
Genetic Diversity and Differentiation of Pedunculate Oak (Quercus robur L.) Populations at the Southern Margin of Its Distribution Range—Implications for Conservation

 ,
, 

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
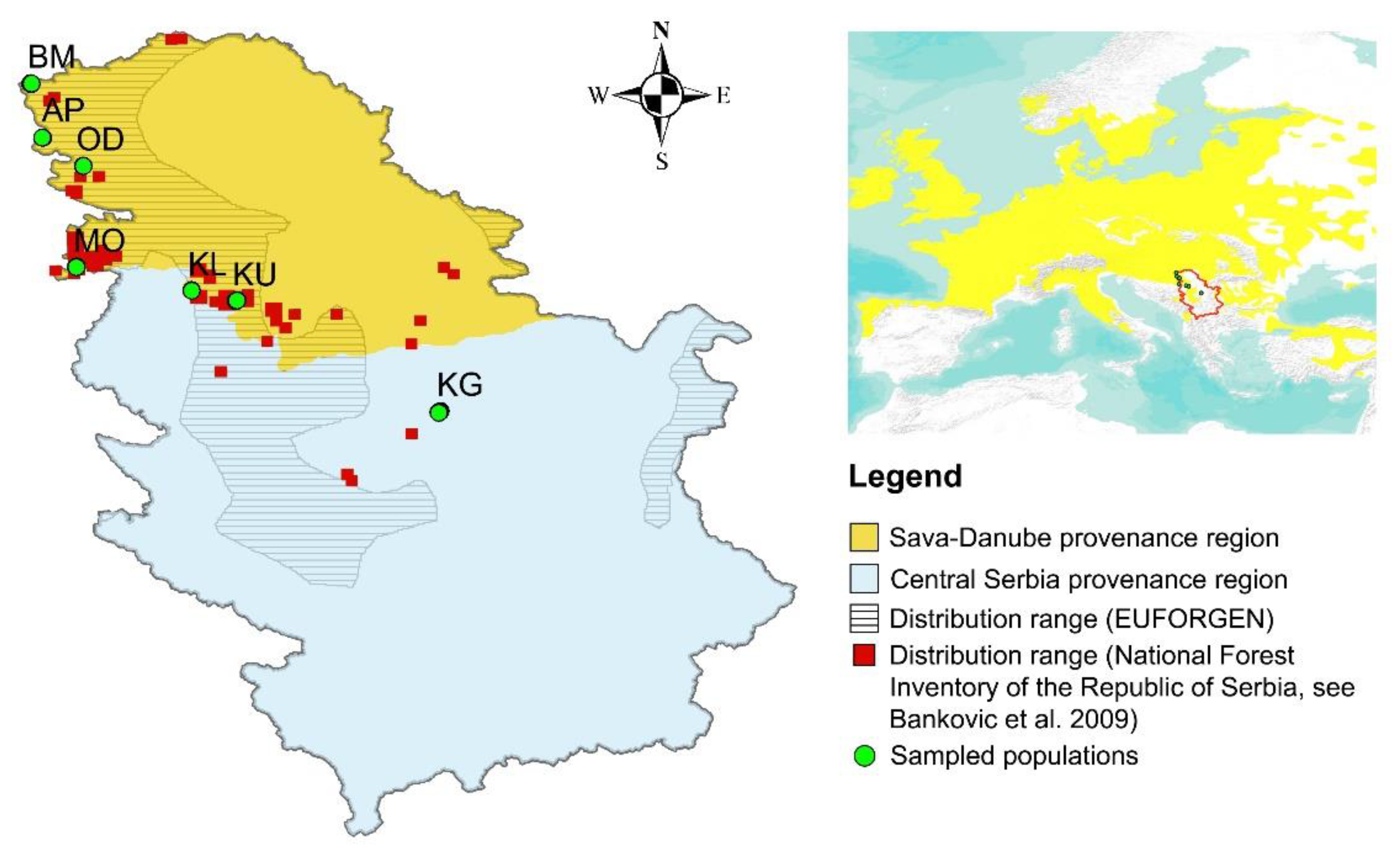
2.1. Plant Material and DNA Isolation
2.2. PCR Amplification and SSR Markers
2.3. Data Analysis
2.3.1. Variation within Population
2.3.2. Variation among Populations
3. Results
3.1. Variation and Genetic Diversity within Populations
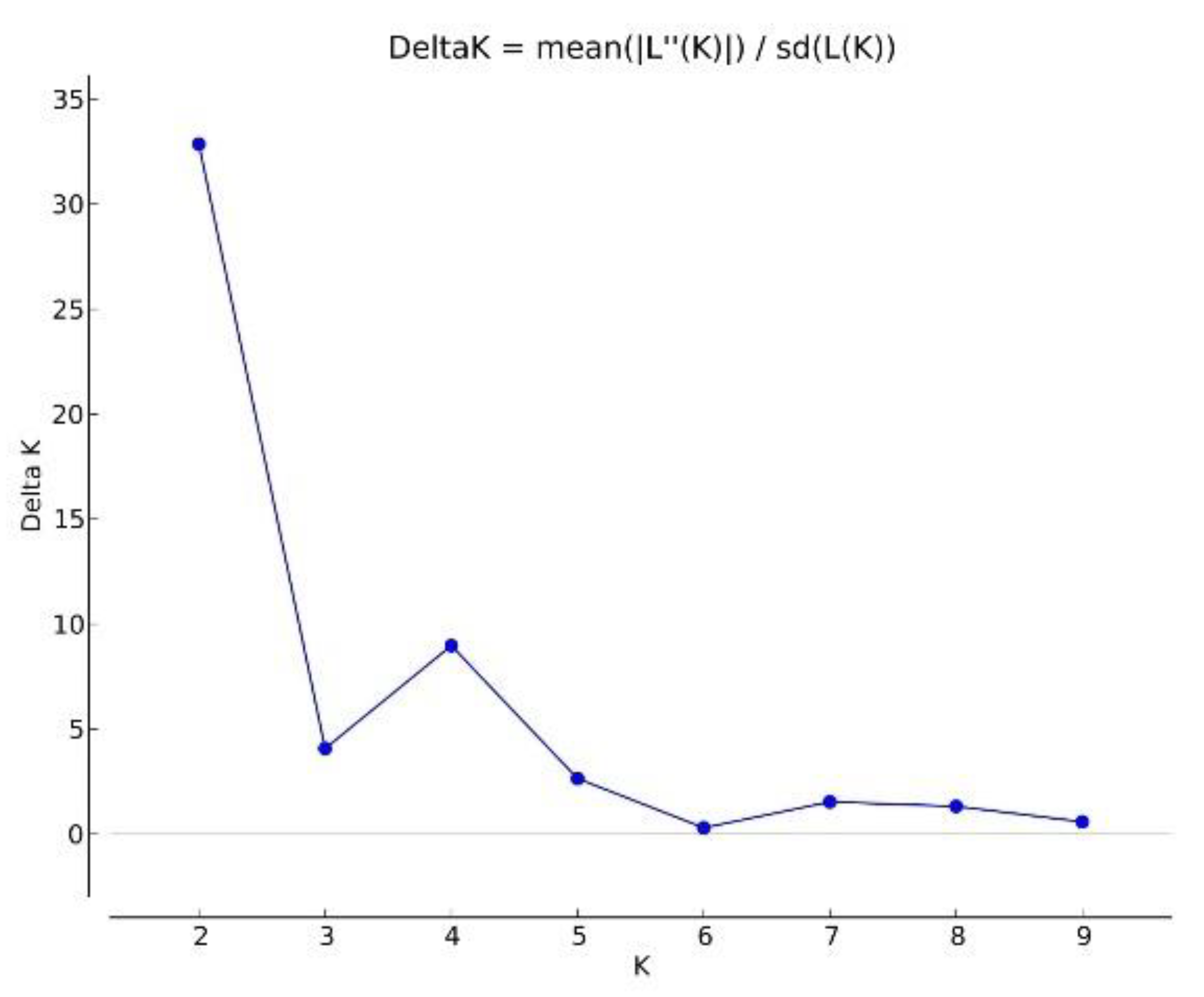
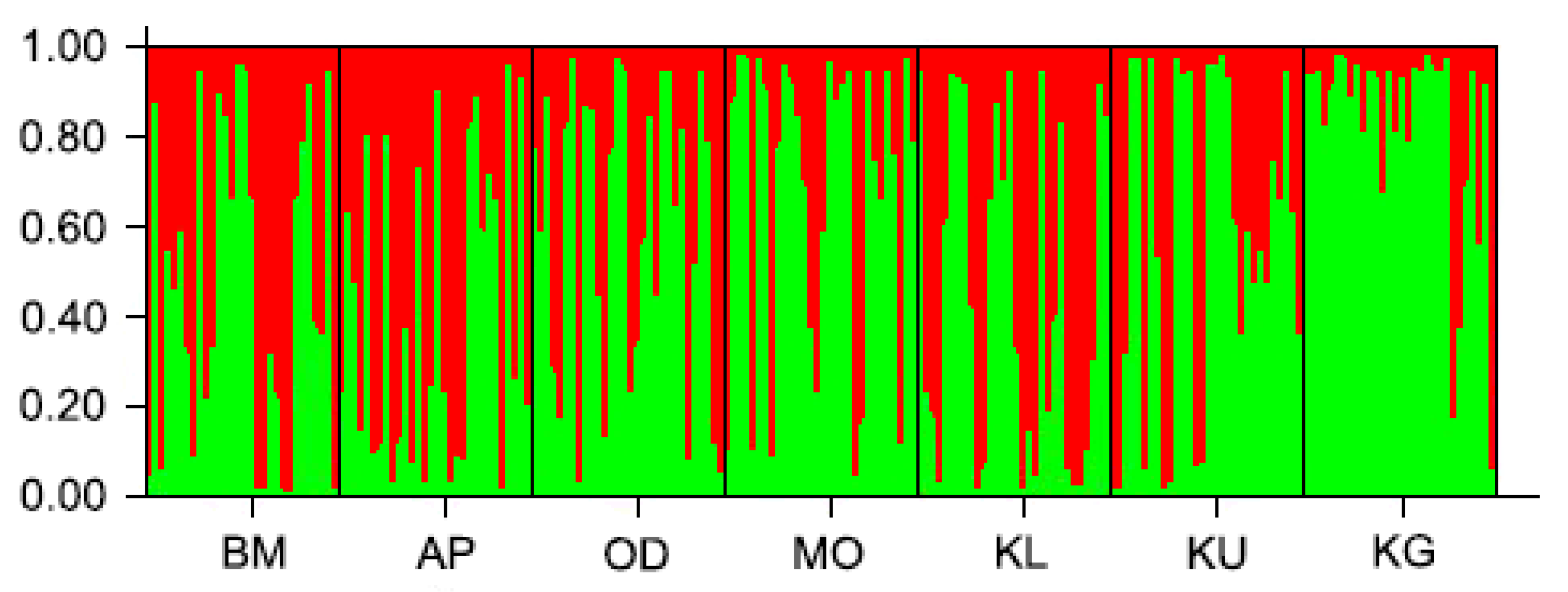
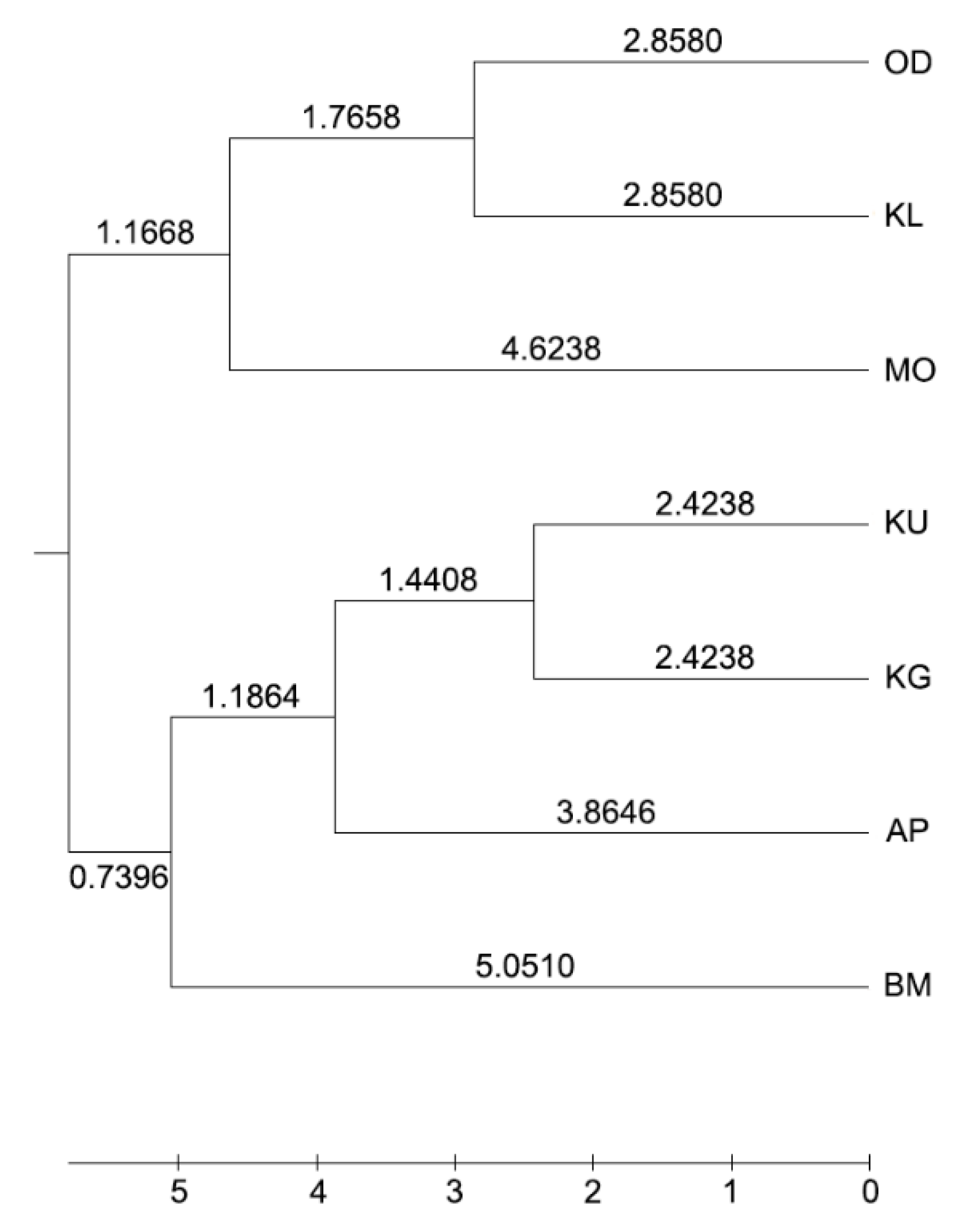
3.2. Genetic Variation between and within Populations
4. Discussion
Implications for Conservation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eaton, E.; Caudullo, G.; Oliveira, S.; de Rigo, D. Quercus robur and Quercus petraea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant., T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; p. e01c6df+. [Google Scholar]
- Birks, H.J.B.; Tinner, W. European tree dynamics and invasions during the Quaternary. In Introduced Tree Species in European Forests: Opportunities and Challenges; Krumm, F., Vítková, L., Eds.; European Forest Institute: Freiburg, Germany, 2016; pp. 22–24. [Google Scholar]
- Temunović, M.; Garnier-Géré, P.; Morić, M.; Franjić, J.; Ivanković, M.; Bogdan, S.; Hampe, A. Candidate gene SNP variation in floodplain populations of pedunculate oak (Quercus robur L.) near the species’ southern range margin: Weak differ-entiation yet distinct associations with water availability. Mol. Ecol. 2020, 29, 2359–2378. [Google Scholar] [CrossRef]
- Muir, G.; Fleming, C.C.; Schlötterer, C. Species status of hybridizing oaks. Nature 2000, 405, 1016. [Google Scholar] [CrossRef]
- Batos, B.; Vilotić, D.; Orlović, S.; Miljković, D. Inter and intra-population variation of leaf stomatal traits of Quercus robur L. in northern Serbia. Arch. Biol. Sci. 2010, 62, 1125–1136. [Google Scholar] [CrossRef]
- Ballian, D.; Memišević, M.; Bogunić, F.; Bašić, N.; Marković, M.; Kajba, D. Morphological variability of pedunculate oak (Quercus robur L.) in the region of Croatia and western Balkans. Šumarski List. 2010, 134, 371–386. [Google Scholar]
- Stojanović, D.B.; Matović, B.; Orlović, S.; Kržič, A.; Trudić, B.; Galić, Z.; Stojnić, S.; Pekeč, S. Future of the main important forest tree species in Serbia from the climate change perspective. SEEFOR 2014, 5, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Madritsch, S.; Wischnitzki, E.; Kotrade, P.; Ashoub, A.; Burg, A.; Fluch, S.; Sehr, E.M. Elucidating drought stress tolerance in European oaks through cross-species transcriptomics. G3-Genes Genom. Genet 2019, 9, 3181–3199. [Google Scholar] [CrossRef] [Green Version]
- Dobreva, M.; Simov, N.; Georgiev, G.; Mirchev, P.; Georgieva, M. First record of Corythucha arcuata (Say) (Heteroptera: Tingidae) on the Balkan Peninsula. Acta Zool. Bulgar. 2013, 65, 409–412. [Google Scholar]
- Evans, M.R. Will natural resistance result in populations of ash trees remaining in British woodlands after a century of ash dieback disease? R. Soc. Open Sci. 2019, 6, 190908. [Google Scholar] [CrossRef] [Green Version]
- Kostić, S.; Levanič, T.; Orlović, S.; Matović, B.; Stojanović, D.B. Pendunctulate and Turkey oaks radial increment and stable carbon isotope response to climate conditions through time. Poplar 2019, 204, 29–35. [Google Scholar]
- Stojanović, D.B.; Levanič, T.; Matović, B.; Orlović, S. Growth decrease and mortality of oak floodplain forests as a response to change of water regime and climate. Eur. J. Forest. Res. 2015, 134, 555–567. [Google Scholar] [CrossRef]
- Thomas, F.M.; Blank, R.; Hartmann, G. Abiotic and biotic factors and their interactions as causes of oak decline in Central Europe. For. Pathol. 2002, 32, 277–307. [Google Scholar] [CrossRef]
- Bussotti, F.; Ferretti, M. Air pollution, forest condition and forest decline in Southern Europe: An overview. Environ. Poll. 1998, 101, 49–65. [Google Scholar] [CrossRef]
- Atkins, K.E.; Travis, J.M.J. Local adaptation and the evolution of species’ ranges under climate change. J. Theor. Biol. 2010, 266, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Matesanz, S.; Guilhaumon, F.; Araújo, M.B.; Balaguer, L.; Benito-Garzón, M.; Cornwell, W.; Gianoli, E.; van Kleunen, M.; Naya, E.D.; et al. The effects of phenotypic plasticity and local ad-aptation on forecasts of species range shifts under climate change. Ecol. Lett. 2014, 17, 1351–1364. [Google Scholar] [CrossRef] [Green Version]
- Lira-Noriega, A.; Manthey, J.D. Relationship of genetic diversity and niche centrality: A survey and analysis. Evolution 2014, 68, 1082–1093. [Google Scholar] [CrossRef] [PubMed]
- Rose, L.; Leuchner, C.; Köckemann, B.; Buschmann, H. Are marginal beech (Fagus sylvatica L.) provenances a source for drought tolerant ecotypes. Eur. J. For. Res. 2009, 128, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Erichsen, E.O.; Budde, K.B.; Sagheb-Talebi, K.; Bagnoli, F.; Vendramin, G.G.; Hansen, O.K. Hyrcanian forests—Stable rear-edge populations harbouring high genetic diversity of Fraxinus excelsior, a common European tree species. Divers. Distrib. 2018, 24, 1521–1533. [Google Scholar] [CrossRef] [Green Version]
- Stojnić, S.; Suchocka, M.; Benito-Garzón, M.; Torres-Ruiz, J.M.; Cochard, H.; Bolte, A.; Delzon, S. Variation in xylem vulnerability to embolism in European beech from geographically marginal populations. Tree Physiol. 2018, 38, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Fady, B.; Aravanopoulos, F.A.; Alizoti, P.; Mátyás, C.; von Wühlisch, G.; Westergren, M.; Belletti, P.; Cvjetkovic, B.; Ducci, F.; Huber, G.; et al. Evolution-based approach needed for the conservation and silviculture of peripheral forest tree populations. For. Ecol. Manag. 2016, 375, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Holliday, J.A.; Suren, H.; Aitken, S.N. Divergent selection and heterogeneous migration rates across the range of Sitka spruce (Picea sitchensis). Proc. R. Soc. B Biol. Sci. 2012, 279, 1675–1683. [Google Scholar] [CrossRef]
- Hódar, J.A.; Castro, J.; Zamora, R. Pine processionary caterpillar Thaumetopoea pityocampa as a new threat for relict Mediterranean Scots pine forests under climatic warming. Biol. Conserv. 2003, 110, 123–129. [Google Scholar] [CrossRef]
- Bigler, C.; Bräker, O.U.; Bugmann, H.; Dobbertin, M.; Rigling, A. Drought as an inciting mortality factor in Scots pine stands of the Valais, Switzerland. Ecosystems 2006, 9, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Konnert, M.; Fady, B.; An Gomory, D.; A’hara, S.; Wolter, F.; Ducci, F.; Koskela, J.; Bozzano, M.; Maaten, T.; Kowalczyk, J. Use and transfer of forest reproductive material in Europe in the context of climate change. In European Forest Genetic Resources Programme (EUFORGEN); Bioversity International: Rome, Italy, 2015; Volume xvi, p. 75. [Google Scholar]
- Alleaume-Benharira, M.; Pen, I.R.; Ronce, O. Geographical patterns of adaptation within a species’ range: Interactions between drift and gene flow. J. Evol. Biol. 2006, 19, 203–215. [Google Scholar] [CrossRef]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Mohammad-Panah, N.; Shabanian, N.; Khadivi, A.; Rahmani, M.S.; Emami, A. Genetic structure of gall oak (Quercus in-fectoria) characterized by nuclear and chloroplast SSR markers. Tree Genet. Genom. 2017, 13, 70. [Google Scholar] [CrossRef]
- An, M.; Deng, M.; Zheng, S.S.; Jiang, X.L.; Song, Y.G. Introgression threatens the geneticdiversity of Quercus austro-cochinchinensis (Fagaceae), an endangered oak: A case inferred by molecular markers. Front. Plant Sci. 2017, 8, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carabeo, M.; Simeone, M.C.; Cherubini, M.; Mattia, C.; Chiocchini, F.; Bertini, L.; Caruso, C.; La Mantia, T.; Villani, F.; Mattioni, C. Estimating the genetic diversity and structure of Quercus trojana Webb populations in Italy by SSRs: Impli-cations for management and conservation. Can. J. For. Res. 2017, 47, 331–339. [Google Scholar] [CrossRef]
- Batos, B.; Miletić, Z.; Orlović, S.; Miljković, D. Variability of nutritive macroelements in pedunculate oak (Quercus robur L.) leaves in Serbia. Genetika 2010, 42, 435–453. [Google Scholar] [CrossRef]
- Villellas, J.; Berjano, R.; Terrab, A.; García, M.B. Divergence between phenotypic and genetic variation within populations of a common herb across Europe. Ecosphere 2014, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kampfer, S.; Lexer, C.; Glossl, J.; Steinkellner, H. Brief report characterization of (GA) n microsatellite loci from Quercus robur. Hereditas 1998, 129, 1–86. [Google Scholar] [CrossRef]
- Steinkellner, H.; Lexer, C.; Turetschek, E.; Glössl, J. Conservation of (GA) n microsatellite loci between Quercus species. Mol. Ecol. 1997, 6, 1189–1194. [Google Scholar] [CrossRef]
- Dow, B.D.; Ashley, M.V.; Howe, H.F. Characterization of highly variable (GA/CT) n microsatellites in the bur oak, Quercus macrocarpa. Theor. Appl. Genet. 1995, 91, 137–141. [Google Scholar] [CrossRef]
- Neophytou, C.; Aravanopoulos, F.A.; Fink, S.; Dounavi, A. Detecting interspecific and geographic differentiation patterns in two interfertile oak species (Quercus petraea (Matt.) Liebl. and Q. robur L.) using small sets of microsatellite markers. For. Ecol. Manag. 2010, 259, 2026–2035. [Google Scholar] [CrossRef] [Green Version]
- Yeh Francis, C.; Yang, R.; Boyle Timothy, B.; Ye, Z.; Mao, J.X. POPGENE Version 1.32, the User-Friendly Shareware for Population Genetic Analysis; Molecular Biology and Biotechnology Centre, University of Alberta: Alberta, AB, Canada, 1999; Available online: https://sites.ualberta.ca/~fyeh/popgene.html (accessed on 6 December 2020).
- Kalinowski, S.T. hp-rare 1.0: A computer program for performing rarefaction on measures of allelic richness. Mol. Ecol. Resour. 2005, 5, 187–189. [Google Scholar] [CrossRef]
- Smouse, P.E.; Banks, S.C.; Peakall, R. Converting quadratic entropy to diversity: Both animals and alleles are diverse, but some are more diverse than others. PLoS ONE 2017, 12, e0185499. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT (version 1.2): A computer program to calculate F-statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L. Maximum likelihood estimation of the frequency of null alleles at microsatellite loci. Conserv. Genet. 2006, 7, 991–995. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Chapuis, M.-P.; Estoup, A. Microsatellite null alleles and estimation of population differentiation. Mol. Biol. Evol. 2007, 24, 621–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalakis, Y.M.; Excoffier, L. A generic estimation of population subdivision using distances between alleles with special reference for microsatellite loci. Genetics 1996, 142, 1061–1064. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.O.D.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Takezaki, N.; Nei, M.; Tamura, K. POPTREE2: Software for constructing population trees from allele frequency data and computing other population statistics with Windows interface. Mol. Biol. Evol. 2010, 27, 747–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [Green Version]
- Cavalli-Sforza, L.L.; Edwards, A.W. Phylogenetic analysis. Models and estimation procedures. Am. J. Hum. Genet. 1967, 19, 233. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Vranckx, G.; Jacquemyn, H.; Mergeay, J.; Cox, K.; Kint, V.; Muys, B.; Honnay, O. Transmission of genetic variation from the adult generation to naturally established seedling cohorts in small forest stands of pedunculate oak (Quercus robur L). For. Ecol. Manag. 2014, 312, 19–27. [Google Scholar] [CrossRef]
- Craciunesc, I.; Ciocîrlan, E.; Sofletea, N.; Curtu, A.L. Genetic diversity of pedunculate oak (Quercus robur L.) in Prejmer natural reserve. Bull. Transilv. Univ. Brasov. For. Wood Ind. Agric. Food Eng. Ser. II 2011, 4, 15. [Google Scholar]
- Pohjanmies, T.; Elshibli, S.; Pulkkinen, P.; Rusanen, M.; Vakkari, P.; Korpelainen, H.; Roslin, T. Fragmentation-related patterns of genetic differentiation in pedunculate oak (Quercus robur) at two hierarchical scales. Silva Fenn. 2016, 50, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Katičić Bogdan, I.; Kajba, D.; Šatović, Z.; Schüler, S.; Bogdan, S. Genetic diversity of pedunculate oak (Quercus robur L.) in clonal seed orchards in Croatia, assessed by nuclear and chloroplast microsatellites. SEEFOR 2018, 9, 29–46. [Google Scholar] [CrossRef]
- Morić, M. Genetska Raznolikost Hrasta Lužnjaka (Quercus robur L.) u Pokusnim Nasadima s Potomstvom iz Odabranih Sjemenskih Sastojina. Ph.D. Thesis, University of Zagreb, Zagreb, Croatia, 2016. [Google Scholar]
- Muir, G.; Schloetterer, C. Evidence for shared ancestral polymorphism rather than recurrent gene flow at microsatellite loci differentiating two hybridizing oaks (Quercus spp.). Mol. Ecol. 2005, 14, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Scotti-Saintagne, C.; Bodénès, C.; Barreneche, T.; Bertocchi, E.; Plomion, C.; Kremer, A. Detection of quantitative trait loci controlling bud burst and height growth in Quercus robur L. Theor. Appl. Genet. 2004, 109, 1648–1659. [Google Scholar] [CrossRef]
- Mandák, B.; Bímová, K.; PlačkovÁ, I.; Mahelka, V.; Chrtek, J. Loss of genetic variation in geographically marginal pop-ulations of Atriplex tatarica (Chenopodiaceae). Ann. Bot. 2005, 96, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Leberg, P.L. Effects of population bottlenecks on genetic diversity as measured by allozyme electrophoresis. Evolution 1992, 46, 477–494. [Google Scholar] [CrossRef] [Green Version]
- Greenbaum, G.; Templeton, A.R.; Zarmi, Y.; Bar-David, S. Allelic richness following population founding events–a stochastic modeling framework incorporating gene flow and genetic drift. PLoS ONE 2014, 9, e115203. [Google Scholar] [CrossRef] [Green Version]
- Ballian, D.; Belletti, P.; Ferrazzini, D.; Bogunić, F.; Kajba, D. Genetic variability of pedunculate oak (Quercus robur L.) in Bosnia and Herzegovina. Period. Biol. 2010, 112, 353–362. [Google Scholar]
- Petit, R.J.; El Mousadik, A.; Pons, O. Identifying populations for conservation on the basis of genetic markers. Conserv. Biol. 1998, 12, 844–855. [Google Scholar] [CrossRef]
- Jiménez, P.; Agundez, D.; Alia, R.; Gil, L. Genetic variation in central and marginal populations of Quercus suber L. Silvae Genet. 1999, 48, 278–283. [Google Scholar]
- Vlatković, S. Šume Ravnog Srema, opšte karakteristike i šumovitost područja. In Monografija 250 Godina Šumarstva Ravnog Srema; Tomović, Z., Ed.; Javno preduzeće “Vojvodinašume”–Petrovaradin, Šumsko gazdinstvo: Sremska Mitrovica, Serbia, 2008; pp. 7–18. [Google Scholar]
- Erdeši, J.; Orlović, S.; Galić, Z.; Radosavljević, N. Istorijat šuma Ravnog Srema. In Monografija 250 Godina Šumarstva Ravnog Srema; Tomović, Z., Ed.; Javno preduzeće “Vojvodinašume”–Petrovaradin, Šumsko gazdinstvo: Sremska Mitrovica, Serbia, 2008; pp. 39–44. [Google Scholar]
- Plavšić, S.; Jezdić, D. Šumarstvo Srema u graničarsko doba. In Monografija 250 Godina Šumarstva Ravnog Srema; Tomović, Z., Ed.; Javno preduzeće “Vojvodinašume”–Petrovaradin, Šumsko gazdinstvo: Sremska Mitrovica, Serbia, 2008; pp. 45–54. [Google Scholar]
- Bordács, S.; Horváth, V.; Pintér, B. Development of Resources of Social Broadleaves’ Reproductive Material Production; OTKA Research Reports; Hungarian National Scientific Research Fund: Pécs, Hungary, 2008. [Google Scholar]
- Volner, H. Drvna industrija Slavonije s posebnim osvrtom na obitelj Gutmann do kraja 1918. godine. Hist. Zb. 2012, 65, 453–476. [Google Scholar]
- Martiník, A.; Dobrovolný, L.; Palátová, E. Tree growing space and acorn production of Quercus robur. Dendrobiology 2014, 71, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Kamler, J.; Dobrovolný, L.; Drimaj, J.; Kadavý, J.; Kneifl, M.; Adamec, Z.; Knott, R.; Martiník, A.; Plhal, R.; Zeman, J.; et al. The impact of seed predation and browsing on natural sessile oak regeneration under different light conditions in an over-aged coppice stand. iForest 2016, 9, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Gradečki-Poštenjak, M.; Novak Agbaba, S.; Licht, R.; Posarić, D. Dinamika plodnošenja i kvaliteta uroda sjemena hrasta lužnjaka (Quercus robur L.) u narušenim ekološkim uvjetima. Šumarski List 2011, 135, 169–181. [Google Scholar]
- Koskela, J.; Lefèvre, F.; Schueler, S.; Kraigher, H.; Olrik, D.C.; Hubert, J.; Ditlevsen, B. Translating conservation genetics into management: Pan-European minimum requirements for dynamic conservation units of forest tree genetic diversity. Biol. Conserv. 2013, 157, 39–49. [Google Scholar] [CrossRef] [Green Version]
- de Vries, S.M.; Alan, M.; Bozzano, M.; Burianek, V.; Collin, E.; Cottrell, J.; Ivankovic, M.; Kelleher, T.C.; Koskela, J.; Rotach, P.; et al. Pan-European strategy for genetic conservation of forest trees and establishment of a core network of dynamic conservation units. In European Forest Genetic Resources Programme (EUFORGEN); Bioversity International: Rome, Italy, 2015. [Google Scholar]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Markert, J.A.; Champlin, D.M.; Gutjahr-Gobell, R.; Grear, J.S.; Kuhn, A.; McGreevy, T.J.; Roth, A.; Mark, J.; Bagley, M.; Nacci, D.E. Population genetic diversity and fitness in multiple environments. BMC Evol. Biol. 2010, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Porth, I.; El-Kassaby, Y.A. Assessment of the genetic diversity in forest tree populations using molecular markers. Diversity 2014, 6, 283–295. [Google Scholar] [CrossRef] [Green Version]
- 23 New Sites Added to UNESCO’s World Network of Biosphere Reserves. Available online: https://en.unesco.org/news/23-new-sites-added-unesco-s-world-network-biosphere-reserves (accessed on 14 June 2017).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abb. | Population | Latitude | Longitude | Age of Population | Altitude (m a.s.l.) | Mean Air Temperature (°C) | Annual Sum of Precipitation (mm) |
|---|---|---|---|---|---|---|---|
| BM | Bački Monoštor | 45°91′ | 18°88′ | 115 | 84 | 11.1 | 613 |
| AP | Apatin | 45°62′ | 18°94′ | 117 | 88 | 11.2 | 649 |
| OD | Odzaci | 45°46′ | 19°16′ | 137 | 85 | 11.2 | 635 |
| MO | Morović | 44°92′ | 19°12′ | 131 | 93 | 11.5 | 498 |
| KL | Klenak | 44°79′ | 19°75′ | 97 | 86 | 11.6 | 688 |
| KU | Kupinovo | 44°74′ | 19°99′ | 146 | 83 | 11.6 | 663 |
| KG | Kragujevac | 44°14′ | 21°09′ | 90 | 109 | 11.8 | 686 |
| Locus | Primer Sequences (5′→3′) | Repeated Motif | Fragment Length (bp) | Ta (°C) |
|---|---|---|---|---|
| QrZAG87 | TCCCACCACTTTGGTCTCTCA GTTGTCAGCAGTGGGATGGGTA | (TC)20 | 110–131 | 56 |
| QrZAG30 | TGCTCCGTCATAATCTTGCTCTGA GCAATCCTATCATGCACATGCACAT | (GA)26 | 172–248 | 56 |
| QrZAG101 | CCTGCACAATCAAATCCTTCACTT GCCATGAACAACGGCGGTATCTAG | (TC)20(AC)15 | 136–160 | 50 |
| QrZAG11 | CCTTGAACTCGAAGGTGTCCTT GTAGGTCAAAACCATTGGTTGACT | (TC)22 | 238–267 | 50 |
| QpZAG110 | GGAGGCTTCCTTCAACCTACT GATCTCTTGTGTGCTGTATTT | (AG)15 | 206–262 | 50 |
| QrZAG112 | TTCTTGCTTTGGTGCGCG GTGGTCAGAGACTCGGTAAGTATTC | (GA)32 | 85–96 | 50 |
| QrZAG96 | CCCAGTCACATCCACTACTGTCC GGTTGGGAAAAGGAGATCAGA | (TC)20 | 135–194 | 55 |
| MSQ13 | TGGCTGCACCTATGGCTCTTAGACACTCAGACCCACCATTTTTCC | (TC)N | 222–246 | 50 |
| QrZAG15 | CGATTTGATAATGACACTATGG CATCGACTCATTGTTAAGCAC | (GA)15 | 144–216 | 45 |
| QrZAG20 | CCATTAQAAAGAAGCAGTATTTTGT GCAACACTCAGCCTATATCTAGAA | (TC)20 | 160–200 | 48 |
| QpZAG1/5 | GCTTGAGAGTTGAGATTTGT GCAACACCCTTTAACTACCA | (GT)5(GA)9 | 160–190 | 55 |
| QpZAG9 | GCAATTACAGGCTAGGCTGG GTCTGGACCTAGCCCTCATG | (AG)12 | 182–210 | 65 |
| Locus | Na | Ne | I | Ho | He | Fst | Fis | fn |
|---|---|---|---|---|---|---|---|---|
| QrZAG87 | 24 | 8.439 | 2.333 | 0.862 | 0.874 | 0.025 | 0.021 | 0.004 |
| QrZAG30 | 36 | 12.876 | 2.749 | 0.590 | 0.920 | 0.026 | 0.355 * | 0.163 |
| QrZAG11 | 36 | 9.360 | 2.513 | 0.676 | 0.883 | 0.048 | 0.179 * | 0.087 |
| QrZAG96 | 18 | 1.773 | 0.907 | 0.238 | 0.404 | 0.065 | 0.412 | 0.102 |
| QrZAG112 | 23 | 5.323 | 1.999 | 0.448 | 0.809 | 0.025 | 0.453 | 0.197 |
| QrZAG20 | 18 | 6.320 | 2.016 | 0.667 | 0.840 | 0.017 | 0.213 * | 0.090 |
| QpZAG9 | 15 | 7.359 | 2.116 | 0.671 | 0.862 | 0.023 | 0.146 * | 0.058 |
| MSQ13 | 11 | 3.523 | 1.411 | 0.478 | 0.694 | 0.040 | 0.211 * | 0.079 |
| QrZAG15 | 12 | 3.432 | 1.419 | 0.571 | 0.697 | 0.053 | 0.200 * | 0.073 |
| QpZAG1/5 | 14 | 4.206 | 1.650 | 0.690 | 0.757 | 0.017 | 0.104 | 0.038 |
| QrZAG101 | 20 | 7.440 | 2.227 | 0.652 | 0.862 | 0.031 | 0.025 | 0.080 |
| QpZAG110 | 26 | 2.732 | 1.531 | 0.595 | 0.622 | 0.022 | 0.040 | 0.014 |
| Mean | 29.99 | 11.464 | 1.906 | 0.595 | 0.769 | 0.032 | 0.209 | 0.082 |
| St. Dev. | 0.012 | 0.497 | 0.060 | 0.021 | 0.017 | 0.004 | 0.038 |
| Population | Ne | He | I | AR | pAR | Fst |
|---|---|---|---|---|---|---|
| OD | 5.642 | 0.748 | 1.820 | 8.69 | 0.59 | |
| KL | 5.872 | 0.740 | 1.841 | 9.44 | 0.91 | |
| KU | 6.316 | 0.793 | 1.998 | 10.47 | 1.12 | |
| BM | 6.168 | 0.783 | 1.931 | 9.56 | 0.52 | |
| MO | 5.648 | 0.746 | 1.810 | 8.78 | 0.41 | |
| AP | 6.429 | 0.790 | 1.994 | 10.26 | 0.87 | |
| KG | 6.382 | 0.781 | 1.949 | 10.21 | 1.10 | |
| Mean | 6.065 | 0.769 | 1.906 | 9.63 | 0.79 | 0.032 |
| Source | df | SS | MS | Est. Var. | % | ΦsT | p Value |
|---|---|---|---|---|---|---|---|
| Among Pops | 6 | 123.876 | 20.646 | 0.298 | 2 | 0.025 | 0.001 |
| Within Pops | 203 | 2376.200 | 11.705 | 11.705 | 98 | ||
| Total | 209 | 2500.076 | 12.003 | 100 |
| FSTENA | ||||||||
| DC INA | Population | OD | KL | KU | BM | MO | AP | KG |
| OD | – | 0.008 * | 0.010 * | 0.015 * | 0.004 * | 0.012 * | 0.004 * | |
| KL | 0.308 * | – | 0.015 * | 0.028 * | 0.015 * | 0.012 * | 0.015 * | |
| KU | 0.342 * | 0.346 * | – | 0.012 * | 0.017 * | 0.005 * | 0.0002 * | |
| BM | 0.333 * | 0.349 * | 0.340 * | – | 0.021 * | 0.017 | 0.004 | |
| MO | 0.314 * | 0.291 * | 0.347 * | 0.307 * | – | 0.018 * | 0.016 * | |
| AP | 0.319 * | 0.318 * | 0.317 * | 0.326 * | 0.320 * | – | 0.002 * | |
| KG | 0.307 | 0.329 * | 0.314 * | 0.315 * | 0.343 * | 0.328 * | – | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kesić, L.; Cseke, K.; Orlović, S.; Stojanović, D.B.; Kostić, S.; Benke, A.; Borovics, A.; Stojnić, S.; Avramidou, E.V. Genetic Diversity and Differentiation of Pedunculate Oak (Quercus robur L.) Populations at the Southern Margin of Its Distribution Range—Implications for Conservation. Diversity 2021, 13, 371. https://doi.org/10.3390/d13080371
Kesić L, Cseke K, Orlović S, Stojanović DB, Kostić S, Benke A, Borovics A, Stojnić S, Avramidou EV. Genetic Diversity and Differentiation of Pedunculate Oak (Quercus robur L.) Populations at the Southern Margin of Its Distribution Range—Implications for Conservation. Diversity. 2021; 13(8):371. https://doi.org/10.3390/d13080371
Chicago/Turabian StyleKesić, Lazar, Klára Cseke, Saša Orlović, Dejan B. Stojanović, Saša Kostić, Attila Benke, Attila Borovics, Srđan Stojnić, and Evangelia V. Avramidou. 2021. "Genetic Diversity and Differentiation of Pedunculate Oak (Quercus robur L.) Populations at the Southern Margin of Its Distribution Range—Implications for Conservation" Diversity 13, no. 8: 371. https://doi.org/10.3390/d13080371
APA StyleKesić, L., Cseke, K., Orlović, S., Stojanović, D. B., Kostić, S., Benke, A., Borovics, A., Stojnić, S., & Avramidou, E. V. (2021). Genetic Diversity and Differentiation of Pedunculate Oak (Quercus robur L.) Populations at the Southern Margin of Its Distribution Range—Implications for Conservation. Diversity, 13(8), 371. https://doi.org/10.3390/d13080371