
Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses?


Abstract
:1. Introduction
2. Material and Methods
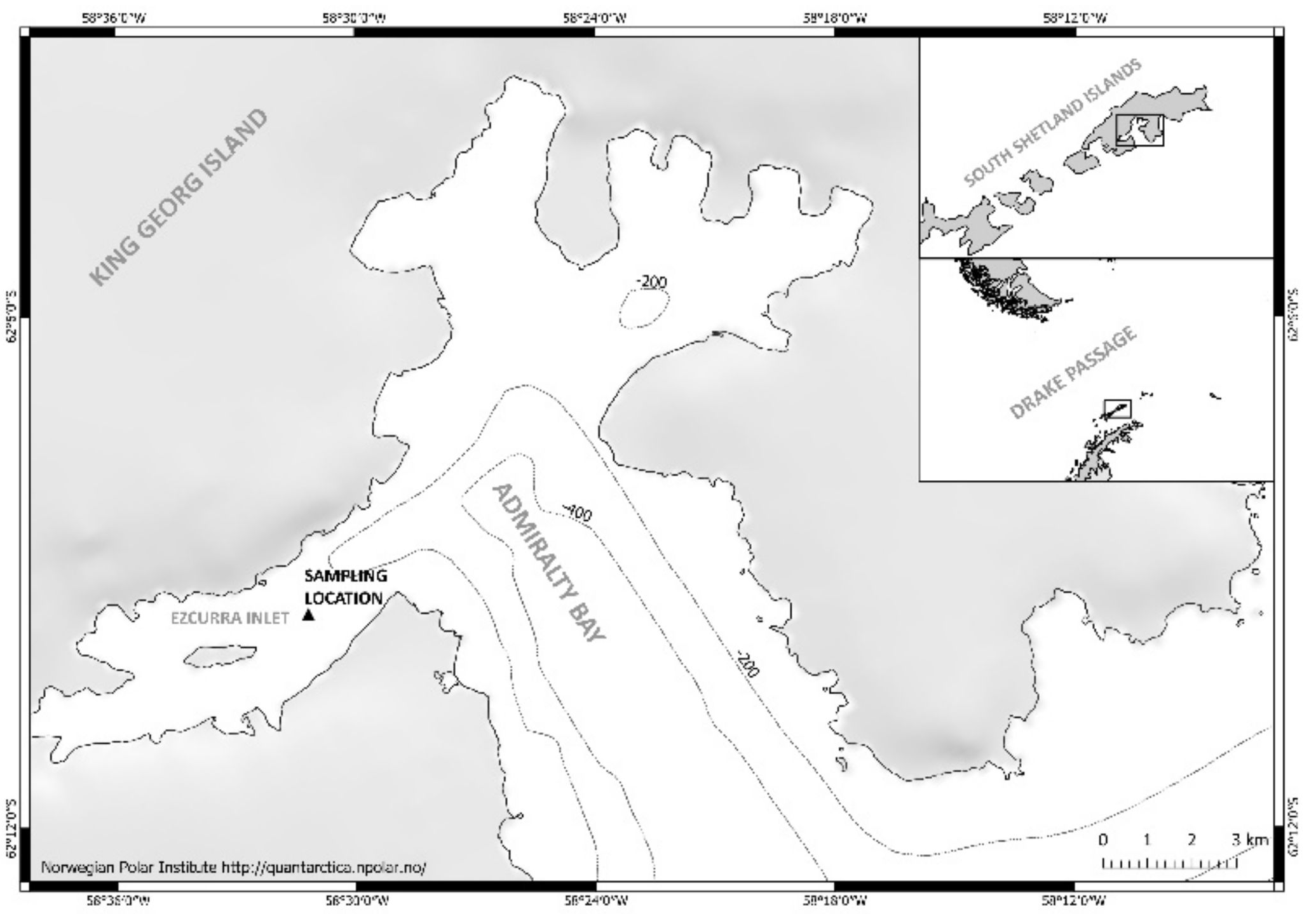
2.1. Study Area
2.2. Sample Collection and Laboratory Studies
2.3. Statistical Analyses
3. Results
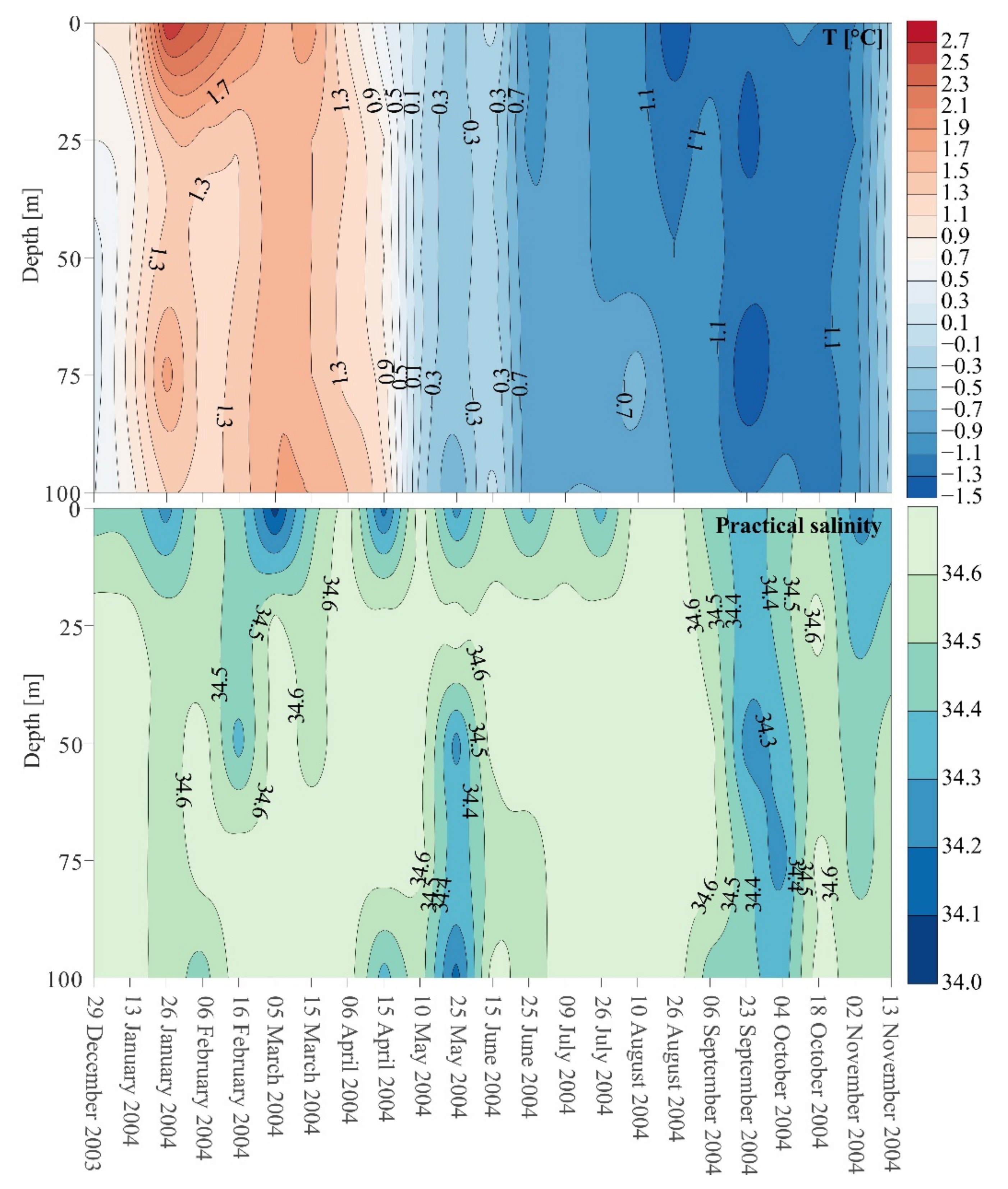
3.1. Hydrological Conditions
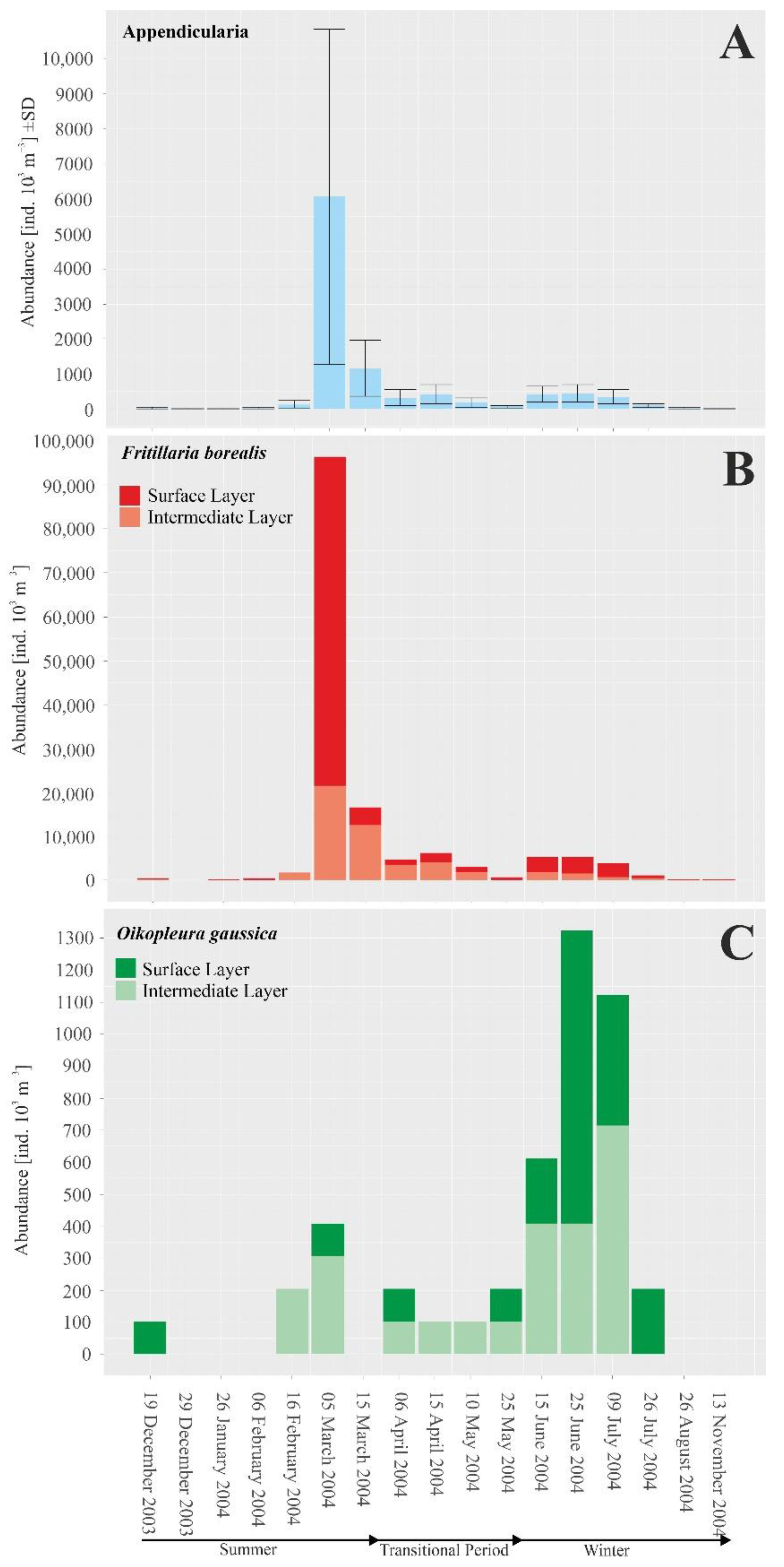
3.2. Appendicularian Diversity and Abundance—Annual Changes
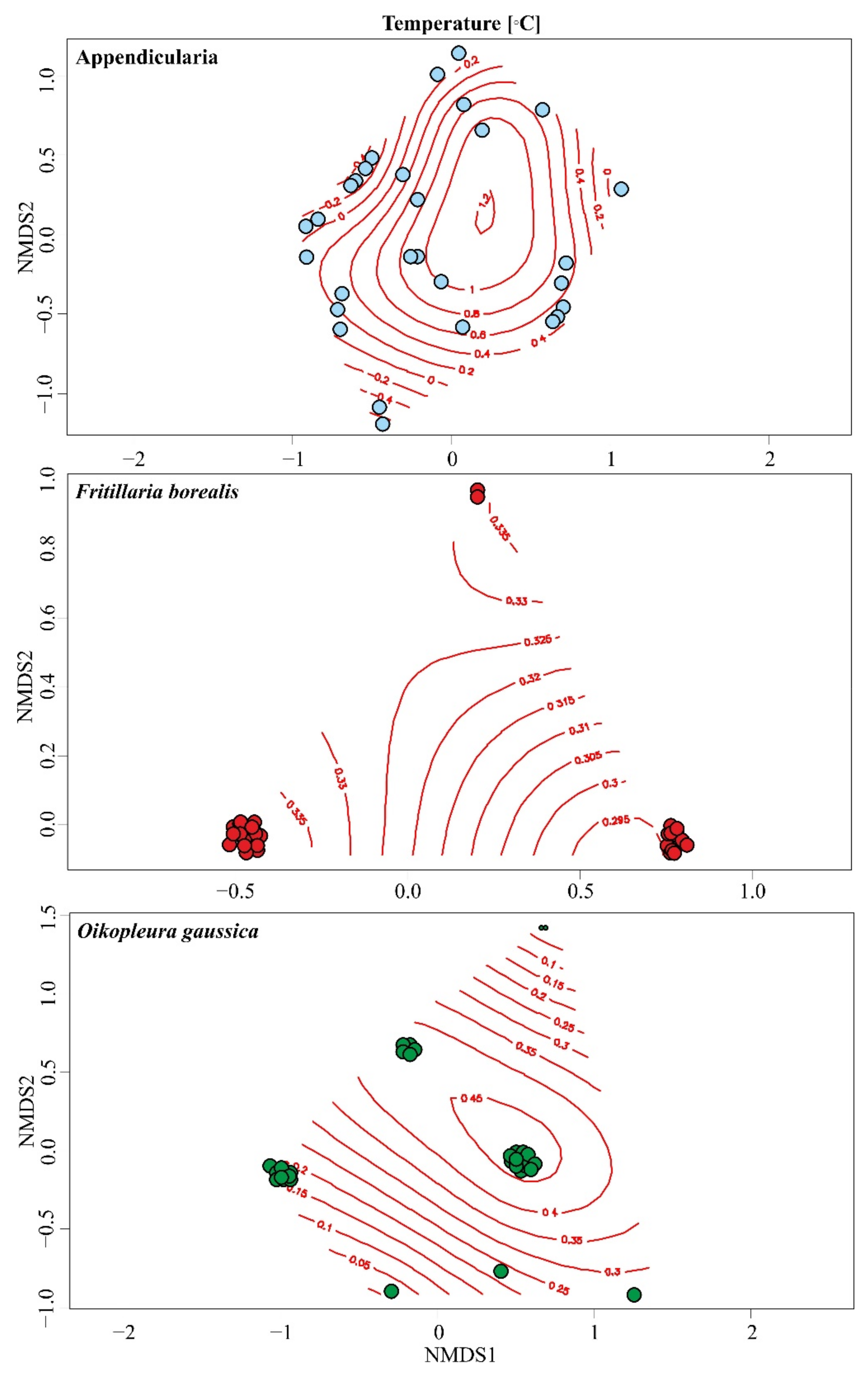
3.3. Environmental Factors Influencing the Annual Changes in the Abundance of Appendicularia
4. Discussion
4.1. Appendicularians Diversity and Abundance
4.2. Environmental Factors Influencing the Abundance of Appendicularia
4.3. Appendicularia (Tunicata) in an Antarctic Glacial Fjord vs. Open Oceanic Waters
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purcell, J.E. Predation by the hydromedusa Aequorea victoria on fish larvae and eggs at a herring spawning ground in British Columbia. Can. J. Fish. Aquat. Sci. 1989, 46, 1415–1427. [Google Scholar] [CrossRef]
- Purcell, J.E.; Sturdevant, M.V. Prey selection and dietary overlap among zooplanktivorous jellyfish and juvenile fishes in Prince William Sound, Alaska. Mar. Ecol. Prog. Ser. 2001, 210, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Di Mauro, R.; Capitanio, F.; Viñas, M.D. Capture efficiency for small dominant mesozooplankters (Copepoda, Appendicularia) off Buenos Aires Province (34° S–41° S), Argentine Sea, using two plankton mesh sizes. Braz. J. Oceanogr. 2009, 57, 205–214. [Google Scholar] [CrossRef]
- Landry, M.R.; Peterson, W.K.; Fagerness, V.L. Mesozooplankton grazing in the Southern California Bight. I. Population abundances and gut pigment contents. Mar. Ecol. Prog. Ser. 1994, 115, 55–71. [Google Scholar] [CrossRef]
- Fernández, D.; López-Urrutia, A.; Fernández, A.; Acuña, J.L.; Harris, R. Retention efficiency of 0.2 to 6 μm particles by the appendicularians Oikopleura dioica and Fritillaria borealis. Mar. Ecol. Prog. Ser. 2004, 266, 89–101. [Google Scholar] [CrossRef] [Green Version]
- López-Urrutia, A.; Acuña, J.L. Gut throughput dynamics in the appendicularian Oikopleura dioica. Mar. Ecol. Prog. Ser. 1999, 191, 195–205. [Google Scholar] [CrossRef]
- Alldredge, A.L. The contribution of discarded appendicularian houses to the flux of particulate organic carbon from oceanic surface waters. In Response of Marine Ecosystems to Global Change: Ecological Impact of Appendicularians; Gorsky, G., Youngbluth, M.J., Deibel, D., Eds.; GB Scientific Publisher: Paris, France, 2005; pp. 309–326. [Google Scholar]
- Winder, M.; Bouquet, J.-M.; Bermúdez, J.R.; Berger, S.A.; Hansen, T.; Brandes, J.; Sazhin, A.F.; Nejstgaard, J.C.; Båmstedt, U.; Jakobsen, H.H.; et al. Increased appendicularian zooplankton alter carbon cycling under warmer more acidified ocean conditions. Limnol. Oceanogr. 2017, 62, 1541–1551. [Google Scholar] [CrossRef]
- Vargas, C.A.; Tönnesson, K.; Sell, A.; Maar, M.; Friis Møller, E.; Zervoudaki, T.; Giannakourou, A.; Christou, E.; Satapoomin, S.; Petersen, J.K.; et al. Importance of copepods versus appendicularians in vertical carbon fluxes in a Swedish fjord. Mar. Ecol. Prog. Ser. 2002, 241, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Robison, B.H.; Reisenbichler, K.R.; Sherlock, R.E. Giant larvacean houses: Rapid carbon transport to the deep seafloor. Science 2005, 308, 1609–1611. [Google Scholar] [CrossRef]
- Capitanio, F.L.; Spinelli, M.L.; Presta, M.L.; Aguirre, G.E.; Cervetto, G.; Pájaro, M.; Derisio, C.M. Ecological Role of Common Appendicularian Species from Shelf Waters Off Argentina. In Plankton Ecology of the Southwestern Atlantic; Hoffmeyer, M.S., Sabatini, M.E., Brandini, F.P., Calliari, D.L., Santinelli, N.H., Eds.; Springer International Publishing AG, Springer Nature: Cham, Switzerland, 2018; pp. 201–218. [Google Scholar]
- Capitanio, F.; Pájaro, M.; Esnal, G.B. Appendicularians (Chordata, Tunicata) in the diet of anchovy Engraulis anchoita in the Argentine sea. Sci. Mar. 1997, 61, 9–15. [Google Scholar]
- Capitanio, F.; Pájaro, M.; Esnal, G.B. Appendicularians: An important food supply for the Argentine anchovy Engraulis anchoita in coastal waters. J. Appl. Ichthyol. 2005, 21, 414–419. [Google Scholar] [CrossRef]
- Lombard, F.; Legendre, L.; Picheral, M.; Sciandra, A.; Gorsky, G. Prediction of ecological niches and carbon export by appendicularians using a new multispecies ecophysiological model. Mar. Ecol. Prog. Ser. 2010, 98, 109–125. [Google Scholar] [CrossRef]
- Fenaux, R. Anatomy and functional morphology of the Appendicularia. In The Biology of Pelagic Tunicates; Bone., Q., Ed.; Oxford University Press: New York, NY, USA, 1998; pp. 25–34. [Google Scholar]
- Kalarus, M.; Panasiuk, A. Spatial Distribution Patterns of Appendicularians in the Drake Passage: Potential Indicators of Water Masses? Diversity 2021, 13, 286. [Google Scholar] [CrossRef]
- Hopcroft, R.R.; Roff, J.C.; Chavez, F. Size paradigms in copepod communities: A re-examination. Hydrobiologia 2001, 453, 133–141. [Google Scholar] [CrossRef]
- Harris, R.P.; Lopez-Urrutia, A.; Acuna, J.L.; Båmstedt, U.; Flood, P.R.; Fhyn, H.J.; Gorsky, G.; Youngbluth, M.J.; Irigoien, I.; Martinussen, M. A comparison of appendicularian seasonal cycles in four distinct European coastal environments. In Response of Marine Ecosystems to Global Change: Ecological Impact of Appendicularians; Gorsky, G., Youngbluth, M.J., Deibel, D., Eds.; Editions des Archives Contemporaires; GB Scientific Publisher: Paris, France, 2005; pp. 255–276. [Google Scholar]
- Hunt, B.P.V.; Hosie, G.W. Zonal structure of zooplankton communities in the Southern Ocean South of Australia: Results from a 2150 km continuous plankton recorder transect. Deep-Sea Res. Part I 2005, 52, 1241–1271. [Google Scholar] [CrossRef]
- Tsujimoto, M.; Takahashi, K.T.; Hirawake, T.; Mitsuo, F. Unusual abundance of appendicularians in the seasonal ice zone (140° E) of the Southern Ocean. Polar Biosc. 2006, 19, 133–141. [Google Scholar]
- Daponte, M.; Esnal, G. The classification of Antarctic appendicularians: The Oikopleura gaussica group. Ant. Sci. 2003, 15, 476–482. [Google Scholar]
- Hosie, G.W.; Schultz, M.B.; Kitchener, J.A.; Cochran, T.G.; Richards, K. Macrozooplankton community structure off East Antarctica (80–150° E) during the Austral summer of 1995/1996. Deep Sea Res. Part II 2000, 47, 2437–2463. [Google Scholar] [CrossRef]
- Hosie, G.W.; Fukuchi, M.; Kawaguchi, S. Development of the Southern Ocean Continuous Plankton Recorder survey. Prog. Oceanogr. 2003, 58, 263–283. [Google Scholar] [CrossRef]
- Takahashi, K.T.; Hosie, G.W.; Kitchener, J.A.; McLeod, D.J.; Odate, T.; Fukuchi, M. Comparison of zooplankton distribution patterns between four seasons in the Indian Ocean sector of the Southern Ocean. Polar Sci. 2010, 4, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Ojima, M.; Takahashi, K.T.; Tanimura, A.; Odate, T.; Fukuchi, M. Spatial distribution of micro- and meso-zooplankton in the seasonal ice zone of east Antarctica during 1983–1995. Polar Sci. 2015, 9, 319–326. [Google Scholar] [CrossRef]
- Fuentes, V.L.; Schnack-Schiel, S.B.; Schloss, I.R.; Esnal, G.G. Mesozooplankton of Potter Cove: Community composition and seasonal distribution in 2002 and 2003, In The Antarctic Ecosystem of Potter Cove, King-George Island (Isla 25 de Mayo), Synopsis of Research Performed 1999–2006 at the Dallmann Laboratory and Jubany Station; Wiencke, C., Ferreyra, G.A., Abele, D., Marenssi, S., Eds.; Reports on Polar and Marine Research; Alfred-Wegener-Institut Für Polar- und Meeresforschung: Bremerhaven, Germany, 2008; Volume 571, pp. 75–84. [Google Scholar]
- Freire, A.S.; Coelho, M.J.C.; Bonecker, S.L.C. Short term spatial—Temporal distribution patterns of zooplankton in Admiralty Bay (Antarctica). Polar Biol. 1993, 13, 433–439. [Google Scholar] [CrossRef]
- Potocka, M.; Kidawa, A.; Panasiuk, A.; Bielecka, L.; Wawrzynek-Borejko, J.; Patuła, W.; Wójcik, K.A.; Plenzler, J.; Janecki, T.; Bialik, R.J. The effect of glacier recession on benthic and pelagic communities: Case study in Herve Cove, Antarctica. J. Mar. Sci. Eng. 2019, 7, 285. [Google Scholar] [CrossRef] [Green Version]
- Mańko, M.K.; Panasiuk-Chodnicka, A.A.; Żmijewska, M.I. Pelagic coelenterates in the Atlantic sector of the Arctic Ocean: Species diversity and distribution as water mass indicators. Oceanol. Hydrobiol. Stud. 2015, 44, 466–479. [Google Scholar] [CrossRef]
- Słomska, A.W.; Panasiuk-Chodnicka, A.A.; Żmijewska, M.I.; Mańko, M.K. Variability of Salpa thompsoni population structure in the Drake Passage in summer season 2010. Pol. Polar Res. 2015, 36, 391–404. [Google Scholar] [CrossRef]
- La, H.S.; Park, K.; Wåhlin, A.; Arrigo, K.R.; Kim, D.S.; Yang, E.J.; Atkinson, A.; Fielding, S.; Im, J.; Kim, T.-W.; et al. Zooplankton and micronekton respond to climate fluctuations in the Amundsen Sea polynya, Antarctica. Sci. Rep. 2019, 9, 10087. [Google Scholar] [CrossRef]
- Panasiuk, A.; Grzonka, L.; Prątnicka, P.; Wawrzynek-Borejko, J.; Szymelfenig, M. Zonal variability of pelagic Siphonophora (Cnidaria) in the Atlantic sector of the southern ocean. J. Sea Res. 2020, 165, 101951. [Google Scholar] [CrossRef]
- Birkenmajer, K. Retreat of Ecology Glacier, Admiralty Bay. King George Island (South Shetland Islands, West Antarctica), 1956–2001. Bull. Pol. Acad. Sci. 2002, 50, 15–30. [Google Scholar]
- Simões, J.C.; Dani, N.; Bremer, U.F.; Aquino, F.E.; Arigony-Neto, J. Small cirque glaciers retreat on Keller Peninsula, Admiralty Bay, King George Island, Antarctica. Pesqui. Antart. Bras. 2004, 4, 49–56. [Google Scholar]
- Pudełko, R.; Angiel, P.J.; Potocki, M.; Jędrejek, A.; Kozak, M. Fluctuation of Glacial Retreat Rates in the Eastern Part of Warszawa Icefield, King George Island, Antarctica, 1979–2018. Remote Sens. 2018, 10, 892. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.D.; Fernández Severini, M.D.; Spetter, C.; López Abbatea, M.C.; Tartara, M.N.; Nahuelhual, E.G.; Marcovecchio, J.E.; Schloss, I.R.; Hoffmeyer, M.S. Effects of glacier melting on the planktonic communities of two Antarctic coastal areas (Potter Cove and Hope Bay) in summer. Reg. Stud. Mar. Sci. 2019, 30, 100731. [Google Scholar] [CrossRef]
- Garcia, M.D.; Dutto, M.S.; Chazarreta, C.J.; Berasategui, A.A.; Schloss, I.R.; Hoffmeyer, M.S. Micro-mesozooplankton successions in an Antarctic coastal environment during a warm year. PLoS ONE 2020, 15, e0232614. [Google Scholar] [CrossRef] [PubMed]
- Antezana, T. Plankton of Southern Chilean fjords: Trends and linkages. Sci. Mar. 1999, 63 (Suppl. S1), 69–80. [Google Scholar] [CrossRef]
- Rakusa-Suszczewski, S. The Maritime Antarctic Coastal Ecosystem of Admiralty Bay; Department of Antarctic Biology, Polish Academy of Sciences: Warsaw, Poland, 1993; pp. 1–216. [Google Scholar]
- Rakusa-Suszczewski, S. The hydrography of Admiralty Bay and its inlets, coves and lagoons (King George Island, Antarctica). Pol. Polar Res. 1995, 17, 29–42. [Google Scholar]
- Wright, F.F. Fiord circulation and sedimentation Southeast Alaska. In Proceedings of the First International Conference of Port Ocean Engineering Under Arctic Conditions, Trondheim, Norway, 23–30 August 1971; Volume 1, pp. 279–289. [Google Scholar]
- Rakusa-Suszczewski, S. The role of near shore research in gaining an understanding of the functioning of the Antarctic ecosystem. Pol. Arch. Hydrobiol. 1980, 27, 229–233. [Google Scholar]
- Tokarczyk, R. Classification of water masses in the Bransfield Strait and southern part of the Drake Passage using a method of statistical multidimensional analysis. Pol. Polar Res. 1987, 8, 333–366. [Google Scholar]
- Szafrański, Z.; Lipski, M. Characteristics of water temperature and salinity at Admiralty Bay (King George Island, South Shetlands Islands, Antarctic) during the austral summer 1978/1979. Pol. Polar Res. 1982, 3, 7–24. [Google Scholar]
- Pęcherzewski, K. Distribution and quantity of suspended matter in Admiralty Bay (king George Island, South Shetland Islands). Pol. Polar Res. 1980, 1, 75–82. [Google Scholar]
- Orsi, H.; Whitworth, T.; Nowlin, W.D., Jr. On the meridional extent and fronts of the Antarctic Circumpolar Current. Deep Sea Res. Part I 1995, 42, 641–673. [Google Scholar] [CrossRef]
- Martinson, D.G.; Stammerjohn, S.E.; Iannuzzi, R.A.; Smith, R.C.; Vernet, M. Western Antarctic Peninsula physical oceanography and spatio–temporal variability. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 1964–1987. [Google Scholar] [CrossRef]
- Cape, M.R.; Vernet, M.; Pettit, E.C.; Wellner, J.; Truffer, M.; Akie, G.; Domack, E.; Leventer, A.; Smith, C.R.; Huber, B.A. Circumpolar Deep Water Impacts Glacial Meltwater Export and Coastal Biogeochemical Cycling Along the West Antarctic Peninsula. Front. Mar. Sci. 2019, 6, 144. [Google Scholar] [CrossRef]
- Moffat, C.; Meredith, M. Shelf–ocean exchange and hydrography west of the Antarctic Peninsula: A review. Philos. Trans. A Math. Phys. Eng. Sci. 2018, 376, 20170164. [Google Scholar] [CrossRef]
- Esnal, G. Apendicularia. In Atlas del Zooplancton del Atlántico Sudoccidental y Métodosde Trabajo con el Zooplancton Marino; Boltovskoy, D., Ed.; Publicación Especial, Instituto Nacional de Investigacióny Desarrollo Pesquero: Mar del Plata, Argentina, 1981; pp. 809–820. [Google Scholar]
- Esnal, G. Appendicularia. In South Atlantic Zooplankton; Boltovskoy, D., Ed.; Backhuys Publication: Leiden, The Netherlands, 1999; pp. 1375–1399. [Google Scholar]
- Fenaux, R. The Classification of Appendicularia (Tunicata): History and Current State. Mem. Inst. Oceanogr. 1993, 17, 1–123. [Google Scholar]
- O’Sullivan, D. A Guide to the Pelagic Tunicates of the Southern Ocean and Adjacent Waters; ANARE Research Notes; Antarctic Division: Kingston, Australia, 1983.
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001; pp. 1–176. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; pp. 1–190. [Google Scholar]
- Vanables, W.N.; Smith, D.M.; R Core Team. An Introduction to R, Notes on R: A Programming Environment for Data Analysis and GraphicsVersion 4.1.0. Available online: https://cran.r-project.org/doc/manuals/r-release/R-intro.pdf (accessed on 10 November 2021).
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Henry, M.; Stevens, H. The vegan package. Community Ecol. Package 2013, 10, 631–637. [Google Scholar]
- Bray, J.B.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Spinelli, M.L.; Franzosi, C.; Olguin Salinas, H.; Capitanio, F.L.; Alder, V.A. Appendicularians and copepods from Scotia Bay (Laurie island, South Orkney, Antarctica): Fluctuations in community structure and diversity in two contrasting, consecutive summers. Polar Biol. 2017, 41, 663–678. [Google Scholar] [CrossRef]
- Gorsky, G.; Youngbluth, M.J.; Deibel, D. Response of Marine Eco-Systems to Global Change: Ecological Impact of Appendicularians; Editions Scientifiques; GB Scientific Publisher: Paris, France, 2005; pp. 1–440. [Google Scholar]
- Atkinson, A.; Ward, P.; Hunt, B.P.V.; Pakhomov, E.A.; Hosie, G.W. An overview of South-ern Ocean zooplankton data: Abundance, biomass, feeding and functional relationships. CCAMLR Sci. 2012, 19, 171–218. [Google Scholar]
- Krajewska, M.; Szymczak-Żyła, M.; Tylmann, W.; Kowalewska, G. Climate change impact on primary production and phytoplankton taxonomy in Western Spitsbergen fjords based on pigments in sediments. Glob. Planet. Chang. 2020, 189, 103158. [Google Scholar] [CrossRef]
- Kittel, W.; Siciński, J.; Żmijewska, M.I.; Bielecka, L.; Błachowiak-Samołyk, K. Antarctic neritic zooplankton community (Admiralty Bay, King George Island, South Shetland Island). Pol. Polar Res. 2001, 22, 11–33. [Google Scholar]
- Acuña, J.-L.; Anadón, R. Appendicularian assemblages in a shelf area and their relationship with temperature. J. Plankton Res. 1992, 14, 1233–1250. [Google Scholar] [CrossRef]
- Ligowski, R. Net phytoplankton of the Admiralty Bay (King George Island, South Shetland Island) in 1983. Pol. Polar Res. 1986, 7, 127–154. [Google Scholar]
- Woźniak, B.; Hapter, R.; Maj, B. The inflow of solar Energy and the irradiance of the euphotic zone in the region of Ezcurra Inlet during the antarctica summer of 1977/78. Oceanologia 1983, 15, 141–173. [Google Scholar]
- Wasiłowska, A.; Kopczyńska, E.E.; Rzepecki, M. Temporal and spatial variation of phytoplankton in Admiralty Bay, South Shetlands: The dynamics of summer blooms shown by pigment and light microscopy analysis. Polar Biol. 2015, 38, 1249–1265. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, M.C.M.; Williams, G.D. Distribution and abundance of Larvaceans in the Southern Ocean between 30 and 80° E. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 905–915. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Fb | Fa | Fan | F | Of | Op | Og | O | App |
|---|---|---|---|---|---|---|---|---|---|
| S | 7541.4 ± 18,917.9 | 6.4 ± 25.5 | 0 | 127.4 ± 296.5 | 0 | 6.4 ± 25.5 | 57.3 ± 90.9 | 12.7 ± 34.8 | 968.9 ± 6964.4 |
| TP | 1630.6 ± 1392.9 | 0 | 0 | 84.9 ± 163.3 | 0 | 0 | 67.9 ± 52.6 | 33.9 ± 52.6 | 227.2 ± 705.8 |
| W | 1293.5 ± 1370.1 | 7.8 ± 28.3 | 7.8 ± 28.3 | 62.7 ± 78.2 | 47.1 ± 114.8 | 7.8 ± 28.3 | 250.9 ± 304.6 | 47.1 ± 98.6 | 215.6 ± 637.5 |
| Season | S | TP | W | |||
|---|---|---|---|---|---|---|
| Av. Sim. | 36.14 | 59.70 | 48.05 | |||
| Av. Sim. and Contrib. [%] | ||||||
| Fritillaria borealis | 30.19 | 83.53 | 48.94 | 81.99 | 35.97 | 81.99 |
| Seasons | Summer and TP | Summer and Winter | TP and Winter | |||
| Av. Diss. | 56.14 | 58.56 | 46.36 | |||
| Av. Diss. and Contrib. [%] | ||||||
| Fritillaria borealis | 36.76 | 80.20 | 31.68 | 54.10 | 21.49 | 46.34 |
| Oikopleura gaussica | 7.38 | 13.14 | 10.17 | 17.36 | 10.17 | 21.93 |
| Ef | Df R | Fb | Fa | Fan | F | Of | Op | Og | O | App |
|---|---|---|---|---|---|---|---|---|---|---|
| Season | 1 33 | 0.196 | 1 | 0.37 | 0.345 | 0.14 | 1 | 0.189 | 0.439 | 0.099 |
| Season + T | 1 1 32 | 0.165 0.046 * | 0.798 0.534 | 0.345 0.947 | 0.345 0.431 | 0.034 * 0.359 | 0.766 0.491 | 0.223 0.889 | 0.458 0.756 | 0.066 0.012 * |
| T | 1 33 | 0.148 | 0.823 | 0.422 | 0.257 | 0.336 | 0.861 | 0.359 | 0.65 | 0.069 |
| S | 1 33 | 0.605 | 0.323 | 1 | 0.515 | 0.095 | 0.344 | 0.318 | 0.495 | 0.311 |
| T + S | 1 1 32 | 0.170 0.626 | 0.799 0.323 | 0.411 0.816 | 0.273 0.629 | 0.323 0.076 | 0.812 0.321 | 0.370 0.239 | 0.649 0.437 | 0.063 0.386 |
| Depth | 1 33 | 0.997 | 0.207 | 0.477 | 0.53 | 0.239 | 0.242 | 0.943 | 0.407 | 0.964 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panasiuk, A.; Kalarus, M. Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses? Diversity 2021, 13, 675. https://doi.org/10.3390/d13120675
Panasiuk A, Kalarus M. Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses? Diversity. 2021; 13(12):675. https://doi.org/10.3390/d13120675
Chicago/Turabian StylePanasiuk, Anna, and Marcin Kalarus. 2021. "Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses?" Diversity 13, no. 12: 675. https://doi.org/10.3390/d13120675
APA StylePanasiuk, A., & Kalarus, M. (2021). Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses? Diversity, 13(12), 675. https://doi.org/10.3390/d13120675