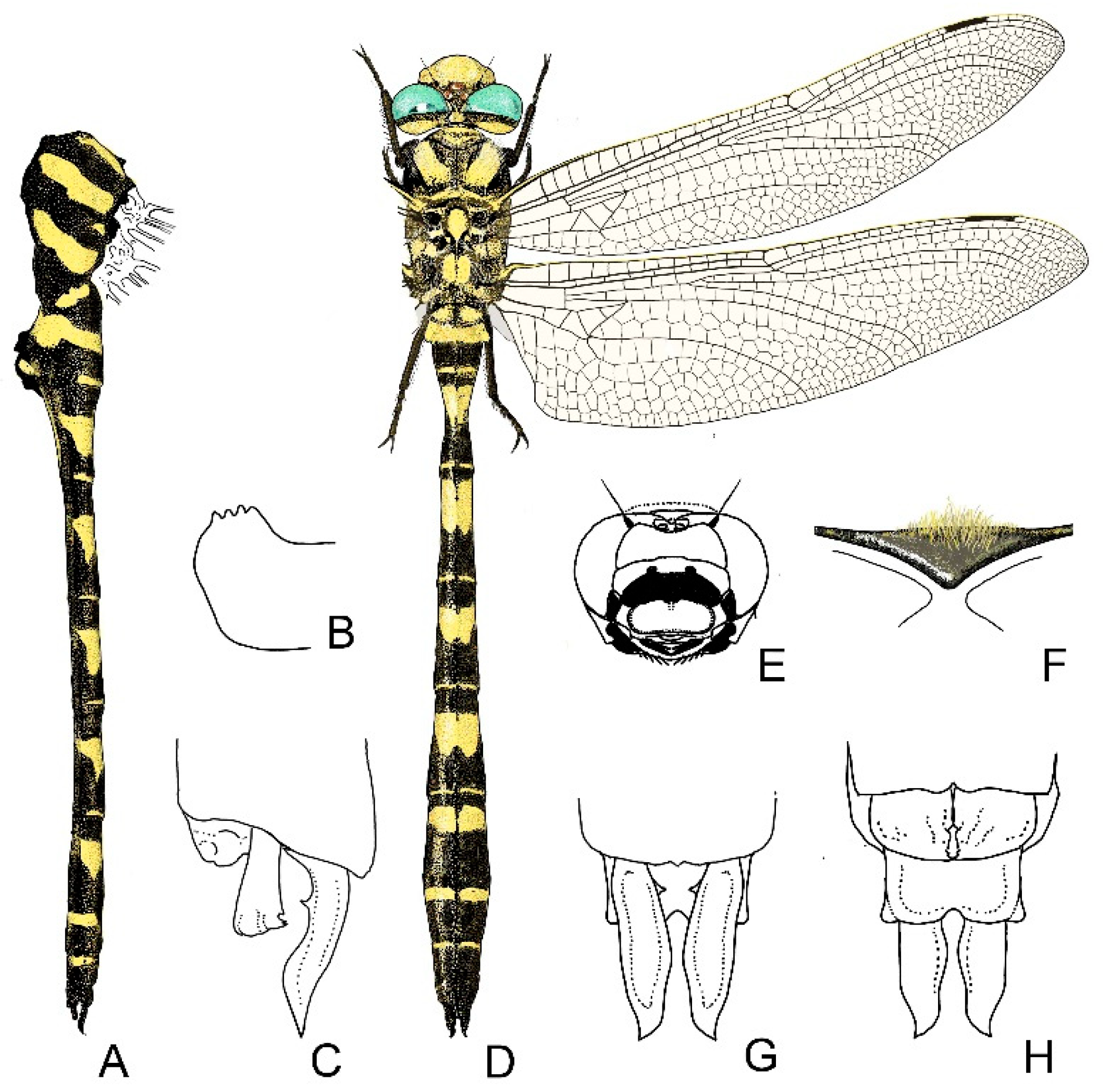
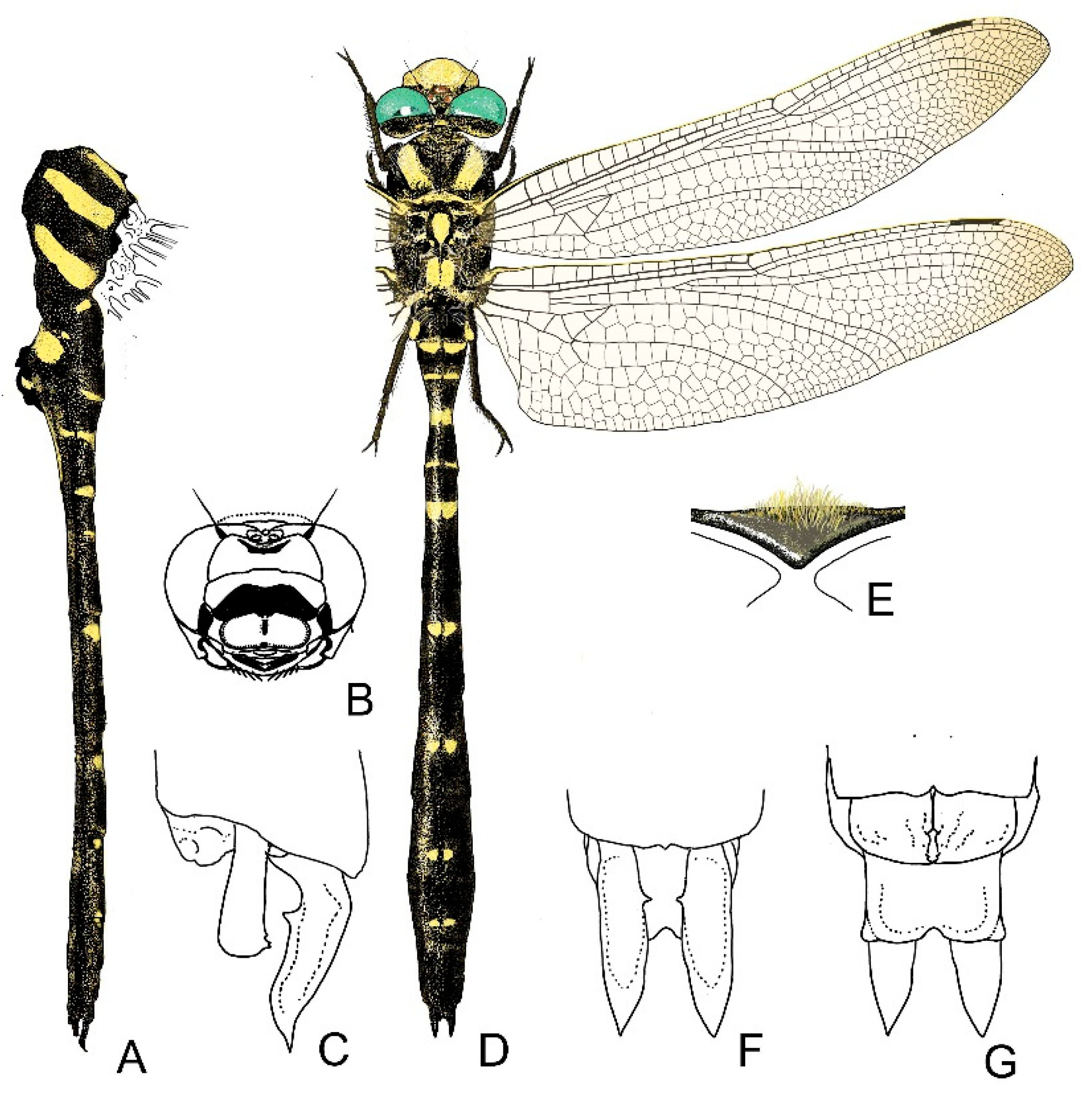
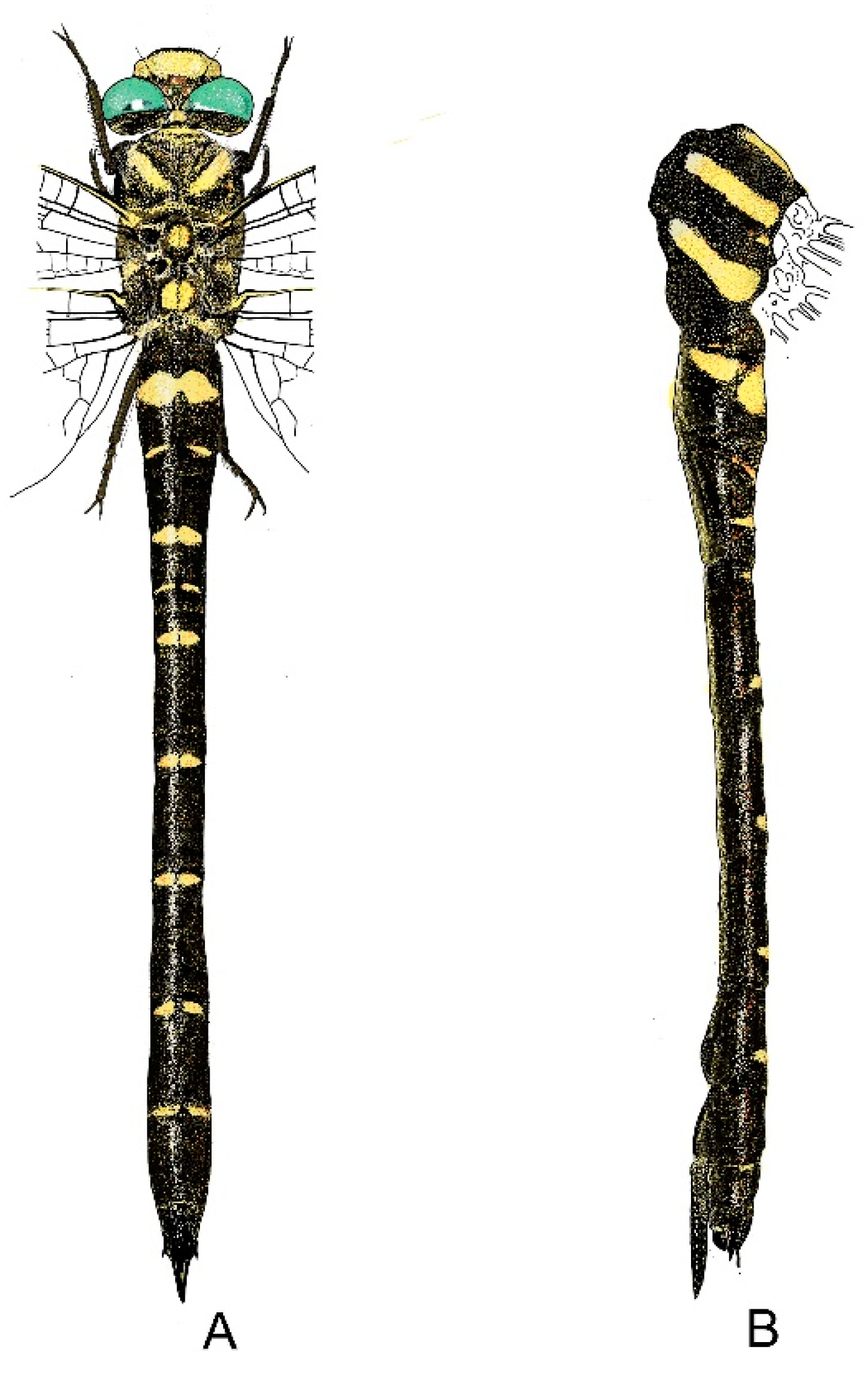
4. Descriptive Part
4.1. Cordulegaster boltonii-Group
4.2. Cordulegaster bidentata-Group
- Cordulegaster charpentieri (Kolenati, 1846) stat. rev.
- Cordulegaster insignis nobilis Morton, 1916, syn. nov.
- Cordulegaster insignis lagodechica Bartenev, 1930, syn. nov.
- Cordulegaster nachitschevanica Skvortsov and Snegovaya, 2015, syn. nov.
- Cordulegaster plagionyx Skvortsov and Snegovaya, 2015, syn. nov.
5. Discussion
5.1. General Discussion on the Genus Cordulegaster in the Western Palaearctic
5.2. Cordulegaster boltonii-Group
5.3. Cordulegaster bidentata-Group
5.4. Cordulegaster coronata-Group
5.5. Key for the Western–Palaearctic Cordulegaster
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Schorr, M.; Paulson, D. World Odonata List—Revision 23. Available online: https://www2.pugetsound.edu/academics/academic-resources/slater-museum/biodiversity-resources/dragonflies/world-odonata-list2/ (accessed on 15 October 2021).
- Fraser, F.C. A revision of the Fissilabioidea (Cordulegasteridae, Petaliidae and Petaluridae) (Order Odonata): Part 1—Cordulegasteridae. Mem. Indian Mus. 1929, 9, 69–167. [Google Scholar]
- Zhang, H. Dragonflies and Damselflies of China; Chongqing University Press: Chongqing, China, 2019; Volume 1, 736p. [Google Scholar]
- Garrison, R.W.; von Ellenrieder, N.; Louton, J.A. Dragonfly Genera of the New World; The Johns Hopkins University Press: Baltimore, MD, USA, 2006. [Google Scholar]
- Boudot, J.-P.; Kalkman, V. Odonata (dragonflies and damselflies). In The Status and Distribution of Freshwater Biodiversity in the Eastern Mediterranean; Smith, K.G., Barrios, V., Darwall, W.R.T., Numa, C., Eds.; IUCN: Gland, Switzerland, 2014; Chapter 5; pp. 57–69. [Google Scholar]
- Kalkman, V.J.; Boudot, J.-P.; Bernard, R.; Conze, K.-J.; De Knijf, D.; Dyatlova, E.; Ferreira, S.; Jović, M.; Ott, J.; Riservato, E.; et al. European Red List of Dragonflies; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Boudot, J.-P. Cordulegaster helladica. The IUCN Red List of Threatened Species 2020. e.T59706A140514673. Available online: https://www.iucnredlist.org/species/59706/140514673 (accessed on 15 October 2021).
- Boudot, J.-P. Cordulegaster helladica ssp. kastalia. The IUCN Red List of Threatened Species 2020. e.T63227A188019481. Available online: https://www.iucnredlist.org/species/63227/188019481 (accessed on 15 October 2021).
- Boudot, J.-P. Cordulegaster vanbrinkae. The IUCN Red List of Threatened Species 2006. E.T59709A11984778. Available online: https://www.iucnredlist.org/species/59709/11984778 (accessed on 15 October 2021).
- Smallshire, D.; Swash, A. Europe’s Dragonflies. A Field Guide to the Damselflies and Dragonflies; Princeton University Press & British Dragonfly Society: Woodstock, GA, USA, 2020. [Google Scholar]
- Van Pelt, G.J. Cordulegaster. In Field Guide to the Dragonflies of Britain and Europe; Dijkstra, K.-D., Lewington, R., Eds.; British Wildlife Publishing: Gillingham, UK, 2020; pp. 220–231. [Google Scholar]
- Verschuren, D. Revision of the larvae of Western–Palaearctic Cordulegaster Leach, 1815 (Odonata, Cordulegastridae) with a key to the considered taxa and a discussion on their affinity. Bull. Ann. Soc. R. Belg. Entomol. 1989, 125, 5–35. [Google Scholar]
- Froufe, E.; Roufe, E.; Ferreira, S.; Boudot, J.-P.; Alves, P.C.; Harris, D.J. Molecular phylogeny of the Western Palaearctic Cordulegaster taxa (Odonata Anisoptera: Cordulegastridae). Biol. J. Linn. Soc. 2014, 111, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Boudot, J.-P. Les Cordulegaster du Paléarctique occidental: Identification et répartition (Odonata, Anisoptera, Cordulegastridae). Martinia 2001, 17, 3–34. [Google Scholar]
- Corso, A. Morphological variability of Cordulegaster trinacriae in Italy (Odonata: Cordulegastridae). Odonatologica 2019, 48, 167–174. [Google Scholar]
- Skvortsov, V.E.; Snegovaya, N.Y. Two new species of Cordulegaster Leach, 1815 from Azerbaijan (Odonata, Cordulegastridae). J. Int. Dragonfly Fund 2015, 85, 1–22. [Google Scholar]
- Schneider, W.G. Verzeichniss der von Herrn Prof. Dr. Lowe im Sommer 1842 in der Türkei und Kleinasien gesammelten Neuroptera, nebst kurzer Beschreibung der neuen Arten. Stettin. Entomol. Ztg. 1845, 6, 110–116. [Google Scholar]
- Schmidt, E. Auf der Spur von Kellemisch. Entomol. Z. 1954, 64, 49–62, 65–72, 74–86, 92–93. [Google Scholar]
- Selys-Longchamps, E.; Hagen, H.A. Revue des Odonates ou Libellules d’Europe. In Mémoires de la Société Royale des Sciences de Liège; 1850; Volume 6, 408p, Available online: https://gallica.bnf.fr/ark:/12148/bpt6k26769q.texteImage (accessed on 15 October 2021).
- De Selys Longchamps, E. Monographie des Gomphines. In Mémoires de la Société Royale des Sciences de Liège; 1858; Volume 9, 460p, plates 1–23; Available online: https://www.biodiversitylibrary.org/bibliography/39398 (accessed on 15 October 2021).
- Kolenati, F.A. Insecta Caucasi. Meletemata Entomologica. Imperialis Academiae Scientiarum 1846, 5, 114–115. [Google Scholar]
- De Selys-Longchamps, E. Synopsis des Gomphines. Bull. Acad. R. Sci. Belg. 1854, 21, 23–112. [Google Scholar]
- De Selys-Longchamps, E. Odonates de l’Asie Mineure. Ann. Soc. Entomol. Belg. 1887, 31, 1–85. [Google Scholar]
- Waterston, A.R. On the Genus Cordulegaster Leach, 1815 (Odonata) with special reference to the Sicilian species. Trans. R. Soc. Edinb. 1976, 69, 457–466. [Google Scholar] [CrossRef]
- Dumont, H.J. Aeschna charpentieri Kolenati, 1846, a synonym of Cordulegaster insignis Schneider, 1845 and on the correct status of Cordulegaster charpentieri auctorum (Anisoptera: Cordulegastridae). Odonatologica 1976, 5, 313–321. [Google Scholar]
- Morton, K.J. Some Palaearctic species of Cordulegaster. Trans. Entomol. Soc. Lond. 1916, 63, 273–290. [Google Scholar] [CrossRef]
- Bartenev, A.N. Neue Arten und Varietäten der Odonaten des West-Kaukasus. Zool. Anz. 1929, 85, 54–68. [Google Scholar]
- Bartenev, A.N. Die paläarktischen Arten der Untergattung Cordulegaster Leach. In Travaux de la Station Biologique du Caucase du Nord; Gorsky Institut Agronomique: Vladicaucase, Russia, 1930; Volume 3, pp. 1–32. (In Russian) [Google Scholar]
- Theischinger, G. Cordulegaster heros sp. nov. und Cordulegaster heros pelionensis ssp. nov., zwei neue Taxa des Cordulegaster boltonii (Donovan)-Komplexes aus Europa (Ansioptera: Cordulegastridae). Odonatologica 1979, 8, 23–38. [Google Scholar]
- Lohmann, H. Revision der Cordulegastridae. 2. Beschreibung neuer Arten in den Gattungen Cordulegaster, Anotogaster, Neallogaster und Sonjagaster (Anisoptera). Odonatologica 1993, 22, 273–294. [Google Scholar]
- St. Quentin, D. Zum Vorkommen von Cordulegaster insignis Schneider in Rumänien. Studii Communicari Sibiu 1971, 16, 205–208. [Google Scholar]
- Schmidt, E. Ergebnisse der Deutschen Afghanistan–Expedition 1956 der Landessammlungen für Naturkunde Karlsruhe sowie der Expeditionen J. Klapperich, Bonn 1952–53 und Dr. K. Lindberg, Lund (Schweden) 1957–1960. Beiträge Nat. Forsch. SW Dtschl. 1961, 19, 399–435. [Google Scholar]
- Asahina, S. A Revision of the Himalayan Dragonflies of the Genus Neallogaster (Odonata, Cordulegastridae); Bulletin of the National Science Museum: Tokyo, Japan, 1982; Volume 8, pp. 153–171. [Google Scholar]
- Garcia-Castellanos, D.; Estrada, F.; Jiménez-Munt, I.; Gorini, C.; Fernàndez, M.; Vergés, J.; De Vicente, R. Catastrophic flood of the Mediterranean after the Messinian salinity crisis. Nature 2009, 462, 778–781. [Google Scholar] [CrossRef] [PubMed]
- Solano, E.; Hardersen, S.; Audisio, P.; Amorosi, V.; Senczuk, G.; Antonini, G. Asymmetric hybridization in Cordulegaster (Odonata: Cordulegastridae): Secondary postglacial contact and the possible role of mechanical constraints. Ecol. Evol. 2018, 8, 9657–9671. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitt, G.M. Post–glacial re–colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Hewitt, G.M. Speciation, hybrid zones and phylogeography—Or seeing genes in space and time. Mol. Ecol. 2001, 10, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Gür, H. The Anatolian diagonal revisited: Testing the ecological basis of a biogeographic boundary. Zool. Middle East. 2016, 62, 189–199. [Google Scholar] [CrossRef]
- Davis, P.H. Distribution patterns in Anatolia with particular reference to endemism. In Plant Life of South–West Asia; Davis, P.H., Harper, P.C., Hedge, I.C., Eds.; Botanical Society of Edinburgh: Edinburgh, UK, 1971; pp. 15–27. [Google Scholar]
- Galimberti, A.; Assandri, G.; Maggioni, D.; Ramazzotti, F.; Baroni, D.; Bazzi, G.; Chiandetti, I.; Corso, A.; Ferri, V.; Galuppi, M.; et al. DNA barcoding and eDNA-based biomonitoring of Italian Odonata. Mol. Ecol. Resour. 2020, 21, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Skvortsov, V.E. The Dragonflies of Eastern Europe and Caucasus: An Illustrated Guide; KMK Scientific Press Ltd.: Moscow, Russia, 2010. [Google Scholar]
- Boudot, J.-P.; Borisov, S.N.; De Knijf, G.; van Grunsven, R.; Schröter, A.; Kalkman, V.J. Atlas of the dragonflies and damselflies of West and Central Asia. In Brachytron 22; Nederlandse Vereniging voor Libellenstudie: Den Haag, The Netherlands, 2021; pp. 4–238. [Google Scholar]
- Schneider, T.; Ikemeyer, D.; Müller, O.; Dumont, H.J. Checklist of the dragonflies (Odonata) of Iran with new records and notes on distribution and taxonomy. Zootaxa 2018, 4394, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Ikemeyer, D. The Damselflies and Dragonflies of Iran—Odonata Persica; NIBUK, Natur in Buch und Kunst: Ruppichteroth, Germany, 2019; 247p. [Google Scholar]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koroiva, R.; Pepinelli, M.; Rodrigues, M.E.; de Oliviera, R.F.; Lorenz-Lemke, A.P.; Kvist, S. DNA barcoding of odonates from the Upper Plata basin: Database creation and genetic diversity estimation. PLoS ONE 2017, 12, e0182283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimenta, A.L.A.; Pinto, A.P.; Takiya, D.M. Integrative taxonomy and phylogeny of the damselfly genus Forcepsioneura Lencioni, 1999 (Odonata: Coenagrionidae: Protoneurinae) with description of two new species from the Brazilian Atlantic Forest. Arthropod Syst. Phylogenie 2019, 77, 397–415. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesische Phylogenetik mit BEAUti und dem BEAST 1.7. Mol. Biol. Evol. 2012, 22, 1185–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambout, A. GitHub. Available online: www.https://github.com/rambaut/figtree (accessed on 15 October 2021).
- Flouri, T.; Jiao, X.; Rannala, B.; Yang, Z. Species Tree Inference with BPP using Genomic Sequences and the Multispecies Coalescent. Mol. Biol. Evol. 2018, 35, 2585–2593. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Van Pelt, G.J.; Vierstraete, A.; Dumont, H.J.; Schneider, T. Case @@@@—Aeschna charpentieri Kolenati, 1846 (Insecta, Odonata): Proposed Replacement of the Current Neotype by a New Neotype; Bulletin of Zoological Nomenclature; 2021; unpublished. [Google Scholar]
- St. Quentin, D. Der Rassenkreis Cordulegaster boltonii (Donovan) (Odonata). Entomol. Nachr. Österreichischer Und Schweiz. Entomol. 1952, 4, 73–75. [Google Scholar]
- Boudot, J.-P.; Jacquemin, G. Revision of Cordulegaster boltonii (Donovan, 1807) in southwestern Europe and northern Africa, with description of C. b. iberica ssp. from Spain (Anisoptera: Cordulegastridae). Odonatologica 1995, 24, 149–173. [Google Scholar]
- Bernard, R.; Daraz, B. Cordulegaster heros and Somatochlora meridionalis in Ukraine: Solving the zoogeographical puzzle at their northern range limits (Odonata: Cordulegastridae, Corduliidae). Odonatologica 2015, 44, 255–278. [Google Scholar]
- Kosterin, O.; Solovyev, V. Odonata found in mid-summer 2015 and 2016 at the north–westernmost Black Sea Coast of the Caucasus, with the first record of Cordulegaster picta Selys, 1854 in Russian Federation. J. Int. Dragonfly Fund 2017, 107, 1–43. [Google Scholar]
- Schröter, A.; Seehausen, M.; Kunz, B.; Günther, A.; Schneider, T.; Jödicke, R. Update of the Odonata fauna of Georgia, southern Caucasus ecoregion. Odonatologica 2015, 44, 279–342. [Google Scholar]
- Schneider, T.; Schneider, E.; Schneider, J.; Müller, O. Rediscovery of Cordulegaster vanbrinkae in Iran (Odonata: Cordulegastridae). Odonatologica 2014, 43, 25–34. [Google Scholar]
- Holuša, O.; Dalecký, V.; Imanpour Namin, J. Habitat choice of Cordulegaster vanbrinkae in Iran (Odonata: Cordulegastridae). Odonatologica 2015, 44, 11–20. [Google Scholar]
- Rastegar, J.; Havaskary, M.; Khodaparast, S.; Rafeii, A. A contribution to the knowledge of Odonata (Insecta) from West Azarbaijan province, northwestern Iran. Entomofauna 2013, 34, 369–375. [Google Scholar]
- Ananian, V.; Tailly, M. Cordulegaster vanbrinkae Lohmann, 1993 (Odonata: Anisoptera) discovered in Armenia. J. Int. Dragonfly Fund 2012, 46, 1–11. [Google Scholar]
- Schweighofer, W. Syntopes Vorkommen von Cordulegaster boltonii und C. heros an einem Bach im westlichen Niederösterreich (Odonata: Cordulegastridae). Libellula 2008, 27, 1–32. [Google Scholar]
- Van Riemsdijk, I.; Arntzen, J.W.; Bogaerts, S.; Franzen, M.; Litvinchuk, S.N.; Olgun, K.; Wielstra, B. The Near East as a cradle of biodiversity: A phylogeography of banded newts (genus Ommatotriton) reveals extensive inter- and intraspecific genetic differentiation. Mol. Phylogenet. Evol. 2017, 114, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Asahina, S. The Odonata of Iraq. Jpn. J. Zool. 1973, 17, 17–36. [Google Scholar]
- Kosswig, C. Zoogeography of the Near East. Syst. Zool. 1955, 4, 49–73. [Google Scholar] [CrossRef]
- Çıplak, B.; Demirsoy, A.; Bozcuk, A.N. Distribution of Orthoptera in relation to the Anatolian diagonal in Turkey. Articulata 1993, 8, 1–20. [Google Scholar]
- Bilgin, R. Back to the Suture: The distribution of intraspecific genetic diversity in and around Anatolia. Int. J. Mol. Sci. 2011, 12, 4080–4103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapli, P.; Botoni, D.; Ilgaz, C.; Kumlutas, Y.; Avcı, A.; Rastegar-Pouyani, N.; Fathinia, B.; Lymberakis, P.; Ahmadzadeh, F.; Poulakakis, N. Molecular phylogeny and historical biogeography of the Anatolian lizard Apathya (Squamata, Lacertidae). Mol. Phylogenet. Evol. 2013, 66, 992–1001. [Google Scholar] [CrossRef]
- Erinç, S. Changes in the physical environment in Turkey since the end of the last glacial. In The Environmental History of the Near and Middle East since the Last Ice Age; Brice, W.C., Ed.; Academic Press: London, UK, 1978; pp. 87–110. [Google Scholar]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Species Identified | Former Taxon/Notes | Latitude | Longitude | Country | Collector/Reference | Genbank COI | Genbank ITS |
|---|---|---|---|---|---|---|---|---|
| bidentata-group | amasina | 39.5629 | 34.1894 | Turkey | leg. G.J. van Pelt | MK779843 | MK861474 | |
| bidentata-group | amasina | 40.9765 | 34.1894 | Turkey | leg. G.J. van Pelt | MK779844 | MK861470 | |
| bidentata-group | amasina | male redescription | 40.5915 | 32.6597 | Turkey | leg. G.J. van Pelt | MK779845 | MK861469 |
| bidentata-group | amasina | 39.5629 | 34.1894 | Turkey | leg. G.J. van Pelt | no data | MK861473 | |
| bidentata-group | bidentata | 50.8101 | 14.7763 | Czech Republic | leg. M. Waldhauser | MK779814 | no data | |
| bidentata-group | bidentata | morphology only | France | leg. T. Schneider | ||||
| bidentata-group | bidentata | morphology only | France | leg. T. Schneider | ||||
| bidentata-group | bidentata | morphology only | France | leg. T. Schneider | ||||
| bidentata-group | bidentata | morphology only | France | leg. T. Schneider | ||||
| bidentata-group | bidentata | morphology only | France | leg. T. Schneider | ||||
| bidentata-group | bidentata | morphology only | France | leg. T. Schneider | ||||
| bidentata-group | bidentata | morphology only | France | leg. T. Schneider | ||||
| bidentata-group | bidentata | morphology only | Germany | leg. T. Schneider | ||||
| bidentata-group | bidentata | morpholgy only | 39.4386 | 23.0464 | Greece | leg. T. Schneider | ||
| bidentata-group | bidentata | was C. b. sicilica, morphology only | Italy | leg. T. Schneider | ||||
| bidentata-group | bidentata | was C. b. sicilica, morphology only | Italy | leg. T. Schneider | ||||
| bidentata-group | bidentata | was C. b. sicilica, morphology only | Italy | leg. T. Schneider | ||||
| bidentata-group | charpentieri | 38.9487 | 46.1958 | Armenia | leg. V. Ananian | MK779820 | MK861491 | |
| bidentata-group | charpentieri | 38.9131 | 46.4604 | Armenia | leg. V. Ananian | MK779821 | MK861463 | |
| bidentata-group | charpentieri | 38.8956 | 46.1875 | Armenia | leg. V. Ananian | no data | MK86149 | |
| bidentata-group | charpentieri | 38.8956 | 46.1875 | Armenia | leg. V. Ananian | no data | MK861467 | |
| bidentata-group | charpentieri | 38.9487 | 46.1958 | Armenia | leg. V. Ananian | no data | MK861460 | |
| bidentata-group | charpentieri | 39.7038 | 45.3468 | Armenia | leg. V. Ananian | no data | MK861462 | |
| bidentata-group | charpentieri | was nachitschevanica | 39.1103 | 45.9144 | Azerbaijan | leg. N. Snegovaya | MK779823 | MK861463 |
| bidentata-group | charpentieri | was plagionyx | 41.6761 | 46.4931 | Azerbaijan | leg. N. Snegovaya | MK779828 | MK861466 |
| bidentata-group | charpentieri | male, was nachitschevanica, morphology only | 39.1103 | 45.9144 | Azerbaijan | leg. N. Snegovaya | ||
| bidentata-group | charpentieri | female, was plagionyx, morphology only | 41.6761 | 46.4931 | Azerbaijan | leg. N. Snegovaya | ||
| bidentata-group | charpentieri | male, was plagionyx, morphology only | 41.6761 | 46.4931 | Azerbaijan | leg. N. Snegovaya | ||
| bidentata-group | charpentieri | charpentieri lagodechica, larva | 41.8404 | 46.2846 | Georgia | leg. M. Lemke | MK779824 | MK861465 |
| bidentata-group | charpentieri | 41.7458 | 44.7426 | Georgia | leg. N. Snegovaya | MK779825 | no data | |
| bidentata-group | charpentieri | charpentieri lagodechica, larva | 41.8404 | 46.2846 | Georgia | leg. M. Lemke | MK779826 | MK861477 |
| bidentata-group | charpentieri | was nobilis | 29.4311 | 53.0842 | Iran | leg. T. Schneider | MN623223 | MN612638 |
| bidentata-group | charpentieri | was nobilis, male | 33.4597 | 49.9472 | Iran | leg. T. Schneider | MK779819 | MK861457 |
| bidentata-group | charpentieri | was nobilis | 35.8926 | 52.0125 | Iran | leg. M. Waldhauser | MK779829 | MK861459 |
| bidentata-group | charpentieri | was nobilis, female paratype | 30.3339 | 52.1564 | Iran | leg. T. Schneider | MK779830 | MK861496 |
| bidentata-group | charpentieri | was nobilis | 29.0022 | 57.5869 | Iran | leg. T. Schneider | MK779831 | MK861471 |
| bidentata-group | charpentieri | was nobilis; morphology only | 29.0022 | 57.5869 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis; morphology only | 29.0022 | 57.5869 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis; morphology only | 29.0022 | 57.5869 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, male paratype, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, female, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, female, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, female, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, male, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, male, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, male, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, paratype male, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | was nobilis, paratype male, morphology only | 30.3339 | 52.1564 | Iran | leg. T. Schneider | ||
| bidentata-group | charpentieri | 40.5616 | 42.3471 | Turkey | leg. G.J. van Pelt | MK779822 | MK861458 | |
| bidentata-group | charpentieri | male redescription, neotype | 38.3834 | 42.7723 | Turkey | leg. G.J. van Pelt | MK779827 | MK861476 |
| bidentata-group | cilicia sp. nov. | was insignis, paratype, morphology only | 34.2511 | 36.0111 | Lebanon | leg. G.A. Mavromoustakis | ||
| bidentata-group | cilicia sp. nov. | was insignis, paratype, morphology only | 34.2511 | 36.0111 | Lebanon | leg. G.A. Mavromoustakis | ||
| bidentata-group | cilicia sp. nov. | was insignis, morphology only | 34.2511 | 36.0111 | Lebanon | leg. G.A. Mavromoustakis | ||
| bidentata-group | cilicia sp. nov. | was insignis, female paratype, morphology only | 34.4578 | 35.8414 | Lebanon | leg. G.A. Mavromoustakis | ||
| bidentata-group | cilicia sp. nov. | was insignis, morphology only | 34.4578 | 35.8414 | Lebanon | leg. G.A. Mavromoustakis | ||
| bidentata-group | cilicia sp. nov. | was insignis, morphology only | 34.4578 | 35.8414 | Lebanon | leg. G.A. Mavromoustakis | ||
| bidentata-group | cilicia sp. nov. | was charpentieri, paratype | 40.6114 | 41.6286 | Turkey | leg. G.J. van Pelt | MK779832 | MK861494 |
| bidentata-group | cilicia sp. nov. | was insignis, paratype, morphology only | 38.7205 | 36.3950 | Turkey | leg. G.J. van Pelt | MK779833 | MK861475 |
| bidentata-group | cilicia sp. nov. | was insignis, paratype, morphology only | 38.0164 | 35.0311 | Turkey | leg. G.J. van Pelt | MK779834 | no data |
| bidentata-group | cilicia sp. nov. | was insignis, male holotype | 38.0614 | 36.4722 | Turkey | leg. G.J. van Pelt | MK779835 | MK861498 |
| bidentata-group | cilicia sp. nov. | was insignis, male paratype | 37.9698 | 34.6767 | Turkey | leg. G.J. van Pelt | MK779836 | MK861468 |
| bidentata-group | cilicia sp. nov. | was insignis | 37.6512 | 34.5328 | Turkey | leg. G.J. van Pelt | MK779837 | MK861461 |
| bidentata-group | cilicia sp. nov. | was insignis | 37.9698 | 34.6766 | Turkey | leg. G.J. van Pelt | MK779839 | MK861472 |
| bidentata-group | cilicia sp. nov. | was insignis | 40.3334 | 42.5905 | Turkey | leg. G.J. van Pelt | MK779840 | MK861495 |
| bidentata-group | cilicia sp. nov./insignis | hybrid: cilcia × insignis | 37.9293 | 32.5000 | Turkey | leg. G.J. van Pelt | MK779838 | MK861497 |
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 34.9569 | 60.1672 | Iran | leg. T. Schneider | MK779857 | MK861452 |
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 34.9569 | 60.1672 | Iran | leg. T. Schneider | ||
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 34.9569 | 60.1672 | Iran | leg. T. Schneider | ||
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 34.9569 | 60.1672 | Iran | leg. T. Schneider | ||
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 34.9569 | 60.1672 | Iran | leg. T. Schneider | ||
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 34.9569 | 60.1672 | Iran | leg. T. Schneider | ||
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 34.9569 | 60.1672 | Iran | leg. T. Schneider | ||
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 34.9569 | 60.1672 | Iran | leg. T. Schneider | ||
| coronata-group | coronata | was sometimes treated as ssp. of insignis | 41.2044 | 74.7661 | Kyrgyzstan | leg. H. Dumont | MK779858 | MK861453 |
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.4887 | 22.5048 | Greece | leg. T. Schneider | ||
| bidentata-group | helladica | morphology only | 38.4887 | 22.5048 | Greece | leg. T. Schneider | ||
| boltonii-group | heros | morphology only | 48.2132 | 16.3847 | Austria | leg. T. Schneider | ||
| boltonii-group | heros | morphology only | 48.2132 | 16.3847 | Austria | leg. T. Schneider | ||
| boltonii-group | heros | 42.4929 | 27.4733 | Bulgaria | leg. G.J. van Pelt | MK779810 | no data | |
| boltonii-group | heros | 42.7339 | 25.4858 | Bulgaria | leg. G.J. van Pelt | MK779810 | no data | |
| boltonii-group | heros | 42.5937 | 23.4102 | Bulgaria | leg. G.J. van Pelt | MK779812 | MK861451 | |
| boltonii-group | heros | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| boltonii-group | heros | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| boltonii-group | heros | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| boltonii-group | heros | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| boltonii-group | heros | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| boltonii-group | heros | morphology only | 38.0353 | 22.1098 | Greece | leg. T. Schneider | ||
| boltonii-group | heros | 42.1837 | 19.2915 | Montenegro | leg. G. De Knijf | MK779813 | no data | |
| bidentata-group | insignis | 37.7284 | 26.8196 | Greece | leg. T. Schneider | MW353685 | MW363787 | |
| bidentata-group | insignis | 37.5833 | 26.1667 | Greece | leg. T. Schneider | MW353686 | MW363788 | |
| bidentata-group | insignis | 37.5833 | 26.1667 | Greece | leg. T. Schneider | MW353687 | MW363789 | |
| bidentata-group | insignis | 39.0690 | 26.3468 | Greece | leg. W. Lopau | MW353702 | no data | |
| bidentata-group | insignis | 36.9287 | 28.7511 | Turkey | Froufe et al., 2014 | KF584941 | KF584979 | |
| bidentata-group | insignis | 37.0287 | 28.7554 | Turkey | Froufe et al., 2014 | KF584941 | no data | |
| bidentata-group | insignis | 37.0287 | 28.7554 | Turkey | Froufe et al., 2014 | KF584941 | no data | |
| bidentata-group | insignis | 37.0287 | 28.7554 | Turkey | Froufe et al., 2014 | KF584941 | KF584979 | |
| bidentata-group | insignis | 37.2196 | 28.3658 | Turkey | Froufe et al., 2014 | KF584942 | no data | |
| bidentata-group | insignis | 37.0287 | 28.7554 | Turkey | Froufe et al., 2014 | KF584943 | KF584979 | |
| bidentata-group | insignis | 41.6771 | 26.5557 | Turkey | leg. G.J. van Pelt | MK779846 | MK861486 | |
| bidentata-group | insignis | 39.3444 | 29.2579 | Turkey | leg. G.J. van Pelt | MK779848 | MK861487 | |
| bidentata-group | insignis | 36.5595 | 30.3501 | Turkey | leg. T. Schneider | MW353707 | MW363797 | |
| boltonii-group | kalkmani sp. nov. | was picta, male holotype | 40.4203 | 42.7446 | Turkey | leg. G.J. van Pelt | MK779808 | MK861450 |
| boltonii-group | kalkmani sp. nov. | was picta, female paratype | 39.0594 | 43.7540 | Turkey | leg. V. Kalkman | MK779809 | MK861449 |
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 41.6319 | 42.5680 | Georgia | leg. T. Schneider | ||
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 43.4462 | 41.7357 | Russia | leg. O. Kosterin | MN623222 | no data |
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 40.6797 | 37.9573 | Turkey | leg. G.J. van Pelt | MK779841 | MK861493 |
| bidentata-group | mzymtae | was sometimes treated as ssp. of insignis | 40.6797 | 37.9573 | Turkey | leg. G.J. van Pelt | MK779842 | MK861492 |
| boltonii-group | picta | 41.5631 | 24.9664 | Bulgaria | leg. G.J. van Pelt | no data | no data | |
| boltonii-group | picta | morphology only | 42.1667 | 42.9833 | Georgia | leg. T. Schneider | ||
| boltonii-group | picta | 41.3093 | 26.1148 | Greece | Froufe et al., 2014 | KF584944 | KF584992 | |
| boltonii-group | picta | 41.3093 | 26.1148 | Greece | Froufe et al., 2014 | KF584944 | KF584993 | |
| boltonii-group | picta | 40.6910 | 24.6209 | Greece | leg. T. Schneider | MK779802 | no data | |
| boltonii-group | picta | 37.7094 | 26.8178 | Greece | leg. T. Schneider | MW353684 | no data | |
| boltonii-group | picta | 44.6541 | 37.9347 | Russia | leg. O. Kosterin | MK779799 | MK861448 | |
| boltonii-group | picta | 44.6541 | 37.9347 | Russia | leg. O. Kosterin | MK779800 | MK861447 | |
| boltonii-group | picta | 44.6541 | 37.9347 | Russia | leg. O. Kosterin | MK779801 | no data | |
| boltonii-group | picta | 39.4200 | 29.9857 | Turkey | leg. G.J. van Pelt | MK779803 | no data | |
| boltonii-group | picta | 39.6602 | 35.8823 | Turkey | leg. G.J. van Pelt | MK779804 | MK861445 | |
| boltonii-group | picta | 41.6281 | 27.5122 | Turkey | leg. G.J. van Pelt | no data | MK861446 | |
| boltonii-group | vanbrinkae | 38.6825 | 48.7831 | Azerbaijan | leg. N. Snegovaya | MK779806 | no data | |
| boltonii-group | vanbrinkae | morphology only | 38.6825 | 48.7831 | Azerbaijan | leg. N. Snegovaya | ||
| boltonii-group | vanbrinkae | 36.0965 | 53.2606 | Iran | leg. T. Schneider | MW353704 | MW363796 | |
| boltonii-group | vanbrinkae | 36.8741 | 54.8872 | Iran | leg. T. Schneider | MK779805 | MK861442 | |
| boltonii-group | vanbrinkae | larva | 36.5567 | 51.5231 | Iran | leg. M. Waldhauser | MK779807 | MK861443 |
| boltonii-group | vanbrinkae | morphology only | 36.4700 | 51.5383 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae | morphology only | 36.4700 | 51.5383 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae | morphology only | 36.8741 | 54.8872 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae | morphology only | 36.8741 | 54.8872 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae | morphology only | 36.8741 | 54.8872 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae | morphology only | 36.8741 | 54.8872 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae | morphology only | 36.8741 | 54.8872 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae | morphology only | 36.8741 | 54.8872 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae | morphology only | 36.8741 | 54.8872 | Iran | leg. T. Schneider | ||
| boltonii-group | vanbrinkae/picta | hybrid: vanbrinkae × picta | 39.3334 | 46.3786 | Armenia | leg. V. Ananian | MK779798 | MK861444 |
| boltonii-group | vanbrinkae/picta? | morphology only | 39.3106 | 46.3775 | Armenia | leg. V. Ananian | ||
| boltonii-group | vanbrinkae/picta? | morphology only | 39.3106 | 46.3775 | Armenia | leg. V. Ananian |
| Actual Used Names and Taxonomic Rank | Revised Name and Taxonomic Rank | Origin of Specimens Studied |
|---|---|---|
| boltonii-group | ||
| C. heros | C. heros | Bulgaria, Montenegro, Greece, Austria |
| C. picta | C. picta | Bulgaria, Turkey, Greece, Georgia, Russia |
| C. picta | C. kalkmani sp.nov. | Eastern Anatolia Turkey |
| C. vanbrinkae | C. vanbrinkae | Armenia, Azerbaijan, Iran |
| bidentata-group | ||
| C. insignis | C. insignis | West Turkey, East Greece |
| charpentieri-complex | ||
| C. insignis amasina | C. amasina stat. rev. | Turkey |
| C. insignis mzymtae sometimes also used on species level C. mzymtae | C. mzymtae | Turkey, Georgia, Russia |
| C. insignis charpentieri | C. charpentieri stat. rev. | Armenia, Turkey, Georgia |
| C. insignis lagodechica | C. charpentieri stat. rev. C. insignis lagodechica syn. nov. | Georgia |
| C. insignis nobilis | C. charpentieri stat. rev. C. insignis nobilis syn. nov. | Turkey, Iran |
| C. nachitschevanica | C. charpentieri stat. rev. C. nachitschevanica syn. nov. | Azerbaijan |
| C. plagionyx | C. charpentieri stat. rev. C. plagionyx syn. nov. | Azerbaijan |
| C. insignis | C. cilicia sp. nov. | Eastern Mediterranean Turkey, Lebanon |
| coronata-group | ||
| C. insignis coronata sometimes used on species level C. coronata | C. coronata | Iran, Kyrgyzstan |
| C. boltonii | C. trinacriae | C. princeps | C. heros | C. picta | C. kalkmani | C. vanbrinkae | Average Distance within the Group | |
|---|---|---|---|---|---|---|---|---|
| C. boltonii | 0.0077 | |||||||
| C. trinacriae | 0.0583 | 0.0048 | ||||||
| C. princeps | 0.0563 | 0.0632 | 0.0044 | |||||
| C. heros | 0.0811 | 0.0865 | 0.0852 | 0.0151 | ||||
| C. picta | 0.0790 | 0.0794 | 0.0865 | 0.0617 | 0.0060 | |||
| C. kalkmani | 0.0698 | 0.0884 | 0.0718 | 0.0625 | 0.0588 | 0.0036 | ||
| C. vanbrinkae | 0.0791 | 0.0838 | 0.0941 | 0.0537 | 0.0395 | 0.0493 | 0.0044 | |
| C. coronata | 0.1108 | 0.1083 | 0.1083 | 0.0883 | 0.0950 | 0.1010 | 0.0991 | 0.0402 |
| C. bidentata | C. insignis | C. amasina | C. mzymtae | C. charpentieri | C. cilicia | Average Distance within the Group | |
|---|---|---|---|---|---|---|---|
| C. bidentata | 0.0098 | ||||||
| C. insignis | 0.0686 | 0.0093 | |||||
| C. amasina | 0.0656 | 0.0623 | 0.0012 | ||||
| C. mzymtae | 0.0590 | 0.0551 | 0.0296 | 0.0012 | |||
| C. charpentieri | 0.0633 | 0.0633 | 0.0452 | 0.0424 | 0.0110 | ||
| C. cilicia | 0.0566 | 0.0628 | 0.0401 | 0.0421 | 0.0322 | 0.0067 | |
| C. coronata | 0.1028 | 0.0972 | 0.1022 | 0.0966 | 0.0942 | 0.0888 | 0.0402 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schneider, T.; Vierstraete, A.; Müller, O.; van Pelt, G.J.; Caspers, M.; Ikemeyer, D.; Snegovaya, N.; Dumont, H.J. Taxonomic Revision of Eastern Part of Western Palaearctic Cordulegaster Using Molecular Phylogeny and Morphology, with the Description of Two New Species (Odonata: Anisoptera: Cordulegastridae). Diversity 2021, 13, 667. https://doi.org/10.3390/d13120667
Schneider T, Vierstraete A, Müller O, van Pelt GJ, Caspers M, Ikemeyer D, Snegovaya N, Dumont HJ. Taxonomic Revision of Eastern Part of Western Palaearctic Cordulegaster Using Molecular Phylogeny and Morphology, with the Description of Two New Species (Odonata: Anisoptera: Cordulegastridae). Diversity. 2021; 13(12):667. https://doi.org/10.3390/d13120667
Chicago/Turabian StyleSchneider, Thomas, Andy Vierstraete, Ole Müller, Gert Jan van Pelt, Max Caspers, Dietmar Ikemeyer, Nataly Snegovaya, and Henri J. Dumont. 2021. "Taxonomic Revision of Eastern Part of Western Palaearctic Cordulegaster Using Molecular Phylogeny and Morphology, with the Description of Two New Species (Odonata: Anisoptera: Cordulegastridae)" Diversity 13, no. 12: 667. https://doi.org/10.3390/d13120667
APA StyleSchneider, T., Vierstraete, A., Müller, O., van Pelt, G. J., Caspers, M., Ikemeyer, D., Snegovaya, N., & Dumont, H. J. (2021). Taxonomic Revision of Eastern Part of Western Palaearctic Cordulegaster Using Molecular Phylogeny and Morphology, with the Description of Two New Species (Odonata: Anisoptera: Cordulegastridae). Diversity, 13(12), 667. https://doi.org/10.3390/d13120667