
Ain’t Nothing like Family—Female Brown Bears Share Their Home Range with Relatives

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Spatial and Genetic Data Collection
2.3. Laboratory Methods
2.4. Genetic Analyses
2.5. Spatial Analysis
3. Results
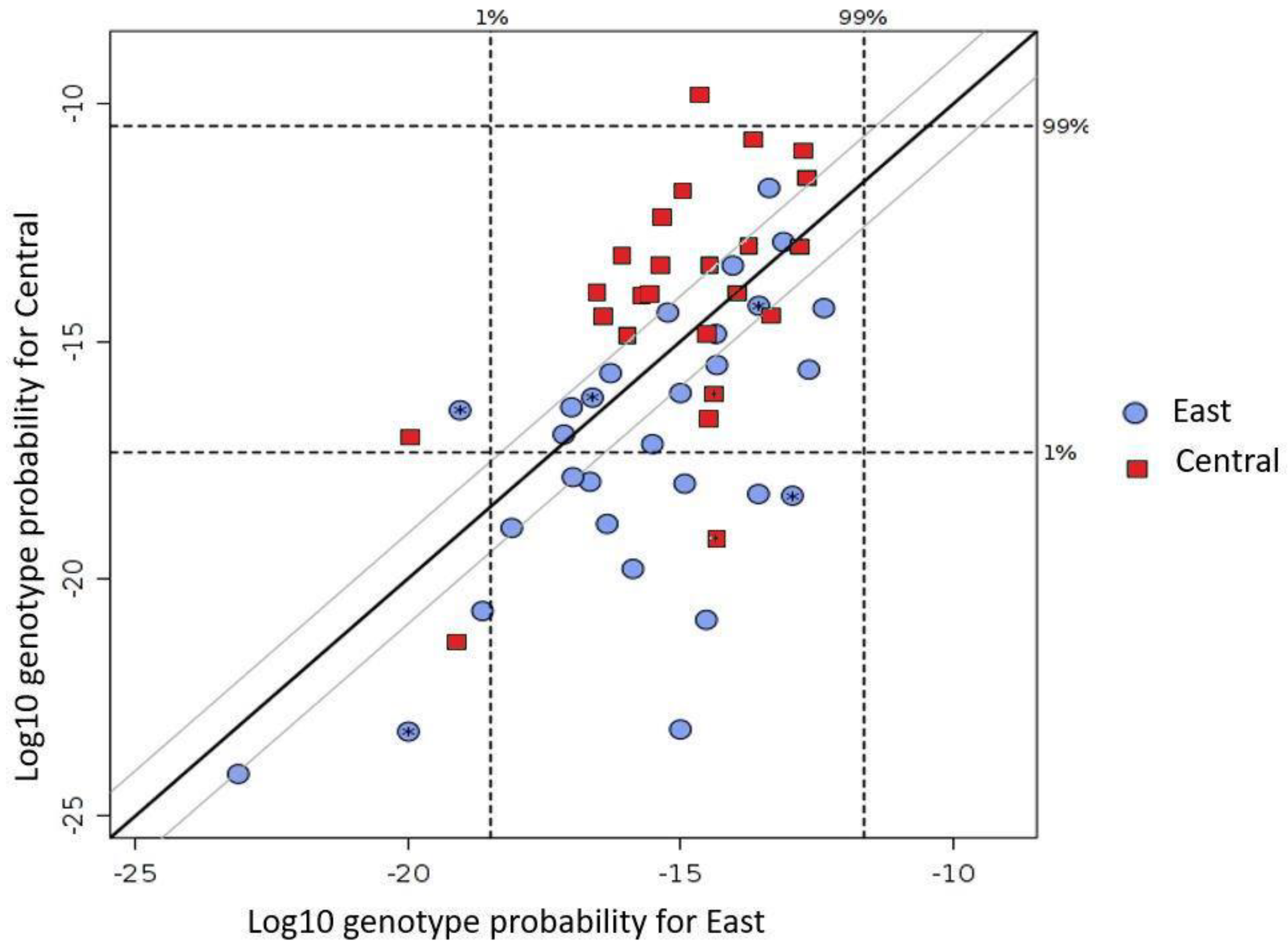
3.1. Genetic Diversity
3.2. Relatedness and Genetic Structure
3.3. Patterns of Space Use and Their Link to Relatedness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whitehead, H. Analysing animal social structure. Anim. Behav. 1997, 53, 1053–1067. [Google Scholar] [CrossRef] [Green Version]
- Keiper, R.R. Social Interactions of the Przewalski Horse (Equus Przewalskii Poliakov, 1881) Herd at the Munich Zoo. Appl. Anim. Behav. Sci. 1988, 21, 89–97. [Google Scholar] [CrossRef]
- Peterson, R.O.; Jacobs, A.K.; Drummer, T.D.; Mech, L.D.; Smith, D.W. Leadership Behavior in Relation to Dominance and Reproductive Status in Gray Wolves, Canis lupus. Can. J. Zool. 2002, 80, 1405–1412. [Google Scholar] [CrossRef] [Green Version]
- Gero, S.; Gordon, J.; Whitehead, H. Individualized Social Preferences and Long-Term Social Fidelity between Social Units of Sperm Whales. Anim. Behav. 2015, 102, 15–23. [Google Scholar] [CrossRef]
- Sandell, M. The Mating Tactics and Spacing Patterns of Solitary Carnivores. In Carnivore Behavior, Ecology, and Evolution; Gittleman, J.L., Ed.; Springer: Boston, MA, USA, 1989; pp. 164–182. ISBN 978-1-4757-4716-4. [Google Scholar]
- Elbroch, L.M.; Lendrum, P.E.; Quigley, H.; Caragiulo, A. Spatial Overlap in a Solitary Carnivore: Support for the Land Tenure, Kinship or Resource Dispersion Hypotheses? J. Anim. Ecol. 2016, 85, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, D.W. The Ecology of Carnivore Social Behaviour. Nature 1983, 301, 379–384. [Google Scholar] [CrossRef]
- Johnson, D.D.P.; Kays, R.; Blackwell, P.G.; Macdonald, D.W. Does the resource dispersion hypothesis explain group living? Trends Ecol. Evol. 2002, 17, 563–570. [Google Scholar] [CrossRef]
- Wagner, A.; Frank, L.; Creel, S. Spatial grouping in behaviourally solitary striped hyaenas, Hyaena hyaena. Anim. Behav. 2008, 75, 1131–1142. [Google Scholar] [CrossRef]
- Macdonald, D.W.; Johnson, D.D.P. Patchwork Planet: The Resource Dispersion Hypothesis, Society, and the Ecology of Life. J. Zool. 2015, 295, 75–107. [Google Scholar] [CrossRef]
- Seidensticker, J.C.; Hornocker, M.G.; Wiles, W.V.; Messick, J.P. Mountain Lion Social Organization in the Idaho Primitive Area. Wildl. Monogr. 1973, 35, 3–60. Available online: http://www.jstor.org/stable/3830509 (accessed on 22 November 2021).
- Guilder, J.; Barca, B.; Arroyo-Arce, S.; Gramajo, R.; Salom, R. Jaguars (Panthera onca) Increase Kill Utilization Rates and Share Prey in Response to Seasonal Fluctuations in Nesting Green Turtle (Chelonia mydas mydas) Abundance in Tortuguero National Park, Costa Rica. Mamm. Biol. 2015, 80, 65–72. [Google Scholar] [CrossRef]
- Elbroch, L.M.; Quigley, H. Social Interactions in a Solitary Carnivore. Curr. Zool. 2017, 63, 357–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreras, P.; Beltrán, J.F.; Aldama, J.J.; Delibes, M. Spatial Organization and Land Tenure System of the Endangered Iberian Lynx (Lynx pardinus). J. Zool. 1997, 243, 163–189. [Google Scholar] [CrossRef]
- Diefenbach, D.R.; Hansen, L.A.; Warren, R.J.; Conroy, M.J. Spatial Organization of a Reintroduced Population of Bobcats. J. Mammal. 2006, 87, 394–401. [Google Scholar] [CrossRef] [Green Version]
- López-Bao, J.V.; Rodríguez, A.; Alés, E. Field Observation of Two Males Following a Female in the Iberian Lynx (Lynx pardinus) during the Mating Season. Mamm. Biol. 2008, 73, 404–406. [Google Scholar] [CrossRef]
- Griffiths, S.W.; Armstrong, J.D. The Benefits of Genetic Diversity Outweigh Those of Kin Association in a Territorial Animal. Proc. Biol. Sci. 2001, 268, 1293–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, G.E.; Brown, J.A. Do Kin Always Make Better Neighbours?: The Effects of Territory Quality. Behav. Ecol. Sociobiol. 1993, 33, 225–231. [Google Scholar] [CrossRef]
- Schenk, A.; Obbard, M.E.; Kovacs, K.M. Genetic Relatedness and Home-Range Overlap among Female Black Bears (Ursus americanus) in Northern Ontario, Canada. Can. J. Zool. 1998, 76, 1511–1519. [Google Scholar] [CrossRef]
- Griffiths, S.; Armstrong, J. Kin-Biased Territory Overlap and Food Sharing among Juvenile Atlantic Salmon. J. Anim. Ecol. 2002, 71, 480–486. [Google Scholar] [CrossRef]
- Støen, O.-G.; Bellemain, E.; Sæbø, S.; Swenson, J.E. Kin-Related Spatial Structure in Brown Bears Ursus arctos. Behav. Ecol. Sociobiol. 2005, 59, 191–197. [Google Scholar] [CrossRef]
- Ordiz, A.; Støen, O.-G.; Swenson, J.E.; Kojola, I.; Bischof, R. Distance-Dependent Effect of the Nearest Neighbor: Spatiotemporal Patterns in Brown Bear Reproduction. Ecology 2008, 89, 3327–3335. [Google Scholar] [CrossRef] [PubMed]
- Innes, R.J.; McEachern, M.B.; Van Vuren, D.H.; Eadie, J.M.; Kelt, D.A.; Johnson, M.L. Genetic Relatedness and Spatial Associations of Dusky-Footed Woodrats (Neotoma fuscipes). J. Mammal. 2012, 93, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Rode, K.D.; Farley, S.D.; Robbins, C.T. Sexual dimorphism, reproductive strategy, and human activities determine resource use by brown bears. Ecology 2006, 87, 2636–2646. [Google Scholar] [CrossRef]
- Dahle, B.; Swenson, J.E. Family Breakup in Brown Bears: Are Young Forced to Leave? J. Mammal. 2003, 84, 5. [Google Scholar] [CrossRef] [Green Version]
- Steyaert, S.M.J.G.; Endrestøl, A.; Hackländer, K.; Swenson, J.E.; Zedrosser, A. The Mating System of the Brown Bear Ursus Arctos: Brown Bear Mating System. Mammal. Rev. 2012, 42, 12–34. [Google Scholar] [CrossRef]
- Craighead, L.; Paetkau, D.; Reynolds, H.V.; Vyse, E.R.; Strobeck, C. Microsatellite Analysis of Paternity and Reproduction in Arctic Grizzly Bears. J. Hered. 1995, 86, 255–261. [Google Scholar] [CrossRef]
- Bellemain, E.; Swenson, J.E.; Taberlet, P. Mating Strategies in Relation to Sexually Selected Infanticide in a Non-Social Carnivore: The Brown Bear. Ethology 2006, 112, 238–246. [Google Scholar] [CrossRef]
- McLellan, B.N.; Hovey, F.W. Natal Dispersal of Grizzly Bears. Can. J. Zool. 2001, 79, 838–844. [Google Scholar] [CrossRef]
- Schwartz, C.C.; Miller, S.D.; Haroldson, M.A. Grizzly bear Ursus arctos. In Wild Mammals of North America: Biology, Management, and Conservation, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2003; pp. 556–586. Available online: http://pubs.er.usgs.gov/publication/70169862 (accessed on 22 November 2021).
- Bellemain, E.; Zedrosser, A.; Manel, S.; Waits, L.P.; Taberlet, P.; Swenson, J.E. The Dilemma of Female Mate Selection in the Brown Bear, a Species with Sexually Selected Infanticide. Proc. Biol. Sci. 2006, 273, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glenn, L.P.; Miller, L.H. Seasonal movements of an Alaska Peninsula brown bear population. In Bears: Their Biology and Management: A Selection of Papers from the 4th International Conference on Bear Research and Management; Martinka, C.J., McArthur, K.L., Eds.; International Association for Bear Research and Management: Kalispell, Montana, 1980; pp. 307–312. [Google Scholar]
- Dahle, B.; Swenson, J.E. Home Ranges in Adult Scandinavian Brown Bears (Ursus arctos): Effect of Mass, Sex, Reproductive Category, Population Density and Habitat Type. J. Zool. 2003, 260, 329–335. [Google Scholar] [CrossRef]
- Dahle, B.; Swenson, J.E. Seasonal Range Size in Relation to Reproductive Strategies in Brown Bears Ursus arctos. J. Anim. Ecol. 2003, 72, 660–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Walle, J.; Pigeon, G.; Zedrosser, A.; Swenson, J.E.; Pelletier, F. Hunting regulation favors slow life histories in a large carnivore. Nat. Commun. 2018, 9, 1100. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L. Effects of Food Supply and Kinship on Social Behavior Movements and Population Growth of Black Bears in Northeastern Minnesota USA. Wildl. Monogr. 1987, 97, 1–72. [Google Scholar]
- Støen, O.-G.; Zedrosser, A.; Sæbø, S.; Swenson, J.E. Inversely Density-Dependent Natal Dispersal in Brown Bears Ursus arctos. Oecologia 2006, 148, 356. [Google Scholar] [CrossRef]
- Kojola, L.; Danilov, P.I.; Laitala, H.-M.; Belkin, V.A.; Yakimov, A. Brown Bear Population Structure in Core and Periphery: Analysis of Hunting Statistics from Russian Karelia and Finland. Ursus 2007, 14, 17–20. Available online: https://www.jstor.org/stable/3872952 (accessed on 22 November 2021).
- Zedrosser, A.; Dahle, B.; Swenson, J.E.; Gerstl, N. Status and Management of the Brown Bear in Europe. Ursus 2001, 12, 9–20. Available online: https://www.jstor.org/stable/3873224 (accessed on 22 November 2021).
- Pulliainen, E. Brown Bear Immigration into Finland from the East. Bears Biol. Manag. 1986, 6, 15–20. [Google Scholar] [CrossRef]
- Kopatz, A.; Eiken, H.G.; Hagen, S.B.; Ruokonen, M.; Esparza-Salas, R.; Schregel, J.; Kojola, I.; Smith, M.E.; Wartiainen, I.; Aspholm, P.E.; et al. Connectivity and Population Subdivision at the Fringe of a Large Brown Bear (Ursus arctos) Population in North Western Europe. Conserv. Genet. 2012, 13, 681–692. [Google Scholar] [CrossRef]
- Danilov, P.I.; Tirronen, K.F.; Belkin, V.V.; Panchenko, D.V.; Fedorov, F.V. Brown Bear and an Estimate of Its Abundance in the European Taiga; PetroPress: Petrozavodsk, Russia, 2014; p. 33. ISBN 978-5-8430-0117-9. [Google Scholar]
- Kolesnikov, V.V.; Dvornikov, M.G.; Zarubin, B.E.; Makarov, V.A.; Makarova, D.S.; Piminov, V.N.; Pankratov, A.P.; Sinitsyn, A.A.; Skumatov, D.V.; Soloviev, V.A.; et al. Scientifically Grounded Proposals for the State System for Monitoring the Resources of the Main Species of Hunting Animals in the Russian Federation; FGBNU VNIIOZ: Kirov, Russia, 2017; p. 37. Available online: http://vniioz-kirov.ru/upload/iblock/f24/f24282455681542b9fe9ff519eabecfd.pdf (accessed on 9 October 2021).
- Heikkinen, S. Karhukanta Suomessa 2020. In Luonnonvara- ja Biotalouden Tutkimus; Luonnonvarakeskus: Helsinki, Finland, 2020; ISBN 978-952-380-177-6. [Google Scholar]
- Kopatz, A.; Kleven, O.; Kojola, I.; Aspi, J.; Norman, A.J.; Spong, G.; Gyllenstrand, N.; Dalén, L.; Fløystad, I.; Hagen, S.B.; et al. Restoration of Transborder Connectivity for Fennoscandian Brown Bears (Ursus arctos). Biol. Conserv. 2021, 253, 108936. [Google Scholar] [CrossRef]
- Kopatz, A.; Eiken, H.G.; Aspi, J.; Kojola, I.; Tobiassen, C.; Tirronen, K.F.; Danilov, P.I.; Hagen, S.B. Admixture and Gene Flow from Russia in the Recovering Northern European Brown Bear (Ursus arctos). PLoS ONE 2014, 9, e97558. [Google Scholar] [CrossRef] [Green Version]
- National Land Survey of Finland. Suomen Pinta-ala Kunnittain 1.1.2021. Available online: https://www.maanmittauslaitos.fi/sites/maanmittauslaitos.fi/files/attachments/2021/02/Vuoden_2021_pinta-alatilasto_kunnat_maakunnat.pdf (accessed on 15 April 2021).
- Statistics Finland, Population. Available online: https://www.tilastokeskus.fi/tup/suoluk/suoluk_vaesto_en.html (accessed on 15 April 2021).
- Penteriani, V.; Lamamy, C.; Kojola, I.; Heikkinen, S.; Bombieri, G.; del Mar Delgado, M. Does Artificial Feeding Affect Large Carnivore Behaviours? The Case Study of Brown Bears in a Hunted and Tourist Exploited Subpopulation. Biol. Conserv. 2021, 254, 108949. [Google Scholar] [CrossRef]
- Paetkau, D.; Strobeck, C. Microsatellite Analysis of Genetic Variation in Black Bear Populations. Mol. Ecol. 1994, 3, 489–495. [Google Scholar] [CrossRef]
- Paetkau, D.; Calvert, W.; Stirling, I.; Strobeck, C. Microsatellite Analysis of Population Structure in Canadian Polar Bears. Mol. Ecol. 1995, 4, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Camarra, J.-J.; Griffin, S.; Uhrès, E.; Hanotte, O.; Waits, L.P.; Dubois-Paganon, C.; Burke, T.; Bouvet, J. Noninvasive Genetic Tracking of the Endangered Pyrenean Brown Bear Population. Mol. Ecol. 1997, 6, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. Genalex 6: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 22 November 2021).
- Jombart, T. Adegenet: A R Package for the Multivariate Analysis of Genetic Markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Ahmed, I. Adegenet 1.3-1: New Tools for the Analysis of Genome-Wide SNP Data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J. FSTAT (Version 1.2): A Computer Program to Calculate F-Statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. Genepop’007: A Complete Re-Implementation of the Genepop Software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Pew, J.; Muir, P.H.; Wang, J.; Frasier, T.R. Related: An R Package for Analysing Pairwise Relatedness from Codominant Molecular Markers. Mol. Ecol. Resour. 2015, 15, 557–561. [Google Scholar] [CrossRef]
- Wang, J. An Estimator for Pairwise Relatedness Using Molecular Markers. Genetics 2002, 160, 1203–1215. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, S.T.; Wagner, A.P.; Taper, M.L. ML-Relate: A computer program for maximum likelihood estimation of relatedness and relationship. Mol. Ecol. Notes. 2006, 6, 576–579. [Google Scholar] [CrossRef]
- McMillan, L.F.; Fewster, R.M. Visualizations for Genetic Assignment Analyses Using the Saddlepoint Approximation Method. Biometrics 2017, 73, 1029–1041. [Google Scholar] [CrossRef]
- Calenge, C. The Package “Adehabitat” for the R Software: A Tool for the Analysis of Space and Habitat Use by Animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Bauder, J.M.; Breininger, D.R.; Bolt, M.R.; Legare, M.L.; Jenkins, C.L.; McGarigal, K. The Role of the Bandwidth Matrix in Influencing Kernel Home Range Estimates for Snakes Using VHF Telemetry Data. Wildl. Res. 2015, 42, 437–453. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.-B. The Ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Fieberg, J.; Kochanny, C.O. Quantifying Home-Range Overlap: The Importance of the Utilization Distribution. J. Wildl. Manag. 2005, 69, 1346–1359. [Google Scholar] [CrossRef]
- Savje, F. Distances: Tools for Distance Metrics Version 0.1.8 from CRAN. Available online: https://rdrr.io/cran/distances/ (accessed on 5 July 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Goslee, S.C.; Urban, D.L. The Ecodist Package for Dissimilarity-Based Analysis of Ecological Data. J. Stat. Softw. 2007, 22, 1–19. [Google Scholar] [CrossRef]
- Franckowiak, R.P.; Panasci, M.; Jarvis, K.J.; Acuña-Rodriguez, I.S.; Landguth, E.L.; Fortin, M.-J.; Wagner, H.H. Model Selection with Multiple Regression on Distance Matrices Leads to Incorrect Inferences. PLoS ONE 2017, 12, e0175194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojola, I.; Hallikainen, V.; Heikkinen, S.; Nivala, V. Inadvertent Shooting of Brown Bear Cubs in Finland: What Can Managers Do to Reduce It? Wildl. Biol. 2021, 2021, wlb.00773. [Google Scholar] [CrossRef]
- Frank, S.C.; Pelletier, F.; Kopatz, A.; Bourret, A.; Garant, D.; Swenson, J.E.; Eiken, H.G.; Hagen, S.B.; Zedrosser, A. Harvest is associated with the disruption of social and fine-scale genetic structure among matrilines of a solitary large carnivore. Evol Appl. 2021, 14, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Dahle, B.; Støen, O.-G.; Swenson, J.E. Factors Influencing Home-Range Size in Subadult Brown Bears. J. Mammal. 2006, 87, 859–865. [Google Scholar] [CrossRef]
- Vincent, J.P.; Bideau, E.; Hewison, A.J.M.; Angibault, J.M. The influence of increasing body weight, kid production, home range and winter grouping in roe deer. J. Zool. 1995, 236, 371–382. [Google Scholar] [CrossRef]
- Lopez, R.R.; Harveson, P.M.; Peterson, M.N.; Silvy, N.H.; Frank, P.A. From the Field: Changes in ranges of Florida Key deer—does population density matter? Wildl. Soc. 2005, 33, 343–348. Available online: https://www.jstor.org/stable/3784875 (accessed on 22 November 2021). [CrossRef]
- Kiefer, S.J.; Weckerly, F.W. Population density and body weight influences on home range size of feral hogs. Calif. Fish Game 2005, 91, 136–142. [Google Scholar]
- Mcloughlin, P.D.; Ferguson, S.H.; Messier, F. Intraspecific Variation in Home Range Overlap with Habitat Quality: A Comparison among Brown Bear Populations. Evol. Ecol. 2000, 14, 39–60. [Google Scholar] [CrossRef]
- Ramos-Fernández, G.; Boyer, D.; Gómez, V.P. A Complex Social Structure with Fission–Fusion Properties Can Emerge from a Simple Foraging Model. Behav. Ecol. Sociobiol. 2006, 60, 536–549. [Google Scholar] [CrossRef] [Green Version]
- Collins, G.H.; Kovach, S.D.; Hinkes, M.T. Home range and movements of female brown bears in southwestern Alaska. Ursus 2005, 16, 181–189. Available online: http://www.jstor.org/stable/3873029 (accessed on 22 November 2021). [CrossRef]
- Blanchard, B.M.; Knight, R.R. Movements of yellowstone grizzly bears. Biol. Conserv. 1991, 58, 41–67. [Google Scholar] [CrossRef]
- Sato, Y.; Kobayashi, Y.; Urata, T.; Takatsuki, S. Home range and habitat use of female brown bear (Ursus arctos) in Urahoro, eastern Hokkaido, Japan. Mamm. Study 2008, 33, 99–109. [Google Scholar] [CrossRef]
- Huber, D.; Roth, H.U. Movements of European brown bears in Croatia. Acta Theriol. 1993, 38, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Kaczensky, P.; Knauer, F.; Krze, B.; Jonozovic, M.; Adamic, M.; Gossow, H. The impact of high speed, high volume traffic axes on brown bears in Slovenia. Biol. Conserv. 2003, 111, 191–204. [Google Scholar] [CrossRef]
- Penteriani, V.; Delgado, M.M.; López-Bao, J.V.; García, P.V.; Monrós, J.S.; Álvarez, E.V.; Corominas, T.S.; Vázquez, V.M. Patterns of movement of released female brown bears in the Cantabrian Mountains, northwestern Spain. Ursus 2017, 28, 165–170. Available online: https://www.jstor.org/stable/44751706 (accessed on 22 November 2021). [CrossRef]
- Nyholm, E.S. Petosiirrot—Riistantutkimuksen vaatimaton kokeilu. (Translocation of large carnivores). Metsästäjä 1995, 44, 12–14. [Google Scholar]
- Kojola, I.; Heikkinen, S. The Structure of the Expanded Brown Bear Population at the Edge of the Finnish Range. Ann. Zool. Fenn. 2006, 43, 258–262. Available online: https://www.jstor.org/stable/23736871 (accessed on 22 November 2021).
- Waits, L.; Taberlet, P.; Swenson, J.E.; Sandegren, F.; Franzén, R. Nuclear DNA Microsatellite Analysis of Genetic Diversity and Gene Flow in the Scandinavian Brown Bear (Ursus arctos). Mol. Ecol. 2000, 9, 421–431. [Google Scholar] [CrossRef]
- Zachos, F.E.; Otto, M.; Unici, R.; Lorenzini, R.; Hartl, G.B. Evidence of a Phylogeographic Break in the Romanian Brown Bear (Ursus arctos) Population from the Carpathians. Mammal. Biol. 2008, 73, 93–101. [Google Scholar] [CrossRef]
- Straka, M.; Paule, L.; Ionescu, O.; Štofík, J.; Adamec, M. Microsatellite Diversity and Structure of Carpathian Brown Bears (Ursus arctos): Consequences of Human Caused Fragmentation. Conserv. Genet. 2012, 13, 153–164. [Google Scholar] [CrossRef]
- Hagen, S.B.; Kopatz, A.; Aspi, J.; Kojola, I.; Eiken, H.G. Evidence of Rapid Change in Genetic Structure and Diversity during Range Expansion in a Recovering Large Terrestrial Carnivore. Proc. R. Soc. B 2015, 282, 20150092. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Central | East | |||||||
|---|---|---|---|---|---|---|---|---|
| Locus | AR | HO | HE | FIS | AR | HO | HE | FIS |
| MU05 | 6.00 | 0.79 | 0.84 | −0.04 | 6.98 | 0.79 | 0.83 | 0.11 |
| MU09 | 6.00 | 0.79 | 0.86 | −0.11 | 9.92 | 0.79 | 0.82 | 0.16 |
| MU15 | 10.87 | 0.75 | 0.80 | 0.21 | 11.03 | 0.72 | 0.89 | 0.14 |
| G1D | 6.00 | 0.88 | 0.83 | −0.10 | 8.70 | 0.96 | 0.83 | −0.12 |
| G10B | 6.00 | 0.78 | 0.82 | 0.05 | 10.26 | 0.83 | 0.77 | −0.06 |
| MU10 | 5.00 | 0.88 | 0.77 | −0.18 | 6.75 | 0.93 | 0.79 | −0.14 |
| MU50 | 5.00 | 0.50 | 0.61 | −0.17 | 10.04 | 0.63 | 0.52 | 0.10 |
| MU51 | 9.00 | 0.75 | 0.80 | 0.34 | 11.19 | 0.57 | 0.87 | 0.20 |
| MU59 | 15.00 | 0.96 | 0.90 | −0.01 | 21.66 | 0.86 | 0.94 | 0.10 |
| G1A | 14.87 | 0.75 | 0.79 | 0.10 | 15.81 | 0.90 | 0.92 | 0.06 |
| G10L | 5.96 | 0.71 | 0.79 | −0.08 | 7.66 | 0.76 | 0.72 | 0.07 |
| Mean | 8.15 | 0.78 | 0.80 | 0.03 | 10.91 | 0.80 | 0.81 | 0.02 |
| Term | Estimate SE | p | |
|---|---|---|---|
| (A) Home range overlap | (Intercept) | 0.149 ± 0.014 | <2.00e−16 *** |
| Pairwise relatedness | 0.174 ± 0.041 | 6.36e−05 *** | |
| Region (eastern Finland) | −0.003 ± 0.045 | 0.952 | |
| (B) Centroid distance | (Intercept) | 3.913 ± 0.077 | <2.00e−16 *** |
| Pairwise relatedness | −1.029 ± 0.290 | 5.19e−04 *** | |
| Region (eastern Finland) | −0.149 ± 0.123 | 0.23 |
| Population | n | Home Range (km2) | Reference |
|---|---|---|---|
| North America | 35 | 2577 | [80] |
| North America (Alaska) | 20 | 356 | [82] |
| NorthAmerica (Yellowstone) | 21 | 884 | [83] |
| Japan (Hokkaido) | 3 | 43 | [84] |
| Croatia | 5 | 58 | [85] |
| Slovenia | 5 | 53 | [86] |
| Spain | 1 | 28 | [87] |
| Scandinavia | 37 | 437 | [21] |
| Scandinavia (South) | 34 | 217 | [34] |
| Scandinavia (North) | 18 | 280 | [34] |
| Finland (Eastern) | 25 | 127 | This study |
| Finland (Central) | 56 | 862 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olejarz, A.; Aspi, J.; Kojola, I.; Nivala, V.; Niskanen, A.K.; Harmoinen, J. Ain’t Nothing like Family—Female Brown Bears Share Their Home Range with Relatives. Diversity 2022, 14, 41. https://doi.org/10.3390/d14010041
Olejarz A, Aspi J, Kojola I, Nivala V, Niskanen AK, Harmoinen J. Ain’t Nothing like Family—Female Brown Bears Share Their Home Range with Relatives. Diversity. 2022; 14(1):41. https://doi.org/10.3390/d14010041
Chicago/Turabian StyleOlejarz, Astrid, Jouni Aspi, Ilpo Kojola, Vesa Nivala, Alina K. Niskanen, and Jenni Harmoinen. 2022. "Ain’t Nothing like Family—Female Brown Bears Share Their Home Range with Relatives" Diversity 14, no. 1: 41. https://doi.org/10.3390/d14010041