
Annelids in Extreme Aquatic Environments: Diversity, Adaptations and Evolution



Abstract
:1. Introduction
2. What Is a Polychaete?
3. What Is a Clitellate?
4. What Is an Extreme Environment? What Is an “Extremophile”?
5. Terrestrial Freshwater
6. Hypersalinity and Aridity
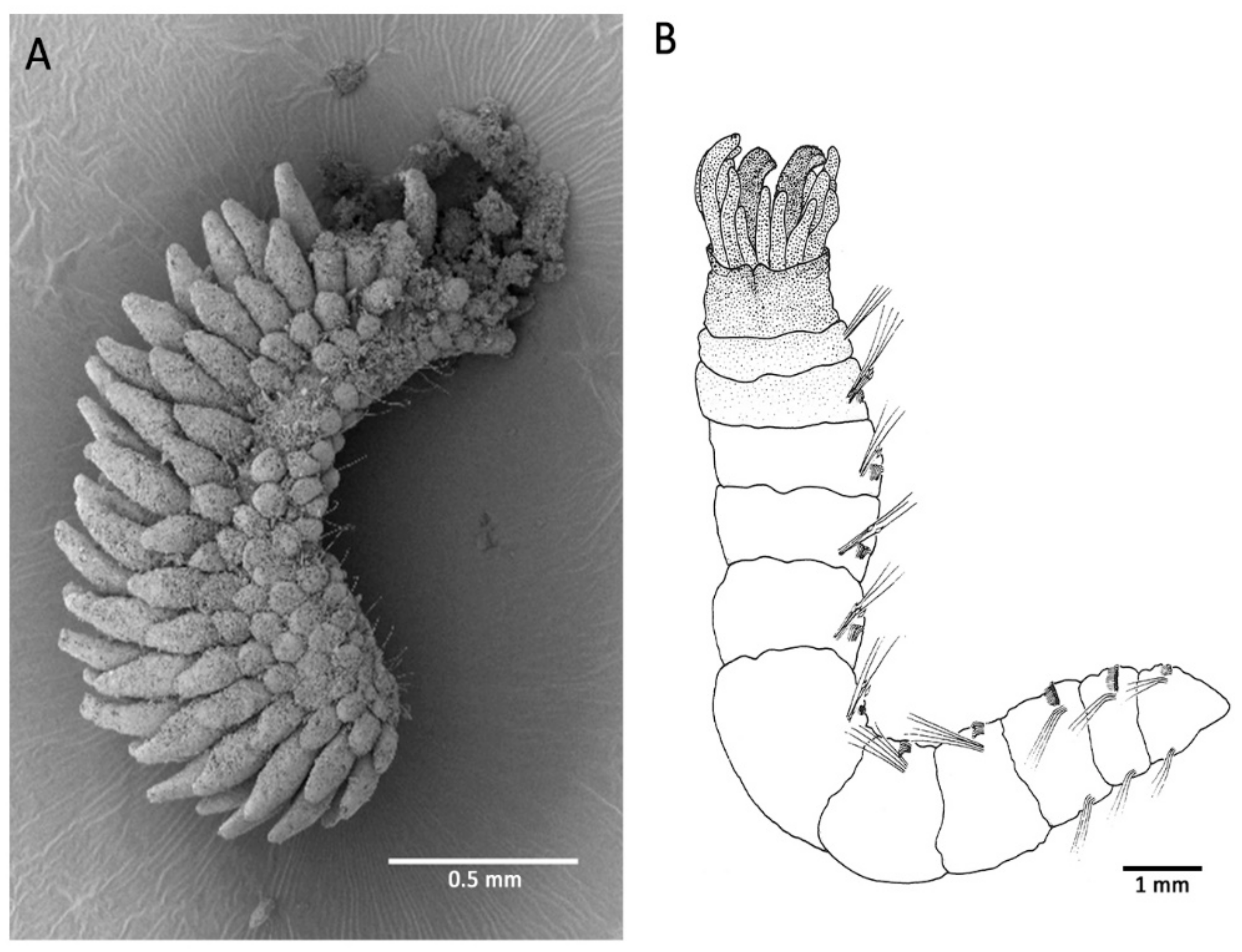
7. Extreme Temperatures, and Vents and Seeps
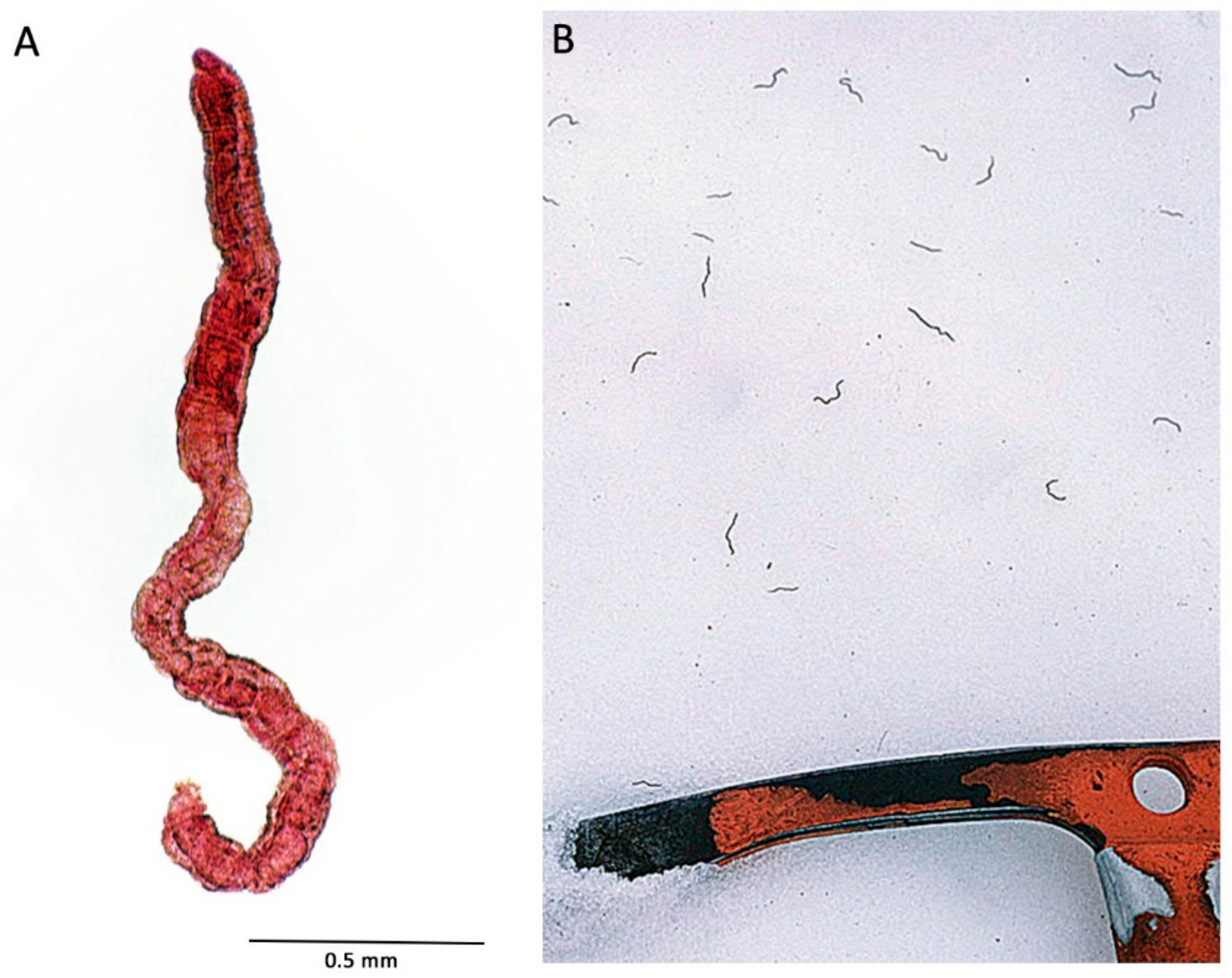
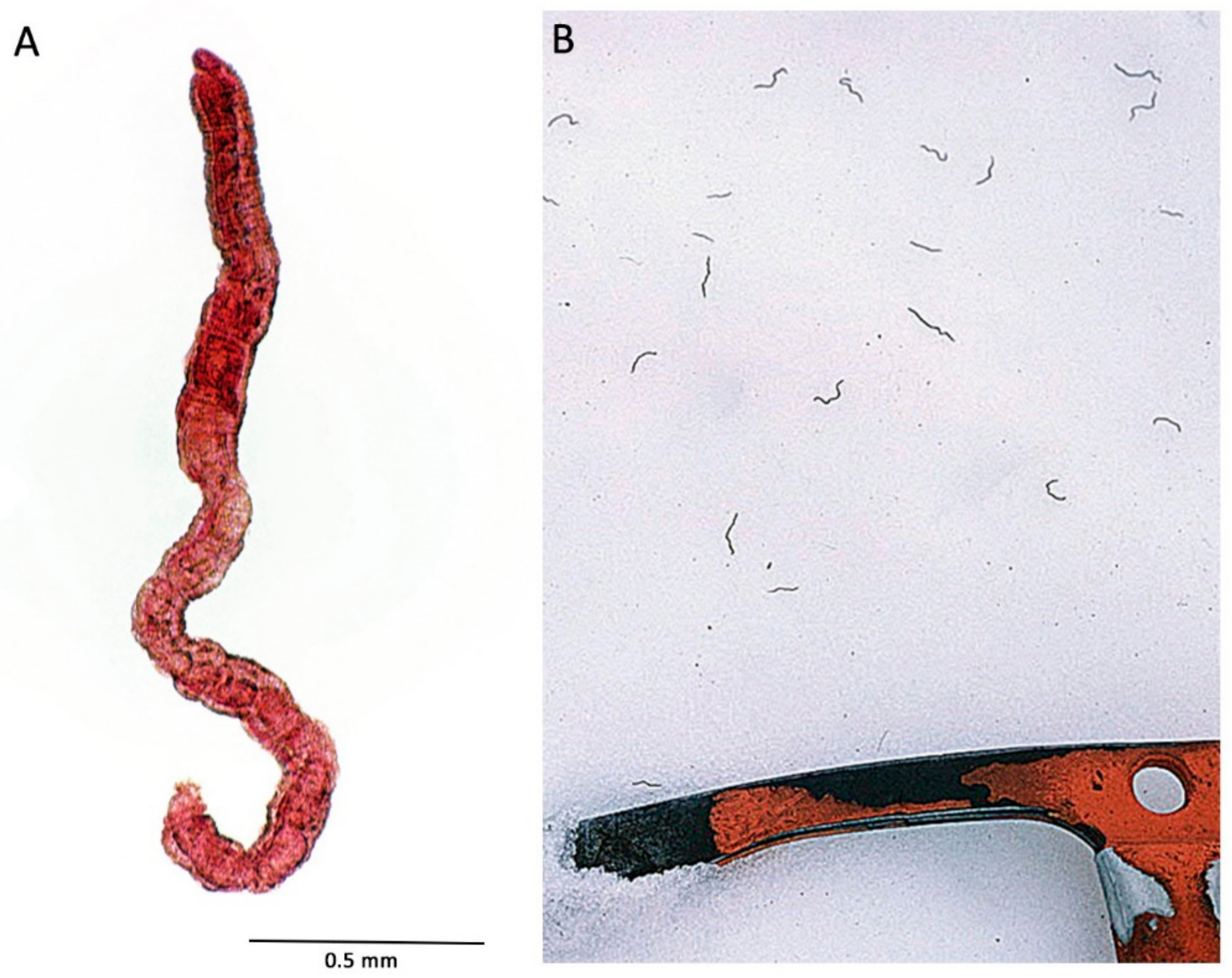
8. Extreme Depths, and Other Depth-Related Factors
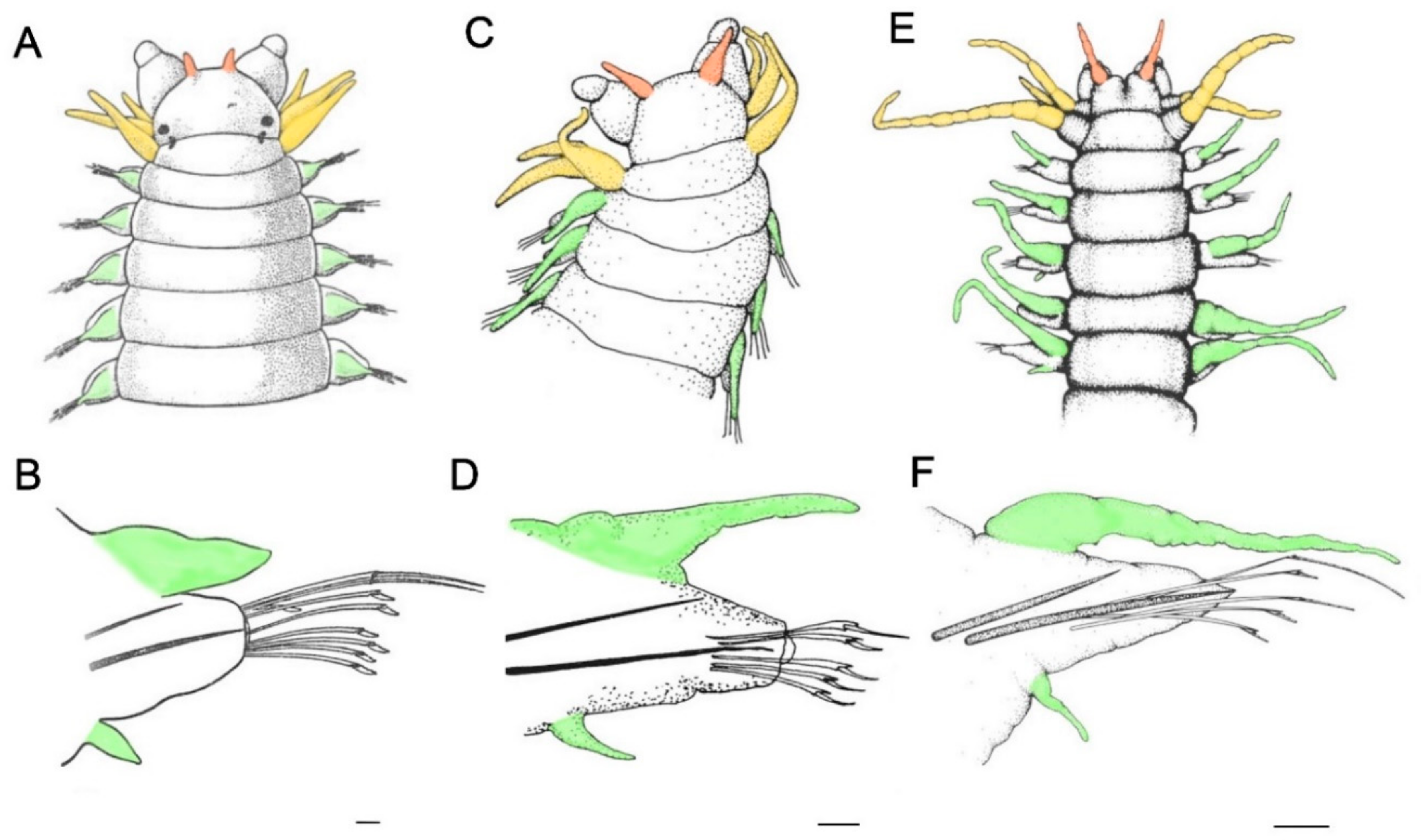
9. Subterranean and Cave Environments
10. High-Energy Habitats: Mobile Sediments and Surf Zone
11. Low Oxygen: Organic Enrichment, Sulfidic, Methanic Sediments
12. Inorganic and Organic Contaminated Sediments
13. Extreme pH
14. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Struck, T.H.; Purschke, G.; Dordel, J.; Hösel, C.; Nesnidal, M.P.; Diersing, F.; Bleidorn, C.; Paul, C.; Hill, N.; Tiedemann, R.; et al. Phylogeny and evolution of Annelida based on molecular data. In Deep Metazoan Phylogeny: The Backbone of the Tree of Life. New Insights from Analyses of Molecules, Morphology, and Theory of Data Analysis; Wägele, J.W., Bartolomaeus, T., Eds.; De Gruyter: Berlin, Germany, 2014; pp. 143–160. [Google Scholar] [CrossRef] [Green Version]
- Weigert, A.; Bleidorn, C. Current status of annelid phylogeny. Org. Divers. Evol. 2016, 16, 345–362. [Google Scholar] [CrossRef]
- Struck, T.H.; Paul, C.; Hill, N.; Hartmann, S.; Hosel, C.; Kube, M.; Lieb, B.; Meyer, A.; Tiedemann, R.; Purschke, G.; et al. Phylogenomic analyses unravel annelid evolution. Nature 2011, 471, 95–98. Available online: http://www.nature.com/nature/journal/v471/n7336/abs/10.1038-nature09864-unlocked.html#supplementary-information (accessed on 20 December 2020). [CrossRef]
- Kuo, D.H. The polychaete-to-clitellate transition: An EvoDevo perspective. Dev. Biol. 2017, 427, 230–240. [Google Scholar] [CrossRef]
- Chapman, A.D. Numbers of Living Species in Australia and the World, 2nd ed.; Australian Biological Resources Study: Canberra, Austrilia, 2009; p. 84. [Google Scholar]
- Struck, T.H. 2. Phylogeny. In Handbook of Zoology. Annelida. Basal Groups and Pleistoannelida, Sedentaria I; Purschke, G., Böggemann, M., Wilfried, W., Eds.; De Gruyter: Berlin, Germany, 2019; pp. 37–68. [Google Scholar] [CrossRef]
- Purschke, G. On the ground pattern of Annelida. Org. Divers. Evol. 2002, 2, 181–196. [Google Scholar] [CrossRef] [Green Version]
- Purschke, G.; Bleidorn, C.; Struck, T. Systematics, evolution and phylogeny of Annelida—a morphological perspective. Mem. Mus. Vic. 2014, 71, 247–269. [Google Scholar] [CrossRef] [Green Version]
- Struck, T.H. Direction of evolution within Annelida and the definition of Pleistoannelida. J. Zool. Syst. Evol. Res. 2011, 49, 340–345. [Google Scholar] [CrossRef]
- Rouse, G.W.; Pleijel, F. Polychaetes; Oxford University Press: Oxford, UK, 2001; p. xiii + 354. [Google Scholar]
- Erséus, C.; Williams, B.W.; Horn, K.M.; Halanych, K.M.; Santos, S.R.; James, S.W.; Châtelliers, M.C.; Anderson, F.E. Phylogenomic analyses reveal a Paleozoic radiation and support a freshwater origin for clitellate annelids. Zool. Scr. 2020, 49, 614–640, in press. [Google Scholar] [CrossRef]
- Erséus, C. Phylogeny of oligochaetous Clitellata. Hydrobiologia 2005, 535–536, 357–372. [Google Scholar]
- Ferraguti, M.; Erséus, C. Sperm types and their use for a phylogenetic analysis of aquatic clitellates. Hydrobiologia 1999, 402, 225–237. [Google Scholar] [CrossRef]
- Martin, P.; Martínez-Ansemil, E.; Pinder, A.; Timm, T.; Wetzel, M.J. Global diversity of oligochaetous clitellates (“Oligochaeta”; Clitellata) in freshwater. Hydrobiologia 2008, 595, 117–127. [Google Scholar] [CrossRef]
- Purschke, G.; Westheide, W.; Rhode, D.; Brinkhurst, R.O. Morphological reinvestigation and phylogenetic relationship of Acanthobdella peledina (Annelida, Clitellata). Zoomorphology 1993, 113, 91–101. [Google Scholar] [CrossRef]
- Rouse, G.W.; Fauchald, K. The articulation of annelids. Zool. Scr. 1995, 24, 269–301. [Google Scholar] [CrossRef]
- Timm, T.; Martin, P.J. Chapter 21—Clitellata: Oligochaeta. In Thorp and Covich’s Freshwater Invertebrates, 4th ed.; Thorp, J.H., Rogers, D.C., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 529–549. [Google Scholar] [CrossRef]
- Westheide, W. The direction of evolution within the Polychaeta. J. Nat. Hist. 1997, 31, 1–15. [Google Scholar] [CrossRef]
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef]
- Gómez, F. Extreme Environment. In Encyclopedia of Astrobiology; Gargaud, M., Amils, R., Quintanilla, J.C., Cleaves, H.J., Irvine, W.M., Pinti, D.L., Viso, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 570–572. [Google Scholar] [CrossRef]
- Macelroy, R.D. Some comments on the evolution of extremophiles. Biosystems 1974, 6, 74–75. [Google Scholar] [CrossRef]
- Glasby, C.J.; Timm, T.; Muir, A.I.; Gil, J. Catalogue of non-marine Polychaeta (Annelida) of the world. Zootaxa 2009, 2070, 1–52. [Google Scholar] [CrossRef]
- Solís-Weiss, V.; Espinasa, L. Lycastilla cavernicola, a new freshwater nereidid from an inland Mexican cave (Polychaeta: Nereididae: Namanereidinae). Proc. Biol. Soc. Wash. 1991, 104, 631–639. [Google Scholar]
- Glasby, C.J.; Timm, T. Global diversity of polychaetes (Polychaeta; Annelida) in freshwater. In Freshwater Animal Diversity, Assessment; Balian, E.V., Lévêque, C., Segers, H., Martens, K., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 107–115. [Google Scholar] [CrossRef]
- Glasby, C.J.; Fiege, D.; Van Damme, K. Stygobiont polychaetes: Notes on the morphology and the origins of groundwater Namanereis (Annelida: Nereididae: Namanereidinae), with a description of two new species. Zool. J. Linn. Soc. 2014, 171, 22–37. [Google Scholar] [CrossRef]
- Glasby, C.J. Population structure and reproductive biology of Ceratonereis limnetica (Polychaeta: Nereididae) at Lower Portland, Hawkesbury River, Australia. Mar. Biol. 1986, 90, 589–595. [Google Scholar] [CrossRef]
- Pudovkina, T.A.; Sitnikova, T.Y.; Matveyev, A.N.; Shcherbakov, D.Y. The history of Manayunkia [Polychaeta: Sedentaria: Sabellidae] propagation in North Eastern Asia. Ecol. Genet. 2014, 12, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Sitnikova, T.Y. Chapter 14: Polychaete worms (Annelida: Polychaeta) [In Russian]. In Index of Animal Species Inhabiting Lake Baikal and Its Catchment Area. Volume 1: Lake Baikal; Timoshkin, O.A., Proviz, V.I., Sitnikova, T.Y., Eds.; Russian Academy of Sciences: Novosibirsk, Russia, 2004; Volume 1, pp. 428–431. [Google Scholar]
- Zerbst-Boroffka, I. Osmotic Adaptation of the Endemic Fauna to the Ancient Freshwater Lake Baikal. Naturwissenschaften 1999, 86, 330–333. [Google Scholar] [CrossRef]
- Hutchings, P.A. Part C. Natural History. In Polychaetes & Allies. The Southern Synthesis. Fauna of Australia Vol 4A Polychaeta, Myzostomida, Pogonophora, Echiura, Sipuncula; Beesley, P.L., Ross, G.J.B., Glasby, C.J., Eds.; CSIRO Publishing: Melbourne, Australia, 2000; pp. 30–37. [Google Scholar]
- Hutchings, P.A.; De Deckker, P.; Geddes, M.C. A new species of Manayunkia (Polychaeta) from ephemeral lakes near the Coorong, South Australia. Trans. R. Soc. S. Aust. 1981, 105, 25–28. [Google Scholar]
- Van Gaest, A.L.; Young, C.M.; Young, J.J.; Helms, A.R.; Arellano, S.M. Physiological and behavioral responses of Bathynerita naticoidea (Gastropoda: Neritidae) and Methanoaricia dendrobranchiata (Polychaeta: Orbiniidae) to hypersaline conditions at a brine pool cold seep. Mar. Ecol. 2007, 28, 199–207. [Google Scholar] [CrossRef]
- Erséus, C. Marine Tubificidae (Oligochaeta) of the Arabian Gulf Coast of Saudi Arabia (part 4). Fauna Saudi Arab. 1988, 9, 19–22. [Google Scholar]
- Erséus, C. Marine Tubificidae (Oligochaeta) from a mangrove habitat in Kenya. Trop. Zool. 1999, 12, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Healy, B. Marionina schrijversi (Oligochaeta Enchytraeidae), a new species from a mangrove in Kenya. Trop. Zool. 1997, 10, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.; Aït Boughrous, A. Guide Taxonomique des Oligochètes Aquatiques du Maghreb; Belgian National Focal Point to the Global Taxonomy Initiative; Royal Belgian Institute of Natural Sciences: Brussels, Belgium, 2012; Volume 12, p. 186. [Google Scholar]
- Boumaiza, M.; Martínez-Ansemil, E.; Giani, N. Les Oligochètes et Aphanoneura des eaux courantes de Tunisie. I—Données faunistiques. Ann. Limnol. 1986, 22, 231–237. [Google Scholar] [CrossRef]
- Schmelz, R.M.; Collado, R. A guide to European terrestrial and freshwater species of Enchytraeidae (Oligochaeta). Soil Org. 2010, 82, 1–176. [Google Scholar]
- Erséus, C. A generic revision of the Phallodrilinae (Oligochaeta, Tubificidae). Zool. Scr. 1992, 21, 5–48. [Google Scholar] [CrossRef]
- Brinkhurst, R.O.; Jamieson, B.G.M.G. Aquatic Oligochaeta of the World; Oliver and Boyd: Edinburgh, Scotland, 1971; p. 860. [Google Scholar]
- Timm, T. A guide to the freshwater Oligochaeta and Polychaeta of Northern and Central Europe. Lauterbornia 2009, 66, 1–235. [Google Scholar]
- Jans, W.; Römbke, J. Funde eines terrestrischen Polychaeten (Annelida) in Wäldern Baden-Württembergs. Carolinea 1989, 47, 158–162. [Google Scholar]
- Dózsa-Farkas, K.; Hong, Y. Three new Hemienchytraeus species (Enchytraeidae, Oligochaeta, Annelida) from Korea, with first records of other enchytraeids and terrestrial polychaetes (Annelida). Zootaxa 2010, 2406, 29–56. [Google Scholar] [CrossRef]
- Schmidt, P. Zonation of the interstitial polychaete Stygocapitella subterranea (Stygocapitellidae) in European sandy beaches. Mar. Biol. 1970, 7, 319–323. [Google Scholar] [CrossRef]
- Purschke, G. Terrestrial polychaetes—models for the evolution of the Clitellata (Annelida) ? Hydrobiologia 1999, 406, 87–99. [Google Scholar] [CrossRef]
- Westheide, W. Interstitial Families; The Linnean Society of London: London, UK, 2008; p. 169. [Google Scholar]
- Rota, E.; de Jong, Y. Fauna Europaea: Annelida—Terrestrial Oligochaeta (Enchytraeidae and Megadrili), Aphanoneura and Polychaeta. Biodivers. Data J. 2015, 3, e5737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struck, T.H.; Koczula, J.; Stateczny, D.; Meyer, C.; Purschke, G. Two new species in the annelid genus Stygocapitella (Orbiniida, Parergodrilidae) with comments on their biogeography. Zootaxa 2017, 4286, 301–332. [Google Scholar] [CrossRef]
- Meca, M.A.; Zhadan, A.; Struck, T.H. The early branching group of Orbiniida sensu Struck et al., 2015: Parergodrilidae and Orbiniidae. Diversity 2021, 13, 29. [Google Scholar] [CrossRef]
- Rota, E.; Lupetti, P. An ultrastructural investigation of Hrabeiella Pizl & Chalupsky, 1984 (Annelida). I. Chaetae and body wall organization. Hydrobiologia 1996, 334, 229–239. [Google Scholar]
- Stout, J.D. Aquatic oligochaetes occurring in forest litter.-I. Trans. R. Soc. N. Z. 1956, 84, 97–102. [Google Scholar]
- Herlant-Meewis, H. Cyst-formation in Aeolosoma hemprichi (Ehr). Biol. Bull. 1950, 99, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Westheide, W.; von Basse, M. Chilling and freezing resistance of two interstitial polychaetes from a sandy tidal beach. Oecologia 1978, 33, 45–54. [Google Scholar] [CrossRef]
- Holmstrup, M. Cold hardiness strategy in cocoons of the lumbricid earthworm Dendrobaena octaedra (Savigny). Comp. Biochem. Physiol. Part A Physiol. 1992, 102, 49–54. [Google Scholar] [CrossRef]
- Zachariassen, K.E. Physiology of cold tolerance in insects. Physiol. Rev. 1985, 65, 799–832. [Google Scholar] [CrossRef] [PubMed]
- Sømme, L.; Birkemoe, T. Cold tolerance and dehydration in Enchytraeidae from Svalbard. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 1997, 167, 264–269. [Google Scholar] [CrossRef]
- Slotsbo, S.; Maraldo, K.; Malmendal, A.; Nielsen, N.C.; Holmstrup, M. Freeze tolerance and accumulation of cryoprotectants in the enchytraeid Enchytraeus albidus (Oligochaeta) from Greenland and Europe. Cryobiology 2008, 57, 286–291. [Google Scholar] [CrossRef]
- Goodman, D. Ecological Investigations of Ice Worms on Casement Glacier, Southeastern Alaska; Ohio State University Research Foundation: Columbus, OH, USA, 1971. [Google Scholar]
- Shain, D.H.; Mason, T.A.; Farrell, A.H.; Michalewicz, L.A. Distribution and behavior of ice worms (Mesenchytraeus solifugus) in South-central Alaska. Can. J. Zool. 2001, 79, 1813–1821. [Google Scholar] [CrossRef]
- Edwards, J.S. How small ectotherms thrive in the cold without really trying. Cryo Lett. 1986, 6, 388–390. [Google Scholar]
- Napolitano, M.J.; Nagele, R.G.; Shain, D.H. The ice worm, Mesenchytraeus solifugus, elevates adenylate levels at low physiological temperature. Comp. Biochem. Physiol. 2004, 137, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Miles, H.B. Heat-death temperature in Allolobophora terrestris (Sav.) forma longa (Ude) and Eisenia foetida (Sav.). Nature 1963, 199, 826. [Google Scholar] [CrossRef]
- Johnston, M.R.; Herrick, B.M. Cocoon Heat Tolerance of Pheretimoid Earthworms Amynthas tokioensis and Amynthas agrestis. Am. Midl. Nat. 2019, 181, 299–309. [Google Scholar] [CrossRef]
- Girguis, P.R.; Lee, R.W. Thermal preference and tolerance of alvinellids. Science 2006, 312, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaill, F.; Bouligand, Y. Supercoil of collagen fibrils in the integument of Alvinella, an abyssal annelid. Tissue Cell 1987, 19, 625–642. [Google Scholar] [CrossRef]
- Gaill, F.; Hunt, S. Tubes of deep sea hydrothermal vent worms Riftia pachyptila (Vestimentifera) and Alvinella pompejana (Annelida). Mar. Ecol. Prog. Ser. 1986, 34, 267–274. [Google Scholar] [CrossRef]
- Jollivet, D.; Desbruyères, D.; Ladrat, C.; Laubier, L. Evidence for differences in the allozyme thermostability of deep-sea hydrothermal vent polychaetes (Alvinellidae):a possible selection by habitat. Mar. Ecol. Prog. Ser. 1995, 123, 125–136. [Google Scholar] [CrossRef]
- Sicot, F.X.; Mesnage, M.; Masselot, M.; Exposito, J.Y.; Garrone, R.; Deutsch, J.; Gaill, F. Molecular adaptation to an extreme environment: Origin of the thermal stability of the Pompeii worm collagen. J. Mol. Biol. 2000, 302, 811–820. [Google Scholar] [CrossRef]
- Rinke, C.; Lee, R.W. Pathways, activities and thermal stability of anaerobic and aerobic enzymes in thermophilic vent paralvinellid worms. Mar. Ecol. Prog. Series 2009, 382, 99–112. [Google Scholar] [CrossRef]
- Dilly, G.F.; Young, C.R.; Lane, W.S.; Pangilinan, J.; Girguis, P.R. Exploring the limit of metazoan thermal tolerance via comparative proteomics: Thermally induced changes in protein abundance by two hydrothermal vent polychaetes. Proc. Biol. Sci. 2012, 279, 3347–3356. [Google Scholar] [CrossRef] [Green Version]
- McHugh, D.; Tunnicliffe, V. Ecology and reproductive biology of the hydrothermal vent polychaete Amphisamytha galapagensis (Ampharetidae). Mar. Ecol. Prog. Ser. 1994, 106, 111–120. [Google Scholar] [CrossRef]
- Erséus, C.M.; Milligan, M.R. Three new species of Tubificidae (Oligochaeta) from an oil seepage area on the continental slope of the northern Gulf of Mexico. Proc. Biol. Soc. Wash. 1989, 102, 887–893. [Google Scholar]
- Kvist, S.; Dreyer, J.; Erséus, C. Two new species of Tubificoides (Annelida: Clitellata: Naididae) from the Blake Ridge methane seep in the northwest Atlantic Ocean. Proc. Biol. Soc. Wash. 2008, 121, 531–540. [Google Scholar] [CrossRef]
- Petrecca, R.F.; Grassle, J.F. Notes on Fauna from Several Deep-Sea Hydrothermal Vent and Cold Seep Soft-Sediment Communities; Springer: New York, NY, USA, 1990; pp. 279–283. [Google Scholar]
- Fisher, C.R.; Childress, J.J.; Arp, A.J.; Brooks, J.M.; Distel, D.; Favuzzi, J.A.; Macko, S.A.; Newton, A.; Powell, M.A.; Somero, G.N.; et al. Physiology, morphology, and biochemical composition of Riftia pachyptila at Rose Garden in 1985. Deep Sea Res. Part A Oceanogr. Res. Pap. 1988, 35, 1745–1758. [Google Scholar] [CrossRef]
- Hessler, R.R.; Smithey, W.M.; Boudrias, M.A.; Keller, C.H.; Lutz, R.A.; Childress, J.J. Temporal change in megafauna at the Rose Garden hydrothermal vent (Galapagos Rift; eastern tropical Pacific). Deep Sea Res. Part A Oceanogr. Res. Pap. 1988, 35, 1681–1709. [Google Scholar] [CrossRef]
- Southward, E.C.; Tunnicliffe, V.; Black, M.B.; Dixon, D.R.; Dixon, L.R.J. Ocean-ridge segmentation and vent tubeworms (Vestimentifera) in the NE Pacific. Geol. Soc. Lond. Spec. Publ. 1996, 118, 211–224. [Google Scholar] [CrossRef]
- Fisher, C.R.; Urcuyo, I.A.; Simpkins, M.A.; Nix, E. Life in the slow lane: Growth and longevity of cold-seep vestimentiferans. Mar. Ecol. 1997, 18, 83–94. [Google Scholar] [CrossRef]
- Bergquist, D.C.; Williams, F.M.; Fisher, C.R. Longevity record for deep-sea invertebrate. Nature 2000, 403, 499–500. [Google Scholar] [CrossRef] [PubMed]
- Cordes, E.E.; Bergquist, D.C.; Redding, M.L.; Fisher, C.R. Patterns of growth in cold-seep vestimenferans including Seepiophila jonesi: A second species of long-lived tubeworm. Mar. Ecol. 2007, 28, 160–168. [Google Scholar] [CrossRef]
- Crane, K.; Hecker, B.; Golubev, V. Hydrothermal vents in Lake Baikal. Nature 1991, 350, 281. [Google Scholar] [CrossRef]
- Grachev, M.; Flalkov, V.; Nakamura, T.; Ohta, T.; Kawai, T. Extant fauna of ancient carbon. Nature 1995, 374, 123–124. [Google Scholar] [CrossRef]
- Sitnikova, T.Y.; Shirokaya, A.A. New data on deep water Baikal limpets found in hydrothermal vents and oil-seeps. Arch. Molluskenkd. Int. J. Malacol. 2013, 142, 257–278. [Google Scholar] [CrossRef]
- Kaygorodova Irina, A. Clitellate communities in anomalous region of Lake Baikal, Eastern Siberia (Russia). Biologia 2016, 71, 31–37. [Google Scholar] [CrossRef]
- Martin, P.; Martens, K.; Goddeeris, B. Oligochaeta from the abyssal zone of Lake Baikal (Siberia, Russia). Hydrobiologia 1999, 406, 165–174. [Google Scholar] [CrossRef]
- Weiss, R.F.; Carmack, E.C.; Koropalov, V.M. Deep-water renewal and biological production in Lake Baikal. Nature 1991, 349, 665–669. [Google Scholar] [CrossRef]
- Erséus, C.; Rota, E. New findings and overview of the oligochaetous Clitellata (Annelida) of the North Atlantic deep sea. Proc. Biol. Soc. Wash. 2003, 116, 892–900. [Google Scholar]
- Erséus, C. Taxonomic revision of the marine genera Bathydrilus Cook and Macroseta Erséus (Oligochaeta, Tubificidae), with descriptions of six new species and subspecies. Zool. Scr. 1979, 8, 139–151. [Google Scholar] [CrossRef]
- Rota, E.; Erséus, C. New records of Grania (Clitellata, Enchytraeidae) in the Northeast Atlantic (from Tromso to the Canary Islands), with descriptions of seven new species. Sarsia 2003, 88, 210–243. [Google Scholar] [CrossRef]
- Erséus, C. Taxonomic revision of the Marine Genus Phallodrilus Pierantoni (Oligochaeta, Tubificidae), with description of thirteen new species. Zool. Scr. 1979, 8, 187–208. [Google Scholar] [CrossRef]
- Kozhova, O.M.; Izmest′eva, L.R. Lake Baikal. Evolution and Biodiversity; Backhuys Publishers: Leiden, The Netherlands, 1998; p. 447. [Google Scholar]
- Martin, P.; Granina, L.; Martens, K.; Goddeeris, B. Oxygen concentration profiles in sediments of two ancient lakes: Lake Baikal (Sideria, Russia) and Lake Malawi (East Africa). Hydrobiologia 1998, 367, 163–174. [Google Scholar] [CrossRef]
- Sherstyankin, P.P.; Alekseev, S.P.; Abramov, A.M.; Stavrov, K.G.; De Batist, M.; Hus, R.; Canals, M.; Casamor, J.L. Computer-based bathymetric map of Lake Baikal. Dokl. Earth Sci. 2006, 408, 564–569. [Google Scholar] [CrossRef]
- Pudovkina, T.A.; Sitnikova, T.Y.; Matveyev, A.N.; Shcherbakov, D.Y. Kindred Relations of Baikal polychaete of the Manayunkia Genus [Polychaeta: Sedentaria: Sabellidae] according to CO1 and settlement history analysis. Russ. J. Genet. Appl. Res. 2016, 6, 129–137. [Google Scholar] [CrossRef]
- Falkner, K.K.; Measures, C.I.; Herbelin, S.E.; Edmond, J.M.; Weiss, R.F. The major and minor element geochemistry of Lake Baikal. Limnol. Oceanogr. 1991, 36, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.; Sonet, G.; Smitz, N.; Backeljau, T. Phylogenetic analysis of the Baikalodrilus species flock (Annelida: Clitellata: Naididae), an endemic genus to Lake Baikal (Russia). Zool. J. Linn. Soc. 2019, 187, 987–1015. [Google Scholar] [CrossRef]
- Culver, D.C.; White, W.B. Encyclopedia of Caves, 2nd ed.; Academic Press: Waltham, MA, USA, 2011; p. 945. [Google Scholar]
- Ginet, R.; Decou, V. Initiation à la Biologie et à L’écologie Souterraines; Jean-Pierre Delarge: Paris, France, 1977; p. 345. [Google Scholar]
- Gibert, J.; Deharveng, L. Subterranean Ecosystems: A Truncated Functional Biodiversity. BioScience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Martínez, A.; Di Domenico, M.; Worsaae, K. Evolution of cave Axiokebuita and Speleobregma (Scalibregmatidae, Annelida). Zool. Scr. 2013, 42, 623–636. [Google Scholar] [CrossRef]
- Martínez, A.; Gonzales, B.C.; Worsaae, K.; Wilkens, H.; Núñez Jiménez, A.; Oromi, P.; Iliffe, T.M. Guide to the Anchialine Ecosystems of Jameos del Agua and Túnel de la Atlántida; Cabildo de Lanzarote, Medio Ambiente: Arrecife, Lanzarote, 2016. [Google Scholar]
- Worsaae, K.; Sterrer, W.; Iliffe, T.M. Longipalpa saltatrix, a new genus and species of the meiofaunal family Nerillidae (Annelida: Polychaeta) from an anchihaline cave in Bermuda. Proc. Biol. Soc. Wash. 2004, 117, 346–362. [Google Scholar]
- Alves, P.R.; Glasby, C.J.; Santos, C.S.G. On the use of troglomorphic characters in Namanereidinae (Annelida; Nereididae) systematics. Zootaxa 2018, 4531, 195–210. [Google Scholar] [CrossRef]
- Matjašič, J.; Sket, B. Développement larvaire du Serpulien cavernicole Marifugia cavatica Absolon et Hrabe (Polychaeta, Sedentaria). Speleology 1966, 2, 9–16. [Google Scholar] [CrossRef]
- Worsaae, K. 2021. 7.9 Errantia incertae sedis: Nerillidae Levinsen. In Handbook of Zoology, Pleistoannelida, Sedentaria III and Errantia I; Purschke, G., Böggemann, M., and W. Westheide, W., Eds.; De Gruyter: Berlin, Germany, 1883; pp. 215–227. [Google Scholar] [CrossRef]
- Creuzé des Châtelliers, M.; Juget, J.; Lafont, M.; Martin, P. Subterranean aquatic Oligochaeta. Freshw. Biol. 2009, 54, 678–690. [Google Scholar] [CrossRef]
- Kerovec, M.; Kučinić, M.; Jalžic, B. Croatobranchus mestrovi sp. n. -predstavnik nove endemske podzemne vrste pijavica. Spelolog 1999, 44–45, 35–36. [Google Scholar]
- Sket, B.; Dovc, P.; Jalzic, B.; Kerovec, M.; Kucinic, M.; Trontelj, P. A cave leech (Hirudinea, Erpobdellidae) from Croatia with unique morphological features. Zool. Scr. 2001, 30, 223–229. [Google Scholar] [CrossRef]
- Timm, T. Profundal oligochaete assemblages in Palearctic lakes. Turk. J. Zool. 2012, 36, 121–131. [Google Scholar] [CrossRef]
- Giani, N.; Martin, P.; Juget, J. A new species of Phreodrilidae (Oligochaeta), Astacopsidrilus naceri sp. nov., from Morocco (North Africa), with notes on the biogeography of the family. Can. J. Zool. 1995, 73, 2375–2381. [Google Scholar] [CrossRef]
- Martínez-Ansemil, E.; Giani, N.; Sambugar, B. Oligochaetes from underground waters of Oman with descriptions of two new species of Phreodrilidae (Oligochaeta): Antarctodrilus arabicus n. sp. and Phreodrilus stocki n. sp. Contrib. Zool. 2002, 71, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Martínez, A.; Di Domenico, M.; Jörger, K.; Norenburg, J.; Worsaae, K. Description of three new species of Protodrilus (Annelida, Protodrilidae) from Central America. Mar. Biol. Res. 2013, 9, 676–691. [Google Scholar] [CrossRef]
- Martínez, A.; Di Domenico, M.; Rouse, G.W.; Worsaae, K. Phylogeny and systematics of Protodrilidae (Annelida) inferred with total evidence analyses. Cladistics 2015, 31, 250–276. [Google Scholar] [CrossRef]
- Di Domenico, M.; da Cunha Lana, P.; Garraffoni, A.R.S. Distribution patterns on interstitial polychaetes in sandy beaches of southern Brazil. Mar. Ecol. 2008, 30, 47–62. [Google Scholar] [CrossRef]
- Di Domenico, M.; Martínez, A.; Lana, P.C.; Worsaae, K. Protodrilus (Protodrilidae, Annelida) from the southern and southeastern Brazilian coasts. Helgol. Mar. Res. 2013, 67, 733–748. [Google Scholar] [CrossRef] [Green Version]
- Paxton, H. Taxonomy and aspects of the life history of Australian beachworms (Polychaeta: Onuphidae). Aust. J. Mar. Freshw. Res. 1979, 30, 265–294. [Google Scholar] [CrossRef]
- Kirtley, D.W. Intertidal Reefs of Sabellariidae, Annelida Polychaeta, along the Coast of Florida. Ph.D. Thesis, The Florida State University, Tallahassee, FL, USA, 1966. [Google Scholar]
- Vovelle, J. Le tube de Sabellaria alveolata (L.) annélide polychète Hermellidae et son ciment étude ecologique, expérimentale, histologique et histochimique. Arch. Zool. Exp. Gén. 1965, 106, 1–187. [Google Scholar]
- Gaill, F.; Hunt, S. Tubes. In The Ultrastructure of Polychaeta; Westheide, W., Hermans, C.O., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 1988; Volume 4, pp. 61–70. [Google Scholar]
- Wang, C.S.; Svendsen, K.K.; Stewart, R.J. Morphology of the adhesive system in the sandcastle worm, Phragmatopoma californica. In Biological Adhesive Systems: From Nature to Technical and Medical Application; von Byern, J., Grunwald, I., Eds.; Springer: Vienna, Austria, 2010; pp. 169–179. [Google Scholar] [CrossRef]
- Morgado, E.H.; Tanaka, M.O. The macrofauna associated with the bryozoan Schizoporella unicornis in southeastern Brazil. Sci. Mar. 2007, 65, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Pérez, C.D.; Vila-Nova, D.A.; Santos, A.M. Associated community with the zoanthid Palythoa caribaeorum (Duchassaing & Michelotti, 1860) (Cnidaria, Anthozoa) from littoral of Pernambuco, Brazil. Hydrobiologia 2005, 548, 207–215. [Google Scholar] [CrossRef]
- Yamanishi, R. Notes on Pholoe parva Imajima & Hartman, 1964 (Annelida: Polychaeta: Sigalionidae). Bull. Osaka Mus. Nat. Hist. 1985, 38, 17–22. [Google Scholar]
- Swedmark, B. The interstitial fauna of marine sand. Biol. Rev. 1964, 39, 1–42. [Google Scholar] [CrossRef]
- Worsaae, K.; Kristensen, R.M. Evolution of interstitial Polychaeta (Annelida). Hydrobiologia 2005, 535–536, 319–340. [Google Scholar]
- Westheide, W. Progenesis as a principle in meiofauna evolution. J. Nat. Hist. 1987, 21, 843–854. [Google Scholar] [CrossRef]
- Struck, T.H.; Golombek, A.; Weigert, A.; Franke, F.A.; Westheide, W.; Purschke, W.; Bleidorn, C.; Halanych, K.M. The evolution of annelids reveals two adaptive routes to the interstitial realm. Curr. Biol. 2015, 25, 1993–1999. [Google Scholar] [CrossRef] [Green Version]
- Giani, N.; Lavandier, P. Les oligochètes du torrent d′Estaragne (Pyrenées Centrales). Bull. Soc. Hist. Nat. Toulouse 1977, 113, 234–243. [Google Scholar]
- Martínez-Ansemil, E.; Collado, R. Distribution patterns of aquatic oligochaetes inhabiting watercourses in the Northwestern Iberian Peninsula. Hydrobiologia 1996, 334, 73–83. [Google Scholar] [CrossRef]
- Gagneur, J.; Giani, N.; Martínez-Ansemil, E. Les oligochètes aquatiques d′Algérie. Bull. Soc. Hist. Nat. Toulouse 1986, 122, 119–124. [Google Scholar]
- Tsutsumi, H. Population dynamics of Capitella capitata (Polychaeta; Capitellidae) in an organically polluted cove. Mar. Ecol. Prog. Ser. 1987, 36, 139–149. [Google Scholar] [CrossRef]
- Dean, H.K. The use of polychaetes (Annelida) as indicator species of marine pollution: A review. Rev. Biol. Trop. 2008, 56, 11–38. [Google Scholar]
- Forbes, T.L.; Forbes, V.E.; Depledge, M.H. Individual physiological responses to environmental hypoxia and organic enrichment: Implications for early soft-bottom community succession. J. Mar. Res. 1994, 52, 1081–1100. [Google Scholar] [CrossRef]
- Warren, L.M. Respiratory adaptations to temporary hypoxia by the polychaete Cirriformia tentaculata. Comp. Biochem. Physiol. Part A Physiol. 1981, 69, 321–324. [Google Scholar] [CrossRef]
- Petersen, M.E.; George, J.D. A new species of Raricirrus from northern Europe, with notes on its biology and a discussion of the affinities of the genus (Polychaeta: Ctenodrilidae). In Systematics, Biology and Morphology of World Polychaeta (Ophelia Supplement 5), Proceedings of the Second International Polychaete Conference, Copenhagen, Denmark, 18–23 August 1986; Petersen, M.E., Kirkegaard, J.B., Eds.; Brill: Leiden, The Netherlands, 1991; pp. 185–208. [Google Scholar]
- Ugland, K.I.; Bjørgesæter, A.; Bakke, T.; Fredheim, B.; Gray, J.S. Assessment of environmental stress with a biological index based on opportunistic species. J. Exp. Mar. Biol. Ecol. 2008, 366, 169–174. [Google Scholar] [CrossRef]
- Cunha, M.R.; Rodrigues, C.F.; Génio, L.; Hilário, A.; Ravara, A.; Pfannkuche, O. Macrofaunal assemblages from mud volcanoes in the Gulf of Cadiz: Abundance, biodiversity and diversity partitioning across spatial scales. Biogeosciences 2013, 10, 2553–2568. [Google Scholar] [CrossRef] [Green Version]
- Glasby, C.J.; Hutchings, P.A. A new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicida: Eunicidae) from northern Australia and a review of similar taxa from the Indo-west Pacific, including the genus Nauphanta Kinberg, 1865. Zootaxa 2010, 2352, 29–45. [Google Scholar] [CrossRef]
- Brinkhurst, R.O. Distribution and Ecology. 1. Microdriles. In Aquatic Oligochaeta of the World; Brinkhurst, R.O., Jamieson, A.J., Eds.; Oliver & Boyd: Edinburgh, Scotland, 1971; pp. 104–146. [Google Scholar]
- Rodriguez, P.; Reynoldson, T.B. The Pollution Biology of Aquatic Oligochaetes; Springer: Dordrecht, The Netherlands, 2011; p. 265. [Google Scholar]
- Giere, O.; Preusse, J.-H.; Dubilier, N. Tubificoides benedii (Tubificidae, Oligochaeta)—a pioneer in hypoxic and sulfidic environments. An overview of adaptative pathways. Hydrobiologia 1999, 406, 235–241. [Google Scholar] [CrossRef]
- Ito, K.; Ito, M.; Hano, T.; Onduka, T.; Mochida, K.; Shono, N.; Nakamura, R. Marine sediment conservation using benthic organisms. Bull. Jpn. Fish. Res. Educ. Agency 2019, 49, 45–51. [Google Scholar]
- Dubilier, N.; Blazejak, A.; Rühland, C. Symbioses between bacteria and gutless marine oligochaetes. In Molecular Basis of Symbiosis; Overmann, J., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 251–275. [Google Scholar] [CrossRef]
- Hourdez, S.; Frederick, L.-A.; Schernecke, A.; Fisher, C.R. Functional respiratory anatomy of a deep-sea orbiniid polychaete from the Brine Pool NR-1 in the Gulf of Mexico. Invertebr. Biol. 2005, 120, 29–40. [Google Scholar] [CrossRef]
- Hourdez, S.; Weber, R.E.; Green, B.N.; Kenney, J.M.; Fisher, C.R. Respiratory adaptations in a deep-sea orbiniid polychaete from Gulf of Mexico brine pool NR-1: Metabolic rates and hemoglobin structure/function relationships. J. Exp. Biol. 2002, 205, 1669–1681. [Google Scholar] [PubMed]
- Weber, R.E.; Vinogradov, S.N. Nonvertebrate hemoglobins: Functions and molecular adaptations. Physiol. Rev. 2001, 81, 569–628. [Google Scholar] [CrossRef] [Green Version]
- Narita, T. Seasonal vertical migration and eastivation of Rhyacodrilus hiemalis (Tubificidae, Clitellata) in the sediment of Lake Biwa, Japan. Hydrobiologia 2006, 564, 87–93. [Google Scholar] [CrossRef]
- Southward, E.C. Class Pogonophora. In Polychaetes & Allies. The Southern Synthesis. Fauna of Australia Vol 4A Polychaeta, Myzostomida, Pogonophora, Echiura, Sipuncula; Beesley, P.L., Ross, G.J.B., Glasby, C.J., Eds.; CSIRO Publishing: Melbourne, Australia, 2000; pp. 331–351. [Google Scholar]
- Desbruyères, D.; Laubier, L. Les Alvinellidae, une famille nouvelle d′annélides polychètes inféodées aux sources hydrothermales sous-marines: Systématique, biologie et écologie. Can. J. Zool. 1986, 64, 2227–2245. [Google Scholar] [CrossRef]
- Tunnicliffe, V.; Desbruyères, D.; Jollivet, D.; Laubier, L. Systematic and ecological characteristics of Paralvinella sulfincola Desbruyères and Laubier, a new polychaete (family Alvinellidae) from northeast Pacific hydrothermal vents. Can. J. Zool. 1993, 71, 286–297. [Google Scholar] [CrossRef]
- Bris, N.L.; Gaill, F. Microbial habitats associated with deep-sea hydrothermal vent invertebrates: Insights from microanalysis and geochemical modeling. In The Vent and Seep Biota: Aspects from Microbes to Ecosystems; Kiel, S., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 51–71. [Google Scholar] [CrossRef]
- Terwilliger, N.B.; Terwilliger, R.C. Hemoglobin from the “pompeii worm,” Alvinella pompejana, an annelid from a deep sea hot hydrothermal vent environment. Mar. Biol. Lett. 1984, 5, 191–201. [Google Scholar]
- Toulmond, A.; Slitine, F.E.I.; De Frescheville, J.; Jouin, C. Extracellular hemoglobins of hydrothermal vent annelids: Structural and functional characteristics in three alvinellid species. Biol. Bull. 1990, 179, 366–373. [Google Scholar] [CrossRef]
- Hourdez, S.; Weber, R.E. Molecular and functional adaptations in deep-sea hemoglobins. J. Inorg. Biochem. 2005, 99, 130–141. [Google Scholar] [CrossRef]
- Desbruyères, D.; Chevaldonné, P.; Alayse, A.M.; Jollivet, D.; Lallier, F.H.; Jouin-Toulmond, C.; Zal, F.; Sarradin, P.M.; Cosson, R.; Caprais, J.C.; et al. Biology and ecology of the “Pompeii worm” (Alvinella pompejana Desbruyères and Laubier), a normal dweller of an extreme deep-sea environment: A synthesis of current knowledge and recent developments. Deep Sea Res. Part II Top. Stud. Oceanogr. 1998, 45, 383–422. [Google Scholar] [CrossRef]
- Zimmermann, J.; Wentrup, C.; Sadowski, M.; Blazejak, A.; Gruber-Vodicka, H.R.; Kleiner, M.; Ott, J.A.; Cronholm, B.; De Wit, P.; Erseus, C.; et al. Closely coupled evolutionary history of ecto- and endosymbionts from two distantly related animal phyla. Mol. Ecol. 2016, 25, 3203–3223. [Google Scholar] [CrossRef] [Green Version]
- Dubilier, N.; Mulders, C.; Ferdelman, T.; de Beer, D.; Pernthaler, A.; Klein, M.; Wagner, M.; Erseus, C.; Thiermann, F.; Krieger, J.; et al. Endosymbiotic sulphate-reducing and sulphide-oxidizing bacteria in an oligochaete worm. Nature 2001, 411, 298–302. [Google Scholar] [CrossRef]
- Glasby, C.J. The Namanereidinae (Polychaeta: Nereididae). Part 1. Taxonomy and phylogeny. Rec. Aust. Mus. Suppl. 1999, 25, 1–129. [Google Scholar] [CrossRef] [Green Version]
- Pinder, A.M.; Brinkhurst, R.O. Review of the Phreodrilidae (Annelida: Oligochaeta: Tubificida) of Australia. Invertebr. Taxon. 1997, 11, 443–523. [Google Scholar] [CrossRef]
- Ward, T.J.; Hutchings, P.A. Effects of trace metals on infaunal species composition in polluted intertidal and subtidal marine sediments near a lead smelter, Spencer Gulf, South Australia. Mar. Ecol. Prog. Ser. 1996, 135, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Grassle, J.F.; Grassle, J.P. Opportunistic life histories and genetic systems in marine benthic polychaetes. J. Mar. Res. 1974, 32, 253–284. [Google Scholar]
- Ito, K.; Ito, M.; Onduka, T.; Ohta, K.; Torii, T.; Hano, T.; Mochida, K.; Ohkubo, N.; Miura, T.; Fujii, K. Differences in the ability of two marine annelid species, Thalassodrilides sp. and Perinereis nuntia, to detoxify 1-nitronaphthalene. Chemosphere 2016, 151, 339–344. [Google Scholar] [CrossRef]
- Ito, M.; Ito, K.; Ohta, K.; Hano, T.; Onduka, T.; Mochida, K. Transcription of a novel P450 gene varies with some factors (pollutant exposure, temperature, time, and body region) in a marine oligochaete (Thalassodrilides sp.). Mar. Pollut. Bull. 2016, 109, 344–349. [Google Scholar] [CrossRef]
- Kempe, S.; Khoo, F.; Guryelik, Y. Hydrography of Lake Van and its drainage area. In The Geology of Lake Van; Degens, E.T., Kurtman, F., Eds.; MTA: Ankara, Turkey, 1978; pp. 30–44. [Google Scholar]
- Litt, T.; Krastel, S.; Sturm, M.; Kipfer, R.; Örcen, S.; Heumann, G.; Franz, S.O.; Ülgen, U.B.; Niessen, F. ‘PALEOVAN’, International Continental Scientific Drilling Program (ICDP): Site survey results and perspectives. Quat. Sci. Rev. 2009, 28, 1555–1567. [Google Scholar] [CrossRef]
- Arslan, N.; Timm, T.; Rojo, V.; VizcaIno, A.; Schmelz, R.M. A new species of Enchytraeus (Enchytraeidae, Oligochaeta) from the profundal of Lake Van, the world′s largest soda Lake (Turkey, East Anatolia). Zootaxa 2018, 4382, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Erséus, C.; Klinth, M.J.; Rota, E.; De Wit, P.; Gustafsson, D.R.; Martinsson, S. The popular model annelid Enchytraeus albidus is only one species in a complex of seashore white worms (Clitellata, Enchytraeidae). Org. Divers. Evol. 2019, 19, 105–133. [Google Scholar] [CrossRef] [Green Version]
- Hrabě, S. Die Oligochaeten des Issykkulsees. (The oligochaetes of Lake Issyk-Kul. In Russian with German summary). Tr. Kirg. Kompleks. Exped. Akad. Nauk SSSR 1935, 3, 73–85. [Google Scholar]
- Timm, T. Enchytraeids (Oligochaeta, Enchytraeidae) as lake-dwellers. In Newsletter on Enchytraeidae No 5, Proceedings of the Symposium on Enchytraeidae, Budapest, Hungary, 24–25 June 1996; Dózsa-Farkas, K., Ed.; Budapest, Hungary, 1996; pp. 47–55. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extreme Environment | Adaptation | Annelid Examples |
|---|---|---|
| Freshwater | osmoregulation | Namanereidinae (P), Fabriciidae (P), Sabellidae (P). Baikalodrilus discolor (O) |
| Land (for polychaetes) | reduced (or an absence) of nuchal organs; epidermis lacking ciliation; fewer types of receptor cells; embryos develop directly inside cocoons | Hrabeiellidae (P), Parergodrilidae (P), Nerillidae (P) |
| Deep-sea brine pools | avoidance | Methanoaricia sp. (P) |
| Hypersalinity, aridity | encysting; asexual reproduction | Manayunkia athalassia (P), Lamprodrilus mrazeki (O), some Aeolosoma species (A) |
| Frozen terrane | blood ‘antifreeze’ and other cryoprotective substances; increase internal energy levels; dehydration to lower mean supercooling point | Dinophilidae (P), Dendrobaena octaedra (O), Enchytraeus albidus (O), Mesenchytraeus solifugus (O) |
| High temperatures | protective tubes and cocoons, heat shock proteins | Eisenia spp. and Amynthas spp. (O), Siboglinidae (P) and Alvinellidae (P) |
| High-energy coasts and fast-flowing streams | small body, flat body, adhesive glands, strong tubes | many polychaete families; Trichodrilus sp. (O) |
| Subterranean | loss of eyes, pigmentation, elongation of sensory appendages (and legs), ability to cope with limited food resources | Namanereis spp. (P), Marifugia cavatica (P), Croatobranchus mestrovi (H), Trichodrilus sp. (O) |
| Low oxygen | gills, unsegmented posterior end, high O2 affinity respiratory pigments, diel and seasonal vertical migration, oxygen debts | Capitella spp. (P), Rhyacodrilus hiemalis (O), Croatobranchus mestrovi (H), Phallodrilinae (O), Aulodrilus sp. (O) |
| Deep-sea vents and cold seeps | symbiotic autotrophic bacteria, host body-cavity ‘homes’ for symbiotic bacteria, long tubes | Siboglinidae, Alvinellidae (P), genera Olavius spp. (O) Inanidrilus spp. (O), Phallodrilinae (O) |
| Pollutants (organic and inorganic) | metabolism of PAHs; assimilation of metals into hard structures, e.g., jaws; biotransformation of aromatic hydrocarbons | Capitella spp. (P). Namanereidinae (P), Thalassodrilides cf. briani (O) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glasby, C.J.; Erséus, C.; Martin, P. Annelids in Extreme Aquatic Environments: Diversity, Adaptations and Evolution. Diversity 2021, 13, 98. https://doi.org/10.3390/d13020098
Glasby CJ, Erséus C, Martin P. Annelids in Extreme Aquatic Environments: Diversity, Adaptations and Evolution. Diversity. 2021; 13(2):98. https://doi.org/10.3390/d13020098
Chicago/Turabian StyleGlasby, Christopher J., Christer Erséus, and Patrick Martin. 2021. "Annelids in Extreme Aquatic Environments: Diversity, Adaptations and Evolution" Diversity 13, no. 2: 98. https://doi.org/10.3390/d13020098
APA StyleGlasby, C. J., Erséus, C., & Martin, P. (2021). Annelids in Extreme Aquatic Environments: Diversity, Adaptations and Evolution. Diversity, 13(2), 98. https://doi.org/10.3390/d13020098