
Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site?


Abstract
:1. Introduction
2. Materials and Methods
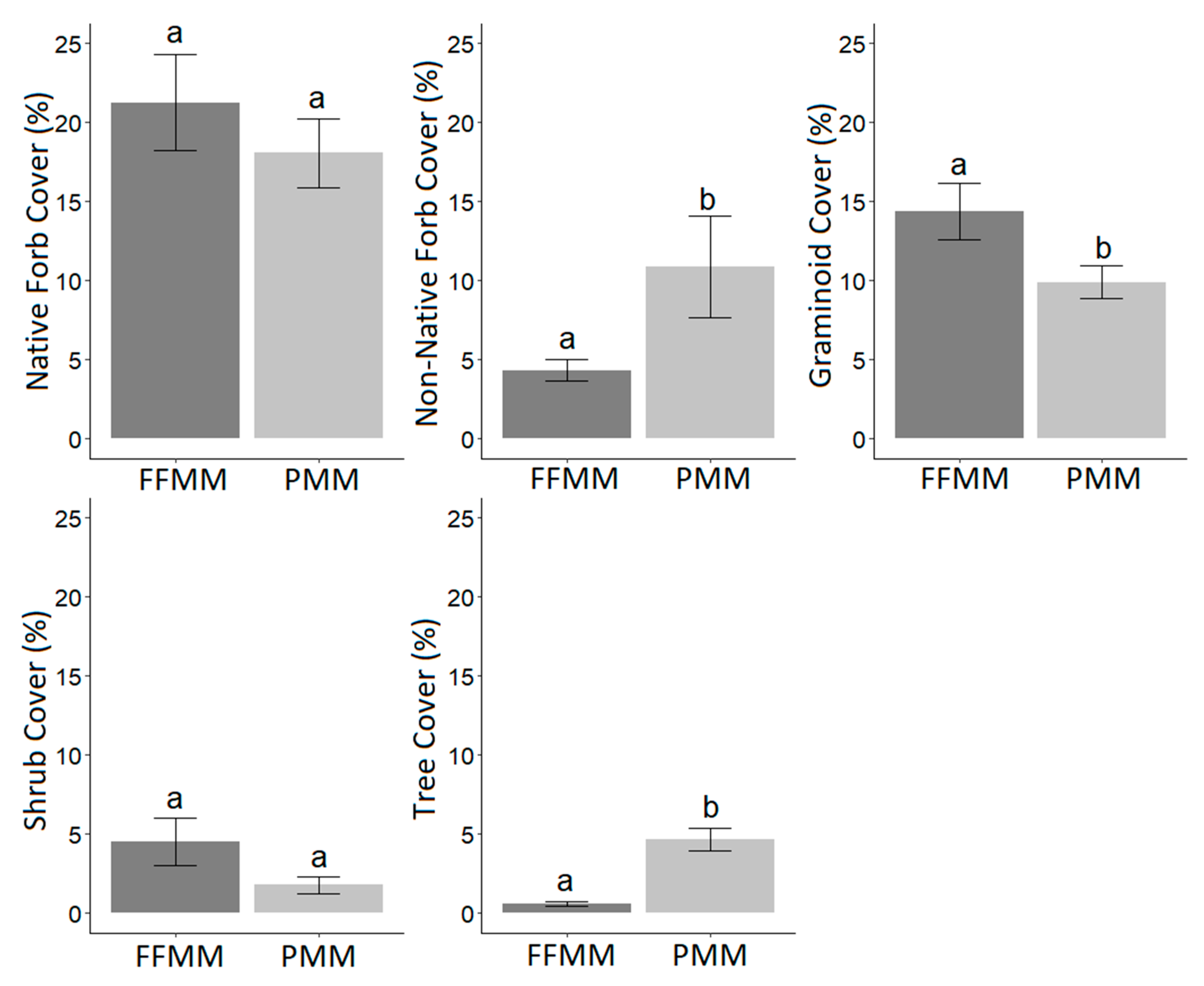
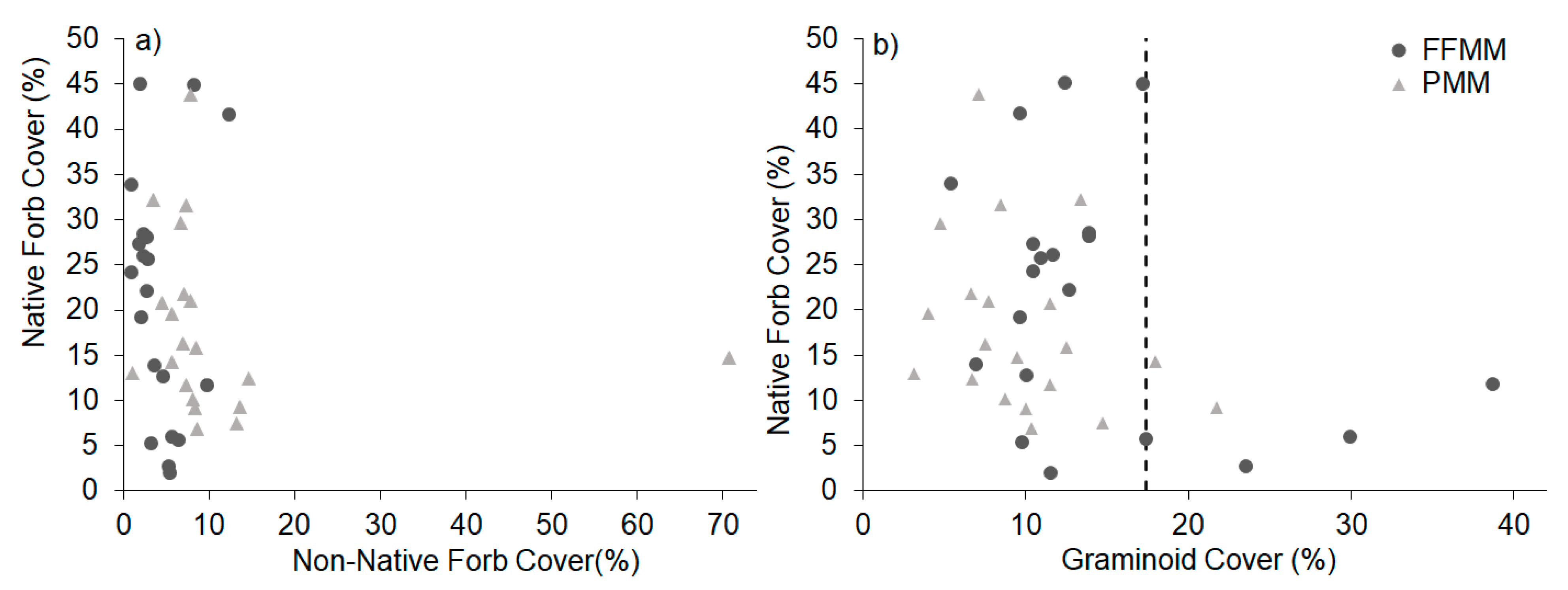
3. Results
4. Discussion
5. Management Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cole, D.E.; King, J.R.; Oyarzun, D.A.; Dietzler, T.H.; McClay, A. Experiences with invasive plant management and ecology in Alberta. Can. J. Plant Sci. 2007, 87, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Small, C.; Degenhardt, D.; Drozdowski, B.; Thacker, S.; Powter, C.; Schoonmaker, A.; Schreiber, S. Optimizing Weed Control for Progressive Reclamation: Literature Review; Canada’s Oil Sands Innovation Alliance: Calgary, AB, Canada, 2018; 48p. [Google Scholar]
- Luck, L.; Bellairs, S.M.; Rossiter-Rachor, N.A. Residual herbicide treatments reduce Andropogon gayanus (Gamba Grass) recruitment for mine site restoration in northern Australia. Ecol. Manag. Restor. 2019, 20, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Franke, M.E.; Zipper, C.; Barney, J.N. Native hardwood tree seedling establishment following invasive autumn-olive (elaeagnus umbellata) removal on a reclaimed coal mine. Invasive Plant Sci. Manag. 2018, 11, 155–161. [Google Scholar] [CrossRef]
- Holmes, P.M. Shrubland restoration following woody alien invasion and mining: Effects of topsoil depth, seed source, and fertilizer addition. Restor. Ecol. 2001, 9, 71–84. [Google Scholar] [CrossRef]
- González-Muñoz, N.; Costa-Tenorio, M.; Espigares, T. Invasion of alien Acacia dealbata on Spanish Quercus robur forests: Impact on soils and vegetation. For. Ecol. Manag. 2012, 269, 214–221. [Google Scholar] [CrossRef]
- Alberta Environment. Guidelines for Reclamation to Forest Vegetation in the Athabasca Oil Sands Region, 2nd ed.; Terrestrial Subgroup of the Reclamation Working Group of the Cumulative Environmental Management Association: Fort McMurray, AB, Canada, 2010. [Google Scholar]
- Baker, H.G. The evolution of weeds. Annu. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Espeland, E.; Perkins, L. Weed establishment and persistence after water pipeline installation and reclamation in the mixed grass prairie of Western North Dakota. Ecol. Restor. 2017, 35, 303–310. [Google Scholar] [CrossRef]
- Errington, R.C.; Pinno, B.D. Early successional plant community dynamics on a reclaimed oil sands mine in comparison with natural boreal forest communities. Ecoscience 2015, 22, 133–144. [Google Scholar] [CrossRef]
- Mackenzie, D.D.; Naeth, M.A. The role of the forest soil propagule bank in assisted natural recovery after oil sands mining. Restor. Ecol. 2009, 18, 418–427. [Google Scholar] [CrossRef]
- Gingras-Hill, T.; Nwaishi, F.C.; Macrae, M.; Price, J.S.; Petrone, R.M. Ecohydrological functioning of an upland undergoing reclamation on post-mining landscape of the Athabasca oil sands region, Canada. Ecohydrology 2018, 11, 1–12. [Google Scholar] [CrossRef]
- Cavieres, L.; Arroyo, M.T.K.; Peñaloza, A.; Molina-Montenegro, M.; Torres, C. Nurse effect of Bolax gummifera cushion plants in the alpine vegetation of the Chilean Patagonian Andes. J. Veg. Sci. 2002, 13, 547–554. [Google Scholar] [CrossRef]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Jordan, N.; Larson, D.L.; Huerd, S.C. Soil modification by invasive plants: Effects on native and invasive species of mixed-grass prairies. Biol. Invasions 2007, 10, 177–190. [Google Scholar] [CrossRef]
- Maestre, F.T.; Bautista, S.; Cortina, J.; Bellot, J. Potential for using facilitation by grasses to establish shrubs on a semiarid degraded steppe. Ecol. Appl. 2001, 11, 1641–1655. [Google Scholar] [CrossRef]
- Alberta Environment and Water. Best Management Practices for Conservation of Reclamation Materials in the Mineable Oil Sands Region of Alberta; Province of Alberta: Edmonton, AB, Canada, 2012; pp. 1–161. [Google Scholar]
- Natural Regions Committee. Natural Regions and Subregions of Alberta; Natural Regions Committee: Edmonton, AB, Canada, 2006.
- Canadian Climate Normals 1981–2010 Station Data. Available online: https://climate.weather.gc.ca/climate_normals/results_1981_2010_e.html?searchType=stnProv&lstProvince=AB&txtCentralLatMin=0&txtCentralLatSec=0&txtCentralLongMin=0&txtCentralLongSec=0&stnID=2704&dispBack=0 (accessed on 22 April 2020).
- Pinno, B.D.; Errington, R.C. Maximizing natural trembling aspen seedling establishment on a reclaimed boreal oil sands site. Ecol. Restor. 2015, 33, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Pinno, B.D.; Hawkes, V.C. Temporal trends of ecosystem development on different site types in reclaimed boreal forests. Forests 2015, 6, 2109–2124. [Google Scholar] [CrossRef] [Green Version]
- Tannas, K.E. Common Plants of the Western Rangelands—Volume 3: Forbs, 1st ed.; Alberta Agriculture, Food and Rural Development: Lethbridge, AB, Canada, 2004. [Google Scholar]
- Tannas, K.E. Common Plants of the Western Rangelands—Volume 1: Grasses and Grass-Like Species, 1st ed.; Alberta Agriculture, Food and Rural Development: Lethbridge, AB, Canada, 2001. [Google Scholar]
- Moss, E.H.; Packer, J. Flora of Alberta, 2nd ed.; University of Toronto Press: Toronto, ON, Canada, 1994. [Google Scholar]
- Tannas, K.E. Common Plants of the Western Rangelands—Volume 2: Trees and Shrubs, 1st ed.; Alberta Agriculture, Food and Rural Development: Lethbridge, AB, Canada, 2003. [Google Scholar]
- United States Department of Agriculture. Available online: https://plants.sc.egov.usda.gov/adv_search.html (accessed on 7 December 2019).
- de Bortoli, L.A.; Pinno, B.D.; Mackenzie, M.D.; Li, E.H. Plant community composition and tree seedling establishment in response to seeding and weeding treatments on different reclamation cover soils. Can. J. For. Res. 2019, 49, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, S.E.; Snively, A.E.K.; Fair, J.M.; Landhausser, S.M. Early trajectories of forest understory development on reclamation sites: Influence of forest floor placement and a cover crop. Restor. Ecol. 2015, 23, 698–706. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Macdonald, S.E.; Hogg, E.H. Ecology of and control strategies for Calamagrostis canadensis in boreal forest sites. Can. J. For. Res. 1993, 23, 2070–2077. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Lieffers, V.J. Rhizome plasticity and clonal foraging of Calamagrostis canadensis in response to habitat heterogeneity. J. Ecol. 1993, 81, 769–776. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Mulak, T.L.; Lieffers, V.J. The effect of roots and litter of Calamagrostis canadensis on root sucker regeneration of Populus tremuloides. Forestry 2007, 80, 481–488. [Google Scholar] [CrossRef]
- Landhausser, S.M.; Stadt, K.J.; Lieffers, V.J. Screening for control of a forest weed: Early competition between three replacement species and Calamagrostis canadensis of Picea glauca. J. Appl. Ecol. 1996, 33, 1517. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Fenniak, T.E. Understory plant communities of boreal mixedwood forests in western Canada: Natural patterns and response to variable-retention harvesting. For. Ecol. Manag. 2007, 242, 34–48. [Google Scholar] [CrossRef]
- Pinno, B.D.; Sherr, I.; Errington, R.C.; Shea, K. Islands – Soil Patches and Plant Community Dynamics on a New Oil Sands Reclamation Design. J. Am. Soc. Min. Reclam. 2016, 5, 28–44. [Google Scholar] [CrossRef]
- Package ‘lmperm’. Available online: https://cran.r-project.org/web/packages/lmPerm/lmPerm.pdf (accessed on 7 December 2019).
- Package ‘mvpart’. Available online: https://mran.microsoft.com/snapshot/2014-12-11/web/packages/mvpart/mvpart.pdf (accessed on 7 December 2019).
- Foster, B.L.; Tilman, D. Dynamic and static views of succession: Testing the descriptive power of the chronosequence approach. Plant Ecol. 2000, 146, 1–10. [Google Scholar] [CrossRef]
- Alday, J.G.; Pallavicini, Y.; Marrs, R.H.; Martínez-Ruiz, C. Functional groups and dispersal strategies as guides for predicting vegetation dynamics on reclaimed mines. Plant Ecol. 2011, 212, 1759–1775. [Google Scholar] [CrossRef]
- Spellman, K.V.; Schneller, L.C.; Mulder, C.P.H.; Carlson, M.L. Effects of non-native Melilotus albus on pollination and reproduction in two boreal shrubs. Oecologia 2015, 179, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Wedin, D. Dynamics of nitrogen competition between successional grasses. Ecology 1991, 72, 1038–1049. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Dhar, A.; Comeau, P.G.; Karst, J.; Pinno, B.D.; Chang, S.X.; Naeth, A.M.; Vassov, R.; Bampfylde, C. Plant community development following reclamation of oil sands mine sites in the boreal forest: A review. Environ. Rev. 2018, 298, 1–13. [Google Scholar] [CrossRef]
- Plants Database: Sonchus arvensis, L. Field Sowthistle. Available online: https://plants.usda.gov/core/profile?symbol=SOAR2 (accessed on 7 December 2019).
- Turkington, R.A.; Cavers, P.B.; Rempel, E. The biology of Canadian weeds. 29. Melilotus alba Desr. and M. offtcinalis (L.) Lam. ROY. Can. J. Planr Sci. 1978, 58, 523–537. [Google Scholar] [CrossRef]
- Provincially Regulated Weeds. Available online: https://www.alberta.ca/provincially-regulated-weeds.aspx (accessed on 7 December 2019).
- Trepanier, K.E.; Pinno, B.D.; Errington, R.C. Dominant drivers of plant community assembly vary by soil type and time in reclaimed forests. Plant Ecol. 2020. [Google Scholar] [CrossRef]
- Brown, C.S.; Bugg, R.L. Effects of established perennial grasses on introduction of native forbs in California. Restor. Ecol. 2001, 9, 38–48. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Lieffers, V.J. Growth of Populus tremuloides in association with Calamagrostis canadensis. Can. J. For. Res. 1998, 28, 396–401. [Google Scholar] [CrossRef]
- Henkel-Johnson, D.; Macdonald, S.E.; Bork, E.W.; Thomas, B.R. Influence of weed composition, abundance, and spatial proximity on growth in young hybrid poplar plantations. For. Ecol. Manag. 2016, 362, 55–68. [Google Scholar] [CrossRef]
- Strong, W.L. Vegetation development on reclaimed lands in the Coal Valley Mine of western Alberta, Canada. Can. J. Bot. 2000, 78, 110–118. [Google Scholar] [CrossRef]
- Audet, P.; Pinno, B.D.; Thiffault, E. Reclamation of boreal forest after oil sands mining: Anticipating novel challenges in novel environments. Can. J. For. Res. 2015, 45, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.A.; Chen, H.Y.H. Understory vegetation dynamics of North American boreal forests. CRC. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Cole, R.J.; Holl, K.D.; Zahawi, R.A. Seed rain under tree islands planted to restore degraded lands in a tropical agricultural landscape. Ecol. Appl. 2010, 20, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 707–720. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| FFMM | PMM | p | ||
|---|---|---|---|---|
| Soil | Soil Temperature (°C) | 14.1 (2.2) | 15.4 (1.4) | 0.090 |
| VWC (%) | 16.5 (4.4) | 27.0 (6.4) | <0.001 | |
| pH | 6.9 (0.7) | 6.5 (0.9) | 0.176 | |
| Average Species Richness Per 1 m2 Quadrat | Tree | 1 | 2 | <0.001 |
| Native Forb | 10 | 9 | 0.118 | |
| Non-Native Forb | 3 | 4 | 0.270 | |
| Graminoid | 5 | 5 | 1 | |
| Shrub | 2 | 1 | 0.980 |
| FFMM | PMM | |||
|---|---|---|---|---|
| Functional Group | Species | Average Cover Per Plot (%) | Species | Average Cover Per Plot (%) |
| Non-Native Forbs | Sonchus arvensis | 2.6 | Melilotus alba | 4.4 |
| Melilotus alba | <1.0 | Sonchus arvensis | 3.7 | |
| Taraxacum officinale | <1.0 | Crepis tectorum | 1.5 | |
| Crepis tectorum | <1.0 | Taraxacum officinale | <1.0 | |
| Native Forbs | Chamerion angustifolium | 6.2 | Chamerion angustifolium | 6.5 |
| Equisetum arvense | 3.9 | Equisetum arvense | 5.8 | |
| Lathyrus ochroleucus | 1.9 | Achillea millefolium | 1.2 | |
| Fragaria virginiana | 1.7 | Lathyrus ochroleucus | <1.0 | |
| Graminoids | Calamagrostis canadensis | 6.8 | Calamagrostis canadensis | 5.3 |
| Poa pratensis | 2.7 | Agropyron trachycaulum | 1.0 | |
| Agropyron trachycaulum | 2.5 | Carex spp. | <1.0 | |
| Leymus innovatus | 1.3 | Poa pratensis | <1.0 | |
| Shrubs | Rubus idaeus | 3.7 | Salix spp. | 1.0 |
| Rosa acicularis | <1.0 | Rubus idaeus | <1.0 | |
| Cornus sericea | <1.0 | Rosa acicularis | <1.0 | |
| Ribes oxyacanthoides | <1.0 | Shepherdia canadensis | <1.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trepanier, K.E.; Burton, B.; Pinno, B.D. Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site? Diversity 2021, 13, 76. https://doi.org/10.3390/d13020076
Trepanier KE, Burton B, Pinno BD. Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site? Diversity. 2021; 13(2):76. https://doi.org/10.3390/d13020076
Chicago/Turabian StyleTrepanier, Kaitlyn E., Brea Burton, and Bradley D. Pinno. 2021. "Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site?" Diversity 13, no. 2: 76. https://doi.org/10.3390/d13020076
APA StyleTrepanier, K. E., Burton, B., & Pinno, B. D. (2021). Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site? Diversity, 13(2), 76. https://doi.org/10.3390/d13020076