
Ocean Acidification and Mollusc Settlement in Posidonia oceanica Meadows: Does the Seagrass Buffer Lower pH Effects at CO2 Vents?

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
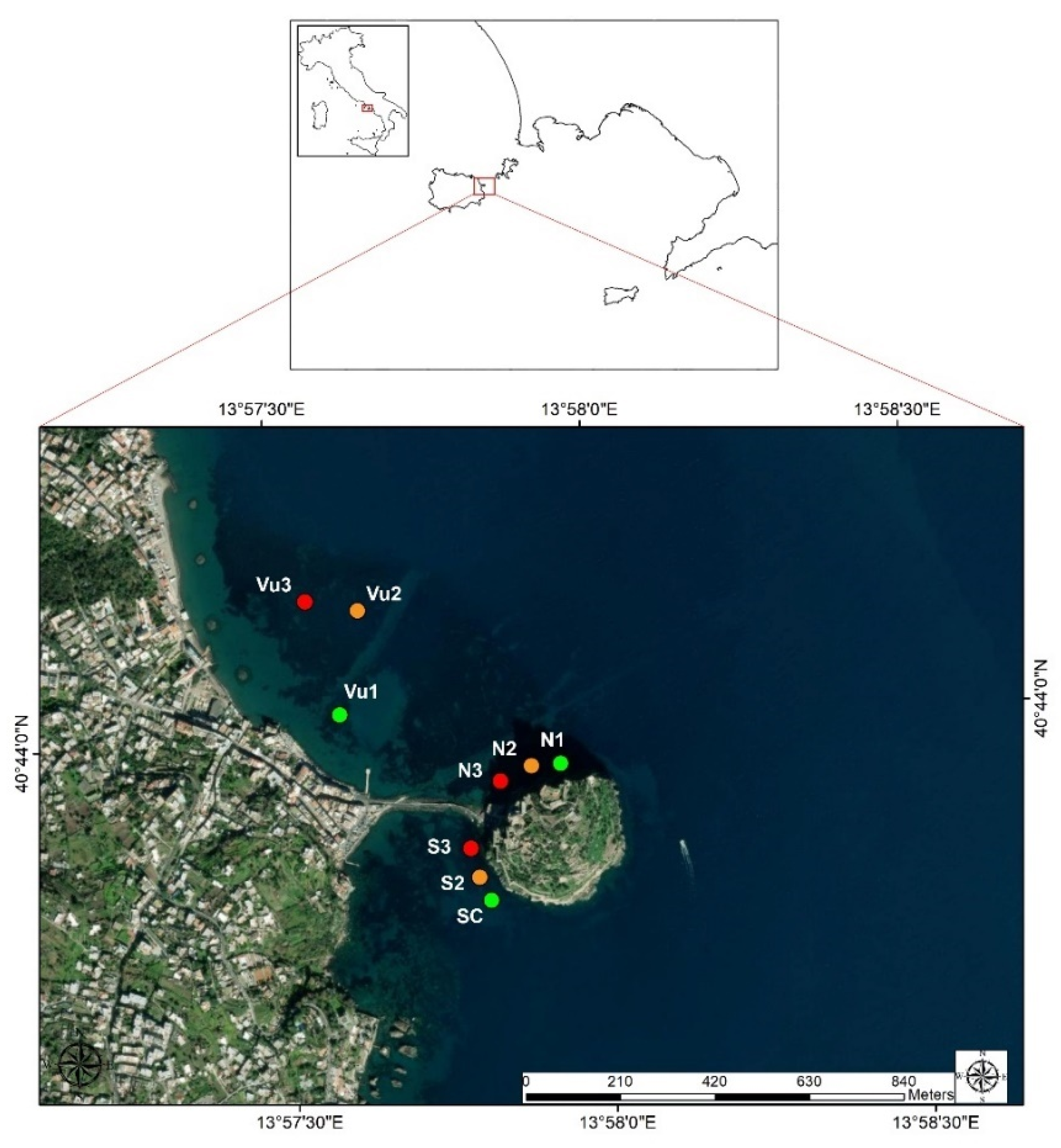
2.1. Study Area
2.2. Sampling and Data Collection
2.3. Statistical Analyses
3. Results
3.1. pH Measures
3.2. Posidonia Oceanica Leaf Phenology and Epiphytic Assemblages
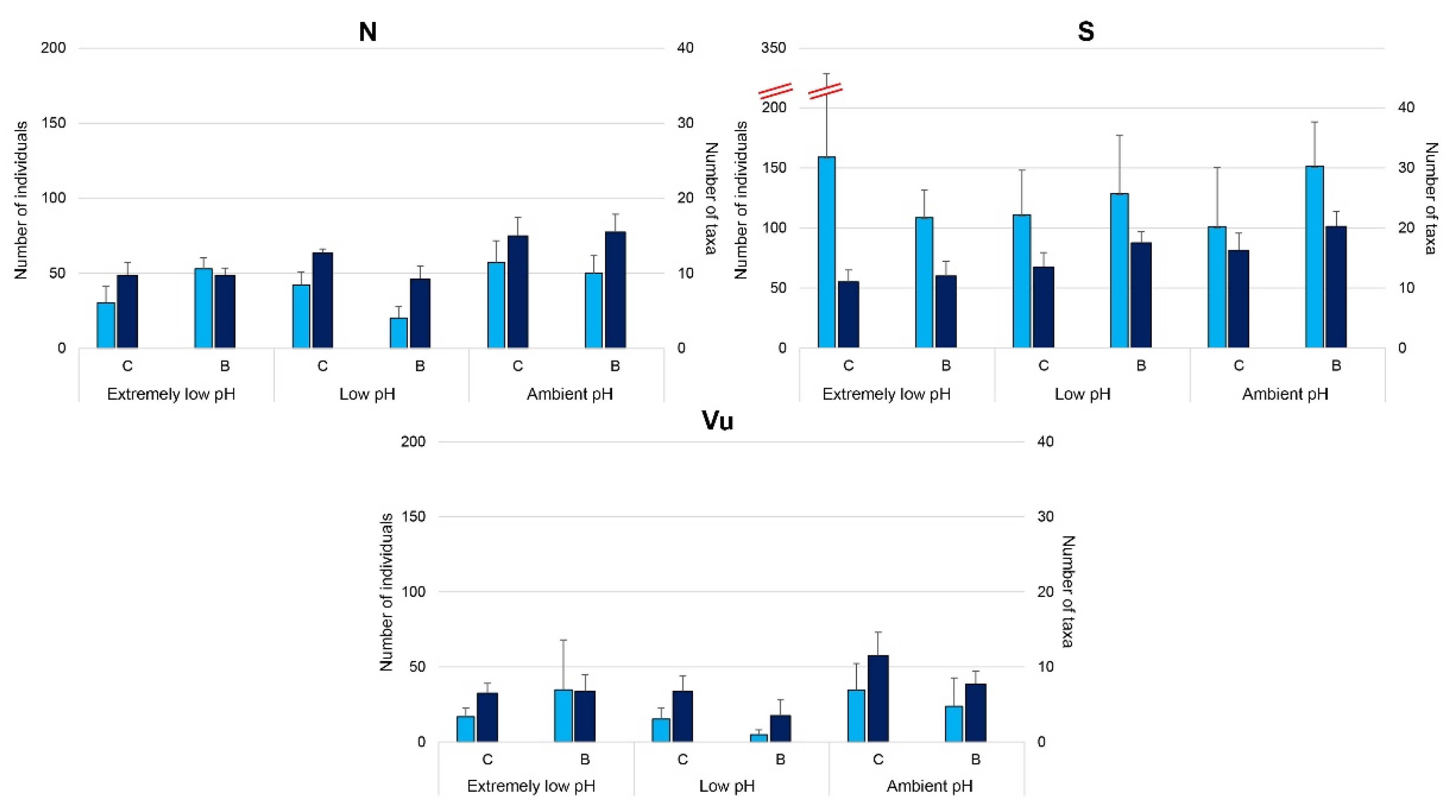
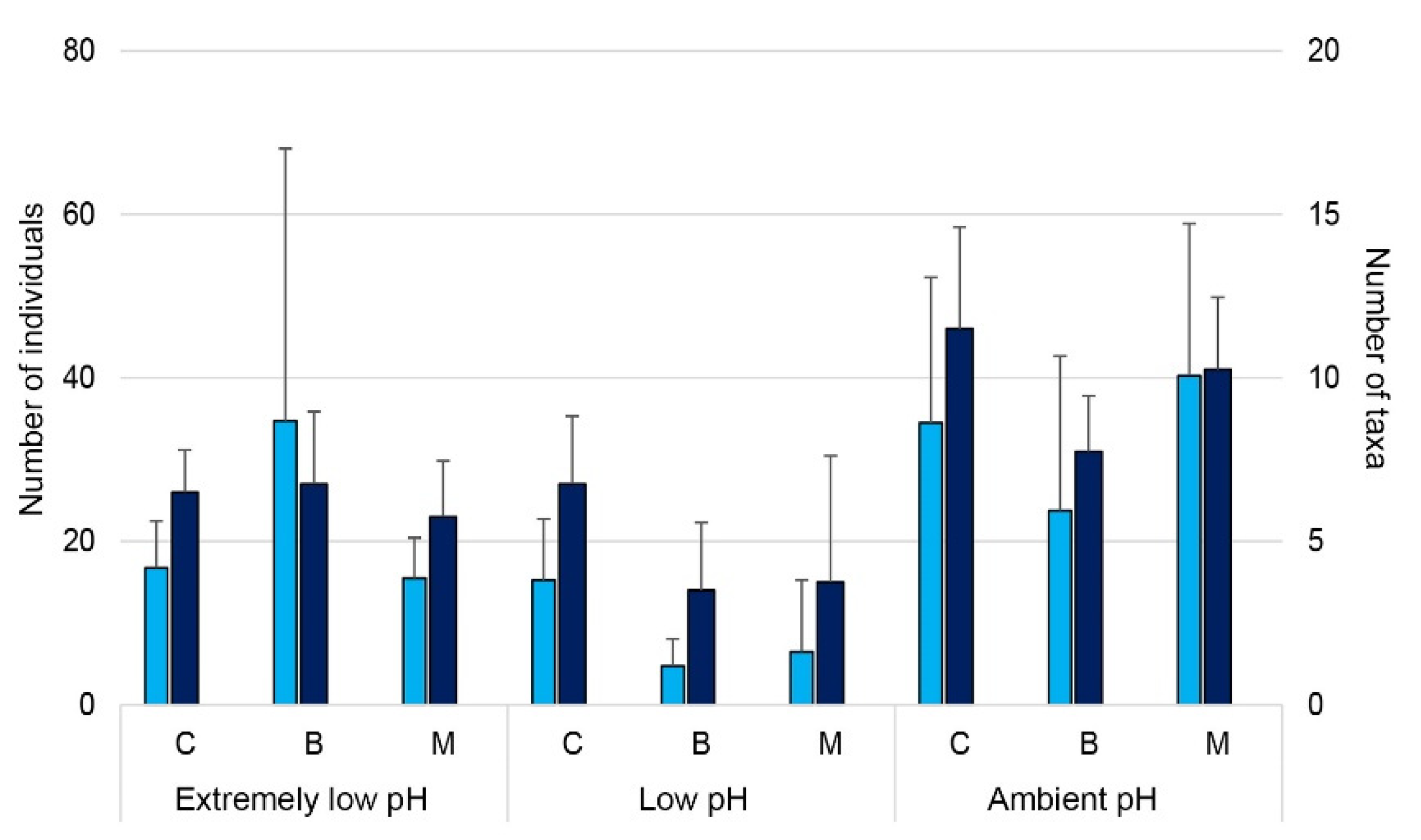
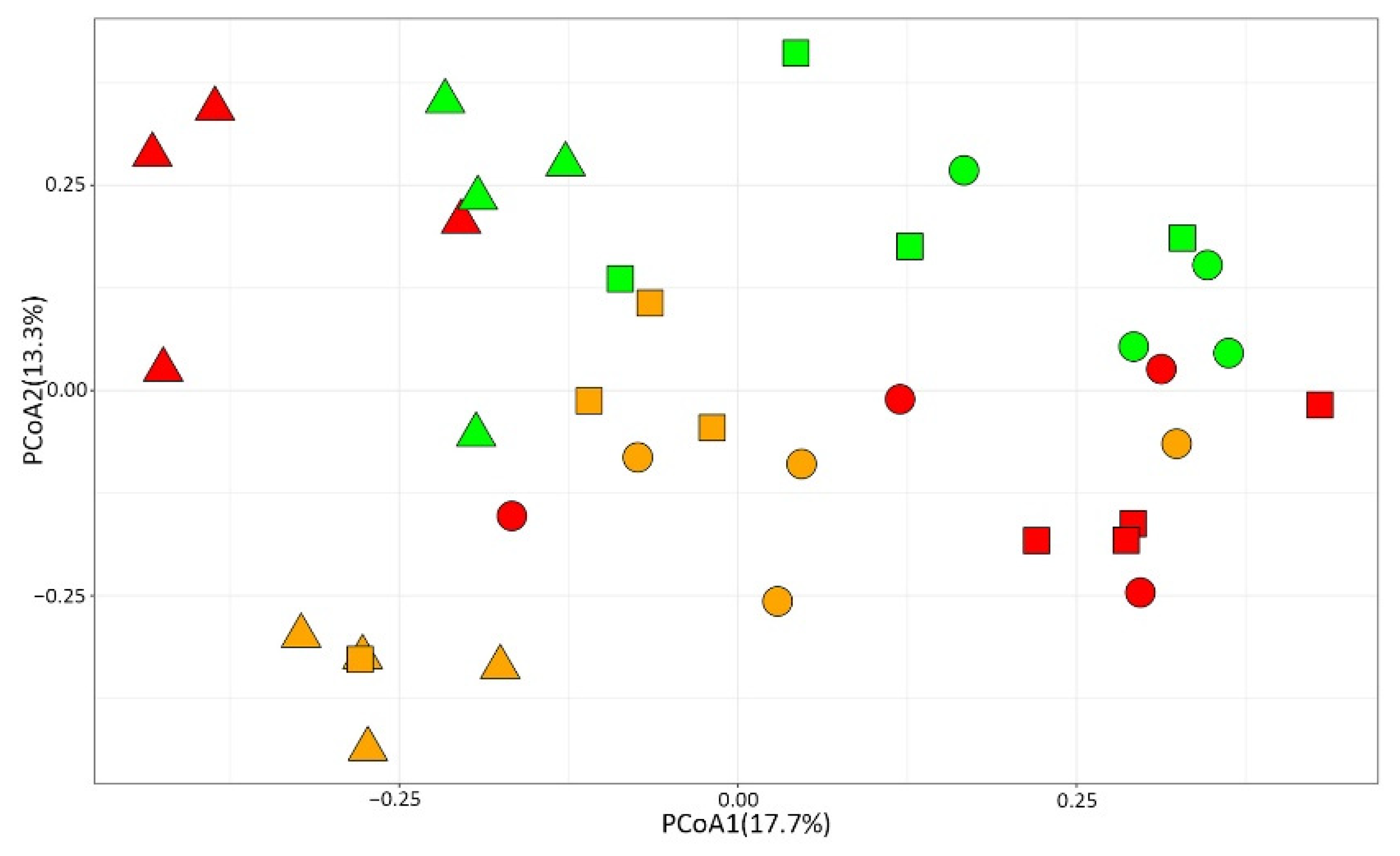
3.3. Effects of OA on Mollusc’ Settlement
3.4. Does Posidonia Oceanica Buffer the Effect of OA on Mollusc Settlement?
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shukla, P.R.; Skea, J.; Calvo Buendia, E.; Masson-Delmotte, V.; Pörtner, H.O.; Roberts, D.C.; Zhai, P.; Slade, R.; Connors, S.; Van Diemen, R. IPCC, 2019: Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019. [Google Scholar]
- Caldeira, K.; Wickett, M.E. Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef]
- Solomon, S.; Qin, D.; Manning, M.; Chen, Z.; Marquis, M.; Averyt, K.B.; Tignor, M.; Miller, H.L. Climate Change 2007: The Physical Science Basis; Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; Nykjaer, L.; Rosenberg, A.A. Cumulative human impacts on Mediterranean and Black Sea marine ecosystems: Assessing current pressures and opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabry, V.J.; Seibel, B.A.; Feely, R.A.; Orr, J.C. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES J. Mar. Sci. 2008, 65, 414–432. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.N.; Singh, G.G. Meta-analysis reveals negative yet variable effects of ocean acidification on marine organisms. Ecol. Lett. 2010, 13, 1419–1434. [Google Scholar] [CrossRef]
- Raven, J.; Caldeira, K.; Elderfield, H.; Hoegh-Guldberg, O.; Liss, P.; Riebesell, U.; Shepherd, J.; Turley, C.; Watson, A. Ocean Acidification Due to Increasing Atmospheric Carbon Dioxide; The Royal Society: London, UK, 2005; ISBN 0854036172. [Google Scholar]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J.A. Ocean acidification: The other CO2 problem. Ann. Rev. Mar. Sci. 2009, 1, 169–192. [Google Scholar] [CrossRef] [Green Version]
- Hall-Spencer, J.M.; Rodolfo-Metalpa, R.; Martin, S.; Ransome, E.; Fine, M.; Turner, S.M.; Rowley, S.J.; Tedesco, D.; Buia, M.C. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 2008, 454, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Teixidó, N.; Gambi, M.C.; Parravacini, V.; Kroeker, K.; Micheli, F.; Villéger, S.; Ballesteros, E. Functional biodiversity loss along natural CO2 gradients. Nat. Commun. 2018, 9, 5149. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, S.; Martínez-Crego, B.; Andolina, C.; Massa-Gallucci, A.; Connell, S.D.; Gambi, M.C. Ocean acidification as a driver of community simplification via the collapse of higher-order and rise of lower-order consumers. Sci. Rep. 2017, 7, 4018. [Google Scholar] [CrossRef] [Green Version]
- Auriemma, R.; De Vittor, C.; Esposito, V.; Gaglioti, M.; Gambi, M.C. Motile Fauna associated to Cystoseira brachycarpa along a gradient of Ocean Acidification at a vent system off Panarea (Aeolian Islands, Italy). Biol. Mar. Mediterr. 2019, 26, 216–219. [Google Scholar]
- Mecca, S.; Casoli, E.; Ardizzone, G.D.; Gambi, M.C. Effects of ocean acidification on phenology and epiphytes of the seagrass Posidonia oceanica at two CO2 vent systems of Ischia (Italy). Mediterr. Mar. Sci. 2020, 21, 70–83. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Micheli, F.; Gambi, M.C. Ocean acidification causes ecosystem shifts via altered competitive interactions. Nat. Clim. Chang. 2013, 3, 156–159. [Google Scholar] [CrossRef]
- Foo, S.A.; Byrne, M.; Ricevuto, E.; Gambi, M.C. The carbon dioxide vents of Ischia, Italy, a natural system to assess impacts of ocean acidification on marine ecosystems: An overview of research and comparisons with other vent systems. Oceanogr. Mar. Biol. 2018, 237–310. [Google Scholar]
- Gonzalez-Delgado, S.; Hernández, J.C. The importance of natural acidified systems in the study of ocean acidification: What have we learned? Adv. Mar. Biol. 2018, 80, 57–99. [Google Scholar] [PubMed]
- Rastrick, S.S.P.; Graham, H.; Azetsu-Scott, K.; Calosi, P.; Chierici, M.; Fransson, A.; Hop, H.; Hall-Spencer, J.; Milazzo, M.; Thor, P. Using natural analogues to investigate the effects of climate change and ocean acidification on Northern ecosystems. ICES J. Mar. Sci. 2018, 75, 2299–2311. [Google Scholar] [CrossRef]
- Cigliano, M.; Gambi, M.C.; Rodolfo-Metalpa, R.; Patti, F.P.; Hall-Spencer, J.M. Effects of ocean acidification on invertebrate settlement at volcanic CO2 vents. Mar. Biol. 2010, 157, 2489–2502. [Google Scholar] [CrossRef] [Green Version]
- Ricevuto, E.; Kroeker, K.J.; Ferrigno, F.; Micheli, F.; Gambi, M.C. Spatio-temporal variability of polychaete colonization at volcanic CO2 vents indicates high tolerance to ocean acidification. Mar. Biol. 2014, 161, 2909–2919. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Bernard, G.; Bonhomme, P.; Charbonnel, E.; Diviacco, G.; Meinesz, A.; Pergent, G.; Pergent-Martini, C.; Ruitton, S.; Tunesi, L. Protection and Conservation of Posidonia oceanica Meadows; RAMOGE, RAC/SPA and GIS Posidonie Publications: Marseille, France, 2012; ISBN 2905540311. [Google Scholar]
- Salomidi, M.; Katsanevakis, S.; Borja, Á.; Braeckman, U.; Damalas, D.; Galparsoro, I.; Mifsud, R.; Mirto, S.; Pascual, M.; Pipitone, C.; et al. Assessment of goods and services, vulnerability, and conservation status of European seabed biotopes: A stepping stone towards ecosystem-based marine spatial management. Mediterr. Mar. Sci. 2012, 13, 49–88. [Google Scholar] [CrossRef] [Green Version]
- Vassallo, P.; Paoli, C.; Rovere, A.; Montefalcone, M.; Morri, C.; Bianchi, C.N. The value of the seagrass Posidonia oceanica: A natural capital assessment. Mar. Pollut. Bull. 2013, 75, 157–167. [Google Scholar] [CrossRef]
- Mazzella, L.; Buia, M.C.; Gambi, M.C.; Lorenti, M.; Russo, G.F.; Scipione, M.B.; Zupo, V. Plant-animal trophic relationships in the Posidonia oceanica ecosystem of the Mediterranean Sea: A review. Plant-Anim. Interact. Mar. Benthos 1992, 46, 165–187. [Google Scholar]
- Giakoumi, S.; Halpern, B.S.; Michel, L.N.; Gobert, S.; Sini, M.; Boudouresque, C.F.; Gambi, M.C.; Katsanevakis, S.; Lejeune, P.; Montefalcone, M.; et al. Towards a framework for assessment and management of cumulative human impacts on marine food webs. Conserv. Biol. 2015, 29, 1228–1234. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Mainesz, A. Découverte de l’herbier de Posidonie. In Cahier du Parc National du Port-Cros; Le-Livre: Sablons, France, 1982; Volume 4, pp. 1–81. [Google Scholar]
- Pergent, G.; Bazairi, H.; Bianchi, C.N.; Boudouresque, C.F.; Buia, M.C.; Calvo, S.; Clabaut, P.; Harmelin-Vivien, M.; Mateo, M.A.; Montefalcone, M. Climate change and Mediterranean seagrass meadows: A synopsis for environmental managers. Mediterr. Mar. Sci. 2014, 15, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Mayot, N.; Boudouresque, C.F.; Leriche, A. Unexpected response of the seagrass Posidonia oceanica to a warm-water episode in the North Western Mediterranean Sea. C. R. Biol. 2005, 328, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Marbà, N.; Duarte, C.M. Mediterranean warming triggers seagrass (Posidonia oceanica) shoot mortality. Glob. Chang. Biol. 2010, 16, 2366–2375. [Google Scholar] [CrossRef]
- Mateo, M.A.; Romero, J.; Pérez, M.; Littler, M.M.; Littler, D.S. Dynamics of millenary organic deposits resulting from the growth of the Mediterranean seagrass Posidonia oceanica. Estuar. Coast. Shelf Sci. 1997, 44, 103–110. [Google Scholar] [CrossRef]
- Mateo, M.A.; Serrano, O. The carbon sink associated to Posidonia oceanica. In Mediterranean Seagrass Meadows: Resilience and Contribution to Climate Change Mitigation; IUCN: Gland, Switzerland; Málaga, Spain, 2012. [Google Scholar]
- Pergent-Martini, C.; Pergent, G.; Monnier, B.; Boudouresque, C.F.; Mori, C.; Valette-Sansevin, A. Contribution of Posidonia oceanica meadows in the context of climate change mitigation in the Mediterranean Sea. Mar. Environ. Res. 2021, 165, 105236. [Google Scholar] [CrossRef]
- Hendriks, I.E.; Olsen, Y.S.; Ramajo, L.; Basso, L.; Steckbauer, A.; Moore, T.S.; Howard, J.; Duarte, C.M. Photosynthetic activity buffers ocean acidification in seagrass meadows. Biogeosciences 2014, 11, 333–346. [Google Scholar] [CrossRef] [Green Version]
- Bergstrom, E.; Silva, J.; Martins, C.; Horta, P. Seagrass can mitigate negative ocean acidification effects on calcifying algae. Sci. Rep. 2019, 9, 1932. [Google Scholar] [CrossRef]
- Ricart, A.M.; Ward, M.; Hill, T.M.; Sanford, E.; Kroeker, K.J.; Takeshita, Y.; Merolla, S.; Shukla, P.; Ninokawa, A.T.; Elsmore, K. Coast-wide evidence of low pH amelioration by seagrass ecosystems. Glob. Chang. Biol. 2021, 27, 2580–2591. [Google Scholar] [CrossRef]
- Wahl, M.; Schneider Covachã, S.; Saderne, V.; Hiebenthal, C.; Müller, J.D.; Pansch, C.; Sawall, Y. Macroalgae may mitigate ocean acidification effects on mussel calcification by increasing pH and its fluctuations. Limnol. Oceanogr. 2018, 63, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Greiner, C.M.; Klinger, T.; Ruesink, J.L.; Barber, J.S.; Horwith, M. Habitat effects of macrophytes and shell on carbonate chemistry and juvenile clam recruitment, survival, and growth. J. Exp. Mar. Bio. Ecol. 2018, 509, 8–15. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [Green Version]
- Casoli, E.; Bonifazi, A.; Ardizzone, G.; Gravina, M.F.; Russo, G.F.; Sandulli, R.; Donnarumma, L. Comparative analysis of mollusc assemblages from different hard bottom habitats in the central Tyrrhenian sea. Diversity 2019, 11, 74. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, S.R.; Ojeda, F.P.; Inestrosa, N.C. Settlement of benthic marine invertebrates. Mar. Ecol. Prog. Ser. 1993, 97, 193–207. [Google Scholar] [CrossRef]
- Ardizzone, G.D.; Belluscio, A.; Gravina, M.F.; Somaschini, A. Colonization and disappearance of Mytilus galloprovincialis Lam. on an artificial habitat in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 1996, 43, 665–676. [Google Scholar] [CrossRef]
- Idato, E. Zonazione verticale della fauna vagile di strato foliare in una prateria di Posidonia oceanica (L.) Delile: 1. Molluschi. Boll. Malacol. 1983, 19, 109–120. [Google Scholar]
- Russo, G.F. Mollusk syntaxon of foliar stratum along a depth gradient in a Posidonia oceanica (L.) Delile meadow: Diel variability. In Proceedings of the International Workshop on Posidonia oceanica Beds, Ischia, Italie, 7–11 October 1985; GIS Posidonie Publishing: Marseille, France, 1985. [Google Scholar]
- Russo, G.F.; Fresi, E.; Buia, M.C.; Vinci, D. Malacofauna delle praterie a Posidonia oceanica della zona di Capo Passero (Sicilia Sud-Orientale): Analisi comparativa con i popolamenti dell’isola d’Ischia. Oebalia 1985, 22, 339–348. [Google Scholar]
- Gambi, M.C.; Lorenti, M.; Russo, G.F.; Scipione, M.B.; Zupo, V. Depth and seasonal distribution of some groups of the vagile fauna of the Posidonia oceanica leaf stratum: Structural and trophic analyses. Mar. Ecol. 1992, 13, 17–39. [Google Scholar] [CrossRef]
- Terlizzi, A.; Russo, G.F. Variabilità strutturale del popolamento a molluschi di alcune praterie di Posidonia oceanica del Mediterraneo. Biol. Mar. Mediterr. 1995, 2, 423–426. [Google Scholar]
- Terlizzi, A.; Russo, G.F. Analisi della dinamica annuale del taxocene a molluschi di una prateria superficiale di Posidonia oceanica: Confronto tra due diverse metodiche di campionamento. Biol. Mar. Mediterr. 1996, 3, 489–492. [Google Scholar]
- Bonfitto, A.; Fellegara, I.; Gillone, G. Sampling techniques and structure of the malacofauna associated to the rhizome zone in Posidonia oceanica (L.) Delile. Boll. Malacol. 1997, 33, 83–88. [Google Scholar]
- Martini, N.; Gillone, G.; Lombardi, C.; Sabelli, B. Mollusc community of a Posidonia oceanica (L.) Delile bed: Annual variability. Boll. Malacol. 2000, 36, 191–194. [Google Scholar]
- Beqiraj, S.; Kashta, L.; Kuci, M.; Kasemi, D.; Mato, X.; Gace, A. Benthic macrofauna of Posidonia oceanica meadows in the Albanian coast. Nat. Montenegrina 2008, 7, 55–69. [Google Scholar]
- Russo, G.F.; Terlizzi, A. Structural patterns in the mollusc assemblages of Posidonia oceanica beds: Methodological, edaphic or biogeographical product? Boll. Malacol. 1997, 33, 89–94. [Google Scholar]
- Albano, P.G.; Sabelli, B. The molluscan assemblages inhabiting the leaves and rhizomes of a deep water Posidonia oceanica settlement in the central Tyrrhenian Sea. Sci. Mar. 2012, 0, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Urra, J.; Ramírez, Á.M.; Marina, P.; Salas, C.; Gofas, S.; Rueda, J.L. Highly diverse molluscan assemblages of Posidonia oceanica meadows in northwestern Alboran Sea (W Mediterranean): Seasonal dynamics and environmental drivers. Estuar. Coast. Shelf Sci. 2013, 117, 136–147. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Micheli, F.; Gambi, M.C.; Martz, T.R. Divergent ecosystem responses within a benthic marine community to ocean acidification. Proc. Natl. Acad. Sci. USA 2011, 108, 14515–14520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambi, M.C.; Gaglioti, M.; Teixidó, N. The CO2 vent’s systems off the island of Ischia (Tyrrhenian Sea). Mem. Descr. Cart. Geol. Italia 2020, 205, 55–64. (In Italian) [Google Scholar]
- Garrard, S.L.; Gambi, M.C.; Scipione, M.B.; Patti, F.P.; Lorenti, M.; Zupo, V.; Paterson, D.M.; Buia, M.C. Indirect effects may buffer negative responses of seagrass invertebrate communities to ocean acidification. J. Exp. Mar. Bio. Ecol. 2014, 461, 31–38. [Google Scholar] [CrossRef]
- Gambi, M.C. Emissioni sommerse di CO2 lungo le coste dell’isola d’Ischia. Rilievi su altre aree come possibili laboratori naturali per lo studio dell’acidificazione e cambiamento climatico a mare. Not. SIBM 2014, 66, 67–79. [Google Scholar]
- Buia, M.C.; Gambi, M.C.; Lorenti, M.; Dappiano, M.; Zupo, V. Aggiornamento sulla distribuzione e sullo stato ambientale dei sistemi a fanerogame marine (Posidonia oceanica e Cymodocea nodosa) delle isole Flegree. Acc. Sc. Lett. Arti Napoli Mem. Soc. Sc. Fis. Mat. 2003, 5, 163–186. [Google Scholar]
- Donnarumma, L.; Lombardi, C.; Cocito, S.; Gambi, M.C. Settlement pattern of Posidonia oceanica epibionts along a gradient of ocean acidification: An approach with mimics. Mediterr. Mar. Sci. 2014, 15, 498–509. [Google Scholar] [CrossRef] [Green Version]
- Lee, J. Global Change, Response Variability, and Their Implications for Coastal Ecosystem Resilience and Functioning; Stanford University: Stanford, CA, USA, 2020. [Google Scholar]
- Ricevuto, E.; Lorenti, M.; Patti, F.P.; Scipione, M.B.; Gambi, M.C. Temporal trends of benthic invertebrate settlement along a gradient of ocean acidification at natural CO2 vents (Tyrrhenian Sea). Biol. Mar. Mediterr. 2012, 19, 49–52. [Google Scholar]
- Riebesell, U. Acid test for marine biodiversity. Nature 2008, 454, 46–47. [Google Scholar] [CrossRef]
- Cox, T.E.; Gazeau, F.; Alliouane, S.; Hendriks, I.E.; Mahacek, P.; Le Fur, A.; Gattuso, J.P. Effects of in situ CO2 enrichment on structural characteristics, photosynthesis, and growth of the Mediterranean seagrass Posidonia oceanica. Biogeosciences 2016, 13, 2179–2194. [Google Scholar] [CrossRef] [Green Version]
- Mirasole, A.; Signa, G.; Gianguzza, P.; Bonaviri, C.; Mazzola, A.; Vizzini, S. Fish assemblages cope with ocean acidification in a shallow volcanic CO2 vent benefiting from an adjacent recovery area. Mar. Environ. Res. 2020, 157, 104851. [Google Scholar] [CrossRef] [PubMed]
- Mirasole, A.; Badalamenti, F.; Di Franco, A.; Gambi, M.C.; Teixidó, N. Boosted fish abundance associated with Posidonia oceanica meadows in temperate shallow CO2 vents. Sci. Total Environ. 2021, 771, 145438. [Google Scholar] [CrossRef]
- Arnold, T.; Mealey, C.; Leahey, H.; Miller, A.W.; Hall-Spencer, J.M.; Milazzo, M.; Maers, K. Ocean acidification and the loss of phenolic substances in marine plants. PLoS ONE 2012, 7, e35107. [Google Scholar] [CrossRef] [Green Version]
- Ricevuto, E.; Vizzini, S.; Gambi, M.C. Ocean acidification effects on stable isotope signatures and trophic interactions of polychaete consumers and organic matter sources at a CO2 shallow vent system. J. Exp. Mar. Bio. Ecol. 2015, 468, 105–117. [Google Scholar]
- Scartazza, A.; Moscatello, S.; Gavrichkova, O.; Buia, M.C.; Lauteri, M.; Battistelli, A.; Lorenti, M.; Garrard, S.L.; Calfapietra, C.; Brugnoli, E. Carbon and nitrogen allocation strategy in Posidonia oceanica is altered by seawater acidification. Sci. Total Environ. 2017, 607, 954–964. [Google Scholar] [CrossRef]
- Gaglioti, M.; Mirasole, A.; Teixidó, N.; Gambi, M.C. Finestre sul futuro climatico dei nostri mari: Sintesi sulla biodiversità marina del sistema di emissione di CO2 del Castello d’Ischia. Ambiente Rischio Comun. 2019, 16, 42–47. [Google Scholar]
- Garrard, S.L. The Effect of Ocean Acidification on Plant-Animal Interactions in a Posidonia Oceanica Meadow; Open University: Milton Keynes, UK, 2013. [Google Scholar]
- Donnarumma, L.; Sandulli, R.; Appolloni, L.; Russo, G.F. Assessing molluscs functional diversity within different coastal habitats of Mediterranean marine protected areas. Ecol. Quest. 2018, 29, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Chiarore, A.; Bertocci, I.; Fioretti, S.; Meccariello, A.; Saccone, G.; Crocetta, F.; Patti, F.P. Syntopic Cystoseira taxa support different molluscan assemblages in the Gulf of Naples (southern Tyrrhenian Sea). Mar. Freshw. Res. 2019, 70, 1561–1575. [Google Scholar] [CrossRef]
- Guidetti, P. Differences among fish assemblages associated with nearshore Posidonia oceanica seagrass beds, rocky–algal reefs and unvegetated sand habitats in the Adriatic Sea. Estuar. Coast. Shelf Sci. 2000, 50, 515–529. [Google Scholar] [CrossRef]
- Gaglioti, M.; Ricevuto, E.; Gambi, M.C. Pattern and map of biodiversity related to ocean acidification in CO2 vents of Ischia. Biol. Mar. Mediterr. 2018, 25, 235–236. [Google Scholar]
- Chan, B.K.K.; Wang, T.W.; Chen, P.C.; Lin, C.W.; Chan, T.Y.; Tsang, L.M. Community structure of macrobiota and environmental parameters in shallow water hydrothermal vents off Kueishan Island, Taiwan. PLoS ONE 2016, 11, e148675. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chan, T.Y.; Chan, B.K.K. Molluscan diversity in shallow water hydrothermal vents off Kueishan Island, Taiwan. Mar. Biodivers. 2018, 48, 709–714. [Google Scholar] [CrossRef]
- Hunt, H.L.; Scheibling, R.E. Role of early post-settlement mortality in recruitment of benthic marine invertebrates. Mar. Ecol. Prog. Ser. 1997, 155, 269–301. [Google Scholar] [CrossRef] [Green Version]
- Connell, S.D.; Doubleday, Z.A.; Hamlyn, S.B.; Foster, N.R.; Harley, C.D.G.; Helmuth, B.; Kelaher, B.P.; Nagelkerken, I.; Sarà, G.; Russell, B.D. How ocean acidification can benefit calcifiers. Curr. Biol. 2017, 27, R95–R96. [Google Scholar] [CrossRef] [Green Version]
- Mutalipassi, M.; Fink, P.; Maibam, C.; Porzio, L.; Buia, M.C.; Gambi, M.C.; Patti, F.P.; Scipione, M.B.; Lorenti, M.; Zupo, V. Ocean acidification alters the responses of invertebrates to wound-activated infochemicals produced by epiphytes of the seagrass Posidonia oceanica. J. Exp. Mar. Bio. Ecol. 2020, 530–531, 151435. [Google Scholar] [CrossRef]
- Invers, O.; Zimmerman, R.C.; Alberte, R.S.; Pérez, M.; Romero, J. Inorganic carbon sources for seagrass photosynthesis: An experimental evaluation of bicarbonate use in species inhabiting temperate waters. J. Exp. Mar. Bio. Ecol. 2001, 265, 203–217. [Google Scholar] [CrossRef]
- Koch, M.; Bowes, G.; Ross, C.; Zhang, X.H. Climate change and ocean acidification effects on seagrasses and marine macroalgae. Glob. Chang. Biol. 2013, 19, 103–132. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | pH Conditions | |||||
|---|---|---|---|---|---|---|
| Class | Vu | N | S | Control | Low pH | Extremely Low pH |
| Polychaeta | 36 | 36 | 33 | 37 | 32 | 33 |
| Amphipoda | 24 | 22 | 26 | 19 | 31 | 25 |
| Tanaidacea | 9 | 7 | 4 | 5 | 5 | 6 |
| Gastropoda | 17 | 22 | 10 | 17 | 12 | 12 |
| Bivalvia | 7 | 8 | 20 | 15 | 13 | 18 |
| Decapoda | 0 | 0 | 0 | 0 | 0 | 0 |
| Cumacea | 1 | 1 | 1 | 2 | 1 | 1 |
| Isopoda | 1 | 2 | 5 | 4 | 4 | 3 |
| Echinodermata | 5 | 2 | 1 | 1 | 2 | 2 |
| Control/Ambient pH | Low pH | Extremely Low pH | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| VuC | VuB | VuM | NC | NB | SC | SB | VuC | VuB | VuM | NC | NB | SC | SB | VuC | VuB | VuM | NC | NB | SC | SB | |
| Bivalvia | |||||||||||||||||||||
| Abra alba (W. Wood, 1802) | 1 | 4 | 7 | ||||||||||||||||||
| Acanthocardia echinata (Linnaeus, 1758) | 11 | 2 | 5 | 3 | 4 | 18 | 14 | 3 | 2 | 14 | 2 | 10 | 1 | 2 | 2 | 8 | |||||
| Anomia ephippium (Linnaeus, 1758) | 9 | 2 | 1 | 3 | 6 | 3 | 1 | 4 | |||||||||||||
| Arca noae (Linnaeus, 1758) | 9 | 1 | 1 | 5 | 1 | 1 | 2 | ||||||||||||||
| Gregariella semigranata (Reeve, 1858) | 1 | 1 | 2 | ||||||||||||||||||
| Haminoea sp. | 1 | ||||||||||||||||||||
| Hiatella arctica (Linnaeus, 1767) | 15 | 2 | 10 | 16 | 21 | 29 | 4 | 28 | 1 | 9 | 9 | 9 | 1 | 7 | 28 | 2 | |||||
| Lima sp. | 10 | 4 | 6 | 8 | 4 | 6 | 9 | 1 | 13 | 5 | 3 | 5 | 3 | 1 | |||||||
| Mimachlamys varia (Linneo, 1758) | 4 | 2 | 1 | 7 | 5 | 2 | 1 | 8 | 1 | 1 | 3 | ||||||||||
| Musculus subpictus (Cantraine, 1835) | 12 | 3 | 2 | 12 | 16 | 2 | 14 | 1 | 2 | 8 | 1 | 4 | 3 | 3 | 1 | 1 | 3 | 2 | 4 | ||
| Mytilus galloprovincialis juv. (Lamarck, 1819) | 23 | 4 | 11 | 18 | 46 | 140 | 187 | 12 | 3 | 222 | 231 | 2 | 1 | 13 | 10 | 340 | 277 | ||||
| Parvicardium sp. | 1 | 3 | 2 | 3 | 5 | ||||||||||||||||
| Pecten sp. | 5 | 2 | 1 | ||||||||||||||||||
| Venerupis sp. | 3 | 2 | 4 | 2 | 14 | 30 | 50 | 5 | 25 | 34 | 12 | 1 | 36 | 14 | |||||||
| Veneridae gen. sp. | 7 | 4 | 2 | 12 | 13 | 1 | 1 | 7 | 15 | ||||||||||||
| Gastropoda | |||||||||||||||||||||
| Alvania cimex (Linneo, 1758) | 7 | 6 | 2 | 6 | |||||||||||||||||
| Alvania discors (T.Brown, 1818) | 3 | 2 | 5 | ||||||||||||||||||
| Alvania lineata (Risso, 1826) | 8 | 47 | 24 | 2 | 17 | 1 | 20 | 2 | 6 | 1 | 8 | 15 | 62 | 76 | 25 | 18 | 2 | 26 | |||
| Alvania pagodula (Bucquoy, Dautzenberg & Dollfus, 1884) | 2 | 20 | |||||||||||||||||||
| Alvania piersmai (Moolenbeek e Hoenselaar, 1989) | 3 | 1 | 2 | 2 | 8 | ||||||||||||||||
| Alvania scabra (Filippi, 1844) | 5 | 1 | 9 | 3 | 4 | 8 | 1 | 1 | |||||||||||||
| Alvania sp. juv. | 1 | 2 | 13 | 1 | 9 | 16 | 4 | 87 | 4 | ||||||||||||
| Alvania subareolata(Monterosato, 1869) | 1 | 3 | 1 | ||||||||||||||||||
| Ammonicera fischeriana (Monterosato, 1869) | 2 | 1 | 1 | 5 | 1 | 2 | 3 | 4 | 8 | 1 | 5 | 1 | 1 | ||||||||
| Bittium latreillii (Payraudeau, 1826) | 4 | 2 | 1 | 1 | 1 | ||||||||||||||||
| Bolma rugosa (Linneo, 1767) | 1 | 5 | |||||||||||||||||||
| Cerithium vulgatum (Bruguière, 1792) | 5 | 2 | 1 | 1 | 1 | 2 | 2 | 1 | 2 | ||||||||||||
| Crisilla beniamina (Monterosato, 1884) | 8 | 3 | 2 | 7 | |||||||||||||||||
| Eatonina fulgida (J. Adams, 1797) | 2 | 7 | 11 | 3 | 1 | 3 | 16 | 4 | 14 | 7 | 4 | 1 | 19 | 22 | 9 | 24 | |||||
| Ecrobia ventrosa (Montagu, 1803) | 3 | 54 | 13 | 13 | 20 | 1 | 10 | 1 | 5 | 11 | 7 | 1 | |||||||||
| Gibberula philippii (Monterosato, 1878) | 1 | 7 | 2 | 2 | |||||||||||||||||
| Gibbula sp. | 2 | 4 | 11 | 1 | 2 | 1 | 20 | 3 | 3 | 4 | 2 | 8 | |||||||||
| Hydrobia acuta neglecta (Muus, 1963) | 2 | 2 | 1 | ||||||||||||||||||
| Jujubinus exasperatus (Pennant, 1777) | 1 | 1 | |||||||||||||||||||
| Jujubinus gravinae (Dautzenberg, 1881) | 4 | 2 | 19 | 23 | 1 | 1 | 1 | 4 | 22 | 1 | 6 | 2 | |||||||||
| Jujubinus striatus (Linnaeus, 1758) | 1 | 3 | 4 | 6 | 5 | 2 | 6 | 4 | 4 | 1 | 2 | 1 | 2 | 1 | |||||||
| Melanella petitiana (Brusina, 1869) | 1 | 1 | 18 | 2 | 1 | 1 | 1 | 15 | 3 | ||||||||||||
| Mitrella sp. | 1 | 2 | |||||||||||||||||||
| Nodulus contortus (Jeffreys, 1856) | 1 | 1 | 45 | 2 | 19 | 2 | 39 | 2 | 7 | 3 | 1 | ||||||||||
| Nudibranchia gen. sp. | 1 | 1 | 2 | 1 | |||||||||||||||||
| Phorcus sp. juv. | 1 | 6 | 9 | 45 | 101 | 2 | 6 | 4 | 44 | 42 | 3 | 8 | 13 | 6 | 20 | ||||||
| Pisinna glabrata (Megerle von Mühlfeld, 1824) | 2 | 1 | 1 | ||||||||||||||||||
| Pusillina marginata (Michaud, 1830) | 3 | 1 | 4 | 9 | 5 | 20 | 9 | 4 | 1 | 15 | 2 | 14 | 13 | 10 | 8 | 5 | |||||
| Retusa truncatula (Bruguière, 1792) | 2 | 1 | |||||||||||||||||||
| Rissoa italiensis (Verduin, 1985) | 1 | 2 | 1 | 3 | 1 | ||||||||||||||||
| Rissoa lilacina (Récluz, 1843) | 10 | 4 | 1 | 1 | 1 | 1 | |||||||||||||||
| Rissoa splendida (Eichwald, 1830) | 16 | 2 | 5 | 1 | 1 | 2 | 7 | ||||||||||||||
| Rissoa variabilis (Megerle von Mühlfeld, 1824) | 4 | 34 | 2 | 40 | 1 | 16 | 1 | 1 | 21 | 36 | 12 | 23 | 13 | 30 | 18 | 16 | 8 | ||||
| Rissoa ventricosa (Desmarest, 1814) | 1 | ||||||||||||||||||||
| Rissoa violacea (Desmarest, 1814) | 1 | ||||||||||||||||||||
| Rissoella inflata (Locard, 1892) | 6 | 3 | 3 | 6 | 4 | 5 | |||||||||||||||
| Scissurella costata (d’Orbigny, 1824) | 2 | 1 | |||||||||||||||||||
| Tricolia entomochelia (Gofas, 1994) | 1 | 2 | |||||||||||||||||||
| Tricolia landinii (Bogi & Campani, 2007) | 1 | 3 | 3 | 1 | |||||||||||||||||
| Tricolia pullus (Linneo, 1758) | 7 | 5 | 8 | 1 | 5 | 5 | 2 | ||||||||||||||
| Vitreolina philippii (Rayneval & Ponzi, 1854) | 1 | 1 | 10 | 1 | 3 | 1 | 2 | 1 | 1 | 4 | |||||||||||
| Polyplacophora | |||||||||||||||||||||
| Leptochiton scabridus (Jeffreys, 1880) | 1 | ||||||||||||||||||||
| Source of Variation | Value | SD Error | t-Value | p-Value |
|---|---|---|---|---|
| Number of taxa | ||||
| Intercept | 2.629 | 0.194 | 13.567 | <0.001 |
| Low pH | −0.355 | 0.118 | −3.009 | 0.002 |
| Extremely low pH | −0.414 | 0.120 | −3.449 | <0.001 |
| Microhabitat (C) | −0.017 | 0.107 | −0.162 | 0.871 |
| Low pH × Microhabitat (C) | 0.096 | 0.165 | 0.582 | 0.561 |
| Extremely low pH × Microhabitat (C) | −0.026 | 0.174 | −0.153 | 0.878 |
| Number of individuals | ||||
| Intercept | 4.056 | 0.419 | 9.667 | <0.001 |
| Low pH | −0.382 | 0.052 | −7.307 | <0.001 |
| Extremely low pH | −0.136 | 0.048 | −2.800 | 0.005 |
| Microhabitat (C) | −0.154 | 0.049 | −3.153 | 0.001 |
| Low pH × Microhabitat (C) | 0.248 | 0.074 | 3.338 | <0.001 |
| Extremely low pH × Microhabitat (C) | 0.171 | 0.072 | 2.379 | 0.017 |
| Source of Variation | Df | SS | MS | F | p-Value |
|---|---|---|---|---|---|
| Site | 2 | 5.376 | 0.233 | 14.840 | 0.001 |
| pH | 2 | 1.065 | 0.046 | 2.932 | 0.001 |
| Microhabitat | 1 | 2.329 | 0.101 | 12.833 | 0.001 |
| Site × pH | 4 | 1.814 | 0.078 | 2.498 | 0.001 |
| Site × Microhabitat | 2 | 0.776 | 0.033 | 2.136 | 0.003 |
| pH × Microhabitat | 2 | 0.716 | 0.031 | 1.972 | 0.010 |
| Site × pH × Microhabitat | 4 | 1.353 | 0.058 | 1.863 | 0.002 |
| Residuals | 53 | 9.623 | 0.417 |
| Source of Variation | Estimate | SD Error | t-Value | p-Value |
|---|---|---|---|---|
| Number of species | ||||
| Intercept | 2.047 | 0.179 | 11.401 | <0.001 |
| Low pH | −0.795 | 0.322 | −2.469 | 0.013 |
| Extremely low pH | −0.138 | 0.263 | −0.525 | 0.599 |
| Microhabitat (C) | 0.395 | 0.232 | 1.698 | 0.089 |
| Microhabitat (M) | 0.279 | 0.238 | 1.175 | 0.24 |
| Low pH × Microhabitat (C) | 0.262 | 0.403 | 0.650 | 0.515 |
| Extremely low pH × Microhabitat (C) | −0.432 | 0.359 | −1.202 | 0.229 |
| Low pH × Microhabitat (M) | −0.211 | 0.441 | −0.477 | 0.633 |
| Extremely low pH × Microhabitat (M) | −0.439 | 0.370 | −1.188 | 0.235 |
| Null deviance: 62.86 on 35 df Residual deviance: 30.58 on 27 df | ||||
| Number of individuals | ||||
| Intercept | 3.167 | 0.102 | 30.874 | <0.001 |
| Low pH | −1.609 | 0.251 | −6.404 | <0.001 |
| Extremely low pH | 0.380 | 0.133 | 2.859 | 0.004 |
| Microhabitat (C) | 0.373 | 0.133 | 2.801 | 0.005 |
| Microhabitat (M) | 0.527 | 0.129 | 4.078 | <0.001 |
| Low pH × Microhabitat (C) | 0.793 | 0.294 | 2.692 | 0.007 |
| Extremely low pH × Microhabitat (C) | −1.118 | 0.200 | −5.583 | <0.001 |
| Low pH × Microhabitat (M) | −0.214 | 0.328 | −0.651 | 0.515 |
| Extremely low pH × Microhabitat (M) | −1.335 | 0.200 | −6.669 | <0.001 |
| Null deviance: 513.63 on 35 df Residual deviance: 251.88 on 27 df | ||||
| Source of Variation | Df | SS | MS | F | p-Value |
|---|---|---|---|---|---|
| pH | 2 | 1.601 | 0.122 | 2.943 | 0.001 |
| Microhabitat | 2 | 1.978 | 0.151 | 3.638 | 0.001 |
| pH × Microhabitat | 4 | 2.147 | 0.164 | 1.974 | 0.010 |
| Residual | 27 | 7.341 | 0.561 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barruffo, A.; Ciaralli, L.; Ardizzone, G.; Gambi, M.C.; Casoli, E. Ocean Acidification and Mollusc Settlement in Posidonia oceanica Meadows: Does the Seagrass Buffer Lower pH Effects at CO2 Vents? Diversity 2021, 13, 311. https://doi.org/10.3390/d13070311
Barruffo A, Ciaralli L, Ardizzone G, Gambi MC, Casoli E. Ocean Acidification and Mollusc Settlement in Posidonia oceanica Meadows: Does the Seagrass Buffer Lower pH Effects at CO2 Vents? Diversity. 2021; 13(7):311. https://doi.org/10.3390/d13070311
Chicago/Turabian StyleBarruffo, Alessandra, Laura Ciaralli, Giandomenico Ardizzone, Maria Cristina Gambi, and Edoardo Casoli. 2021. "Ocean Acidification and Mollusc Settlement in Posidonia oceanica Meadows: Does the Seagrass Buffer Lower pH Effects at CO2 Vents?" Diversity 13, no. 7: 311. https://doi.org/10.3390/d13070311
APA StyleBarruffo, A., Ciaralli, L., Ardizzone, G., Gambi, M. C., & Casoli, E. (2021). Ocean Acidification and Mollusc Settlement in Posidonia oceanica Meadows: Does the Seagrass Buffer Lower pH Effects at CO2 Vents? Diversity, 13(7), 311. https://doi.org/10.3390/d13070311