
Description of a New Species of the Genus Cryptomonas (Cryptophyceae: Cryptomonadales), Isolated from Soils in a Tropical Forest


Abstract
1. Introduction
2. Materials and Methods
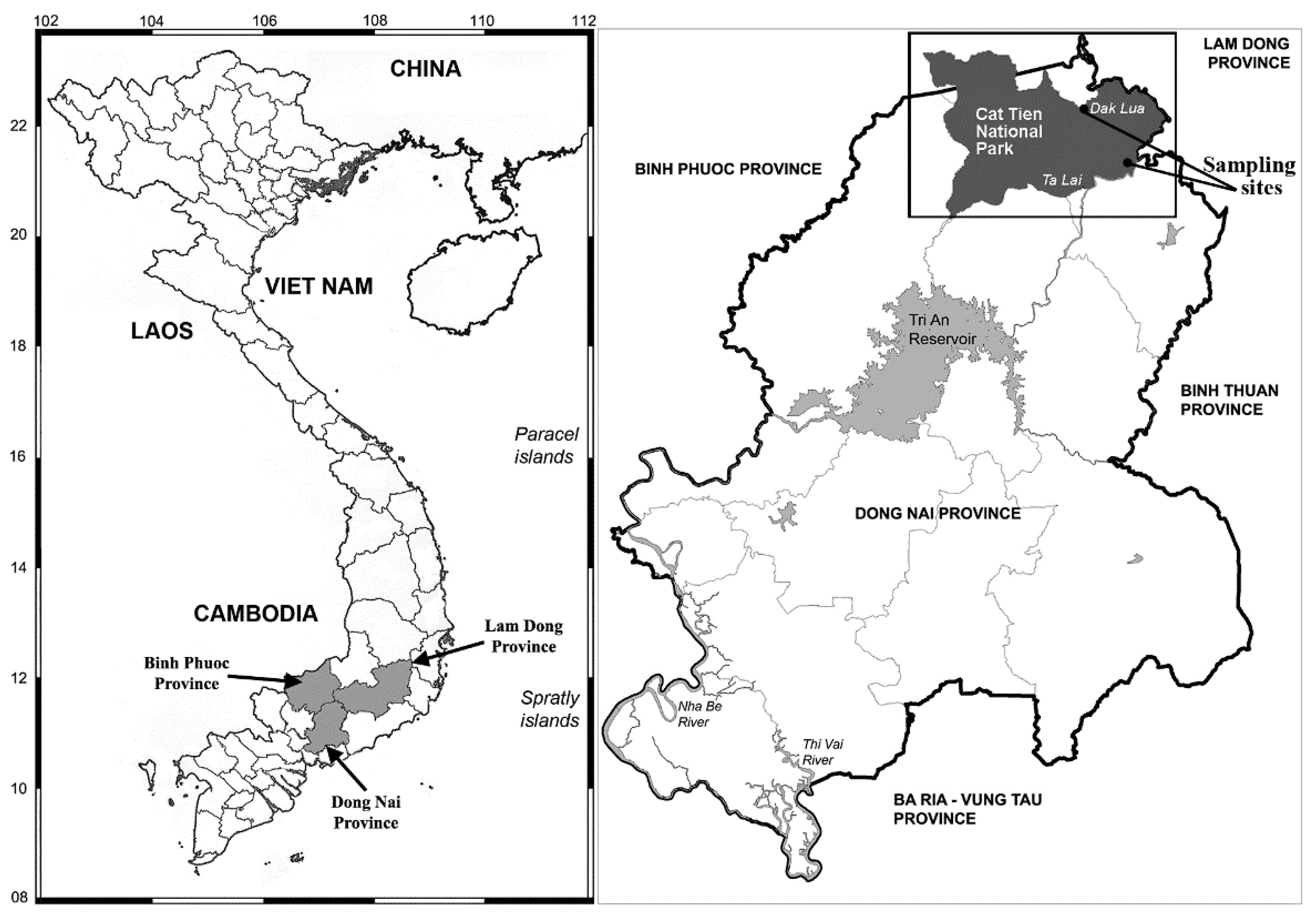
2.1. Study Area
2.2. Samples and Collections
2.3. Culturing
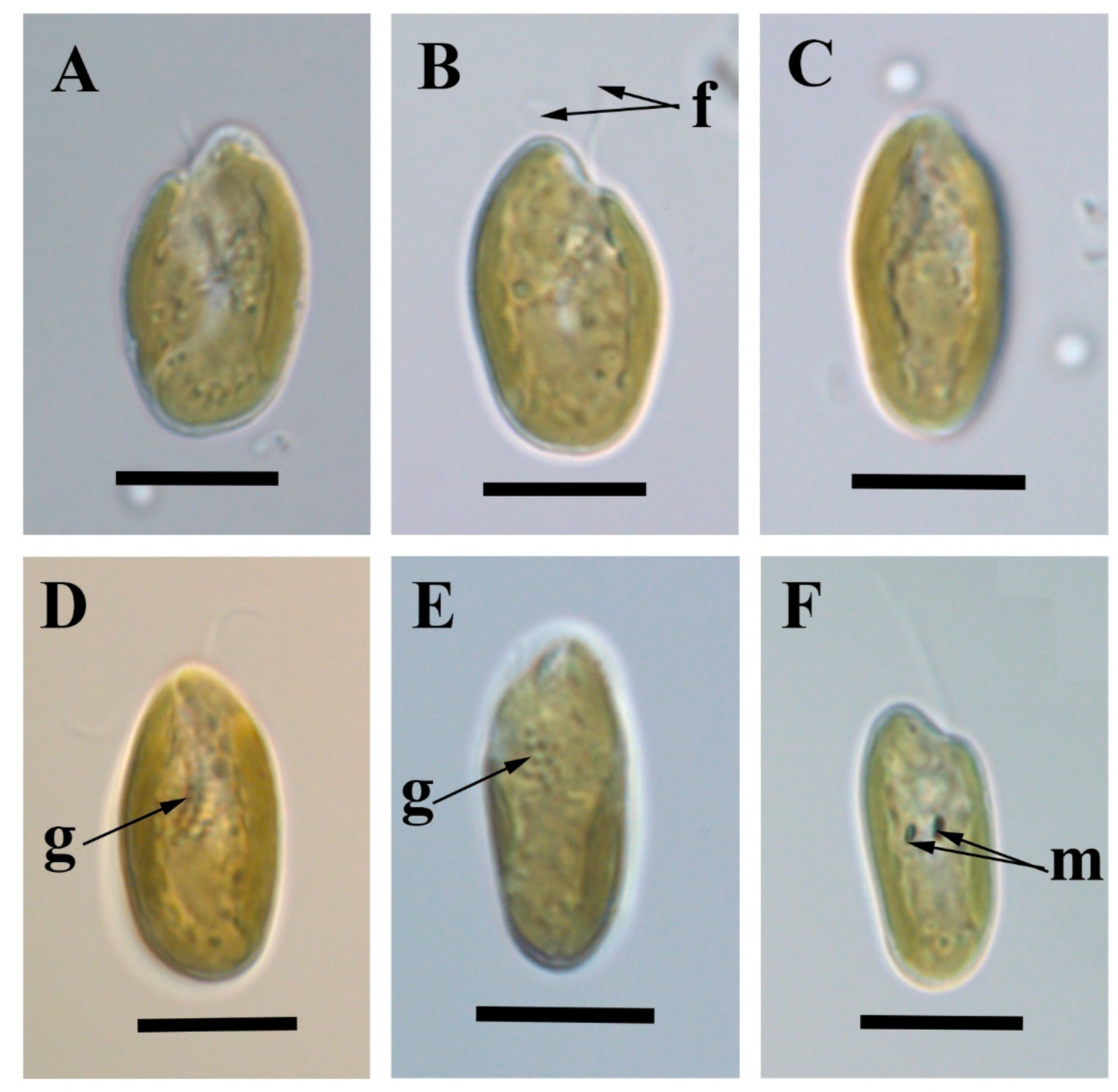
2.4. Light Microscopy
2.5. Amplification and Sequencing
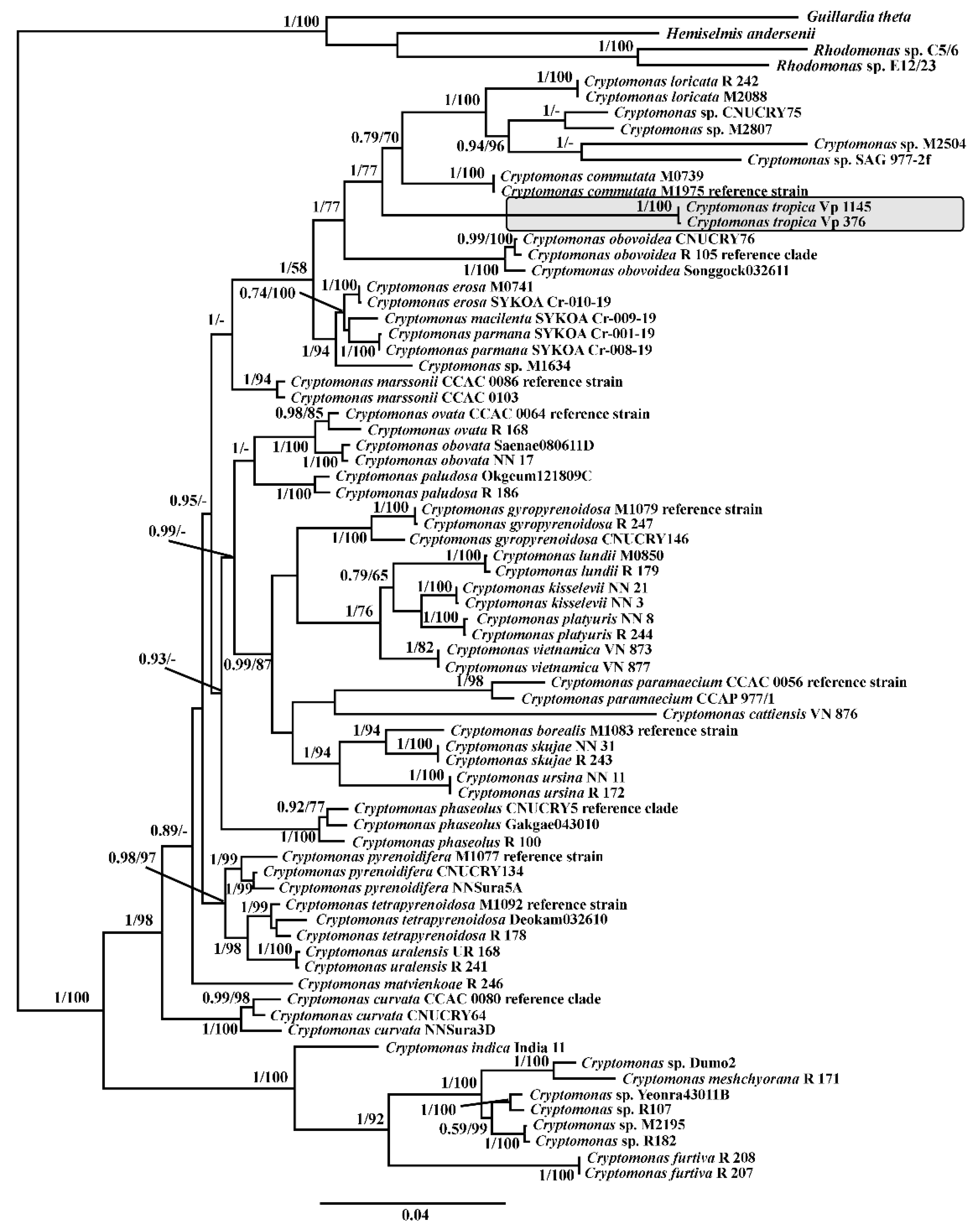
2.6. Alignment and Phylogenetic Analysis of SSU, LSU rDNA and psbA cpDNA
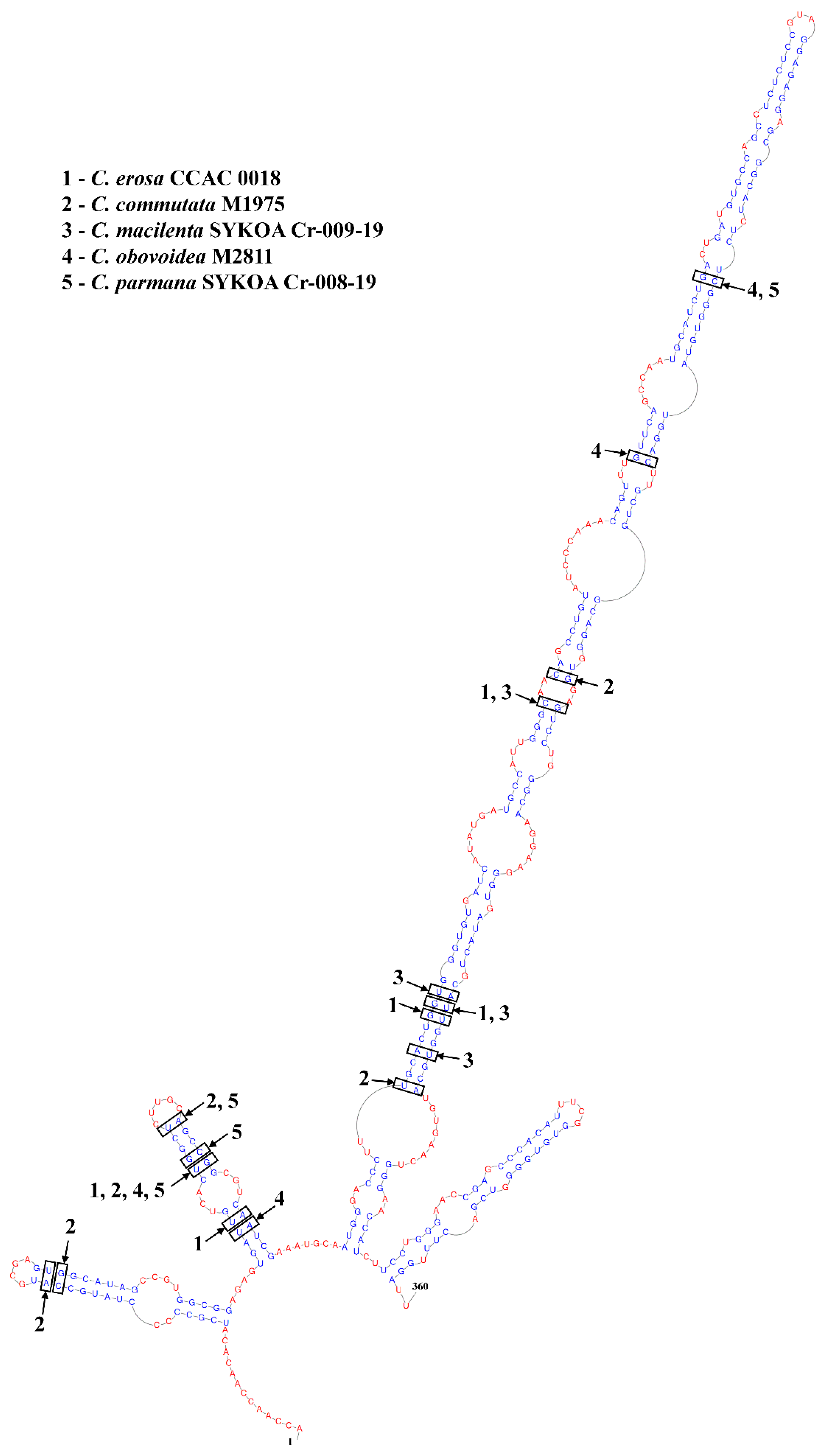
2.7. ITS2 Annotation, Secondary Structure Modeling, Alignment and Phylogeny
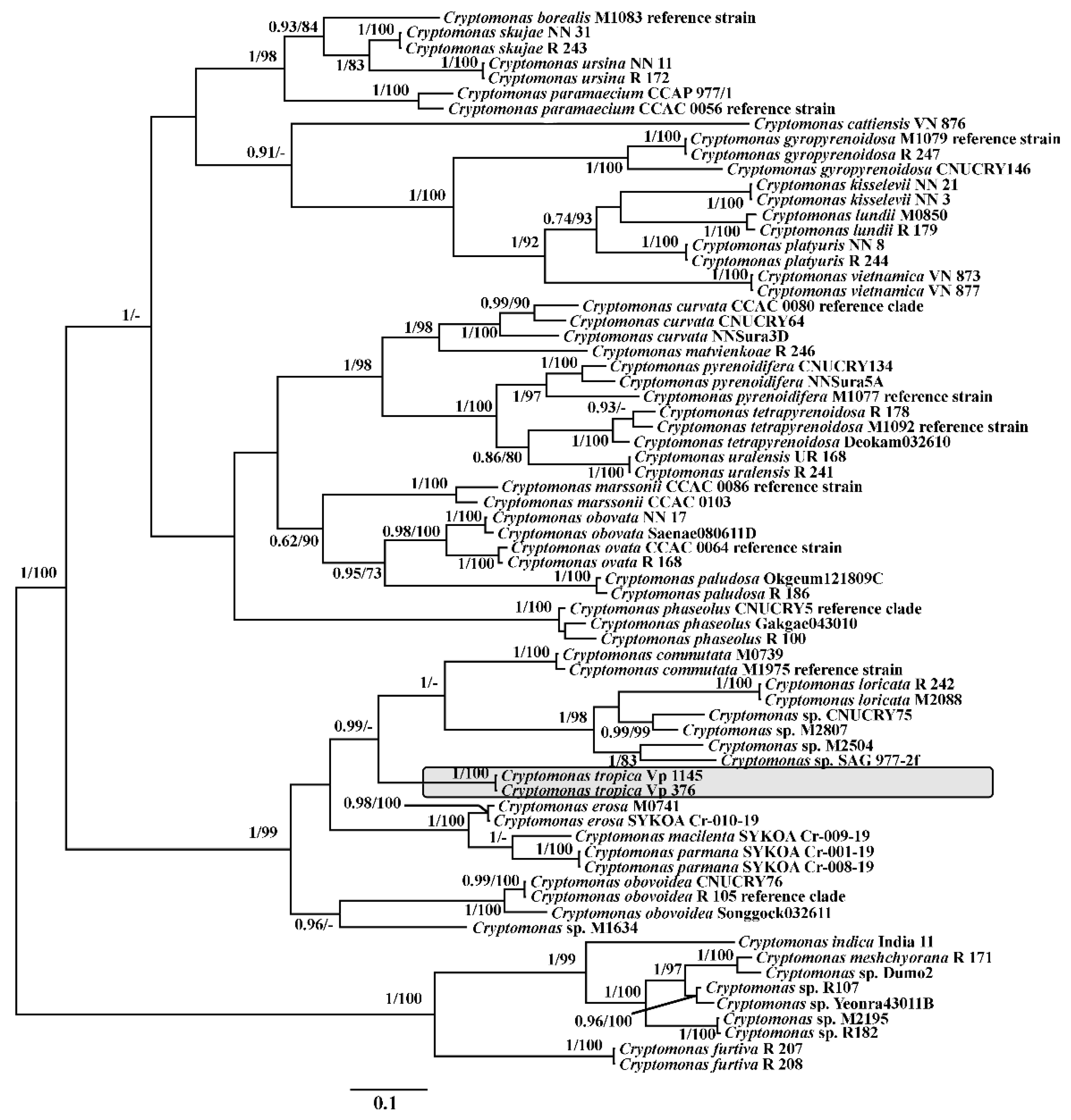
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ehrenberg, C.G. Animalia Evertebrata. Symbolae physicae seu icones et descriptiones animalium evertebratorum sepositis insectis quae ex itinere per Africanum Borealem et Asiam Occidentalem Friderici Guilelmi Hemprich et Christiani Godofredi Ehrenberg medicinae et chirurgiae doctorum studio novae aut illustratae redierunt. In Symbolae Physicae; Pars Zoologica, I.V., Hemprich, P.C., Ehrenberg, C.G., Eds.; Ex Officina Academica: Mittler, Berlin, 1831; 71p. [Google Scholar]
- AlgaeBase. Available online: https://www.algaebase.org/search/genus/detail/?genus_id=43571 (accessed on 13 October 2022).
- Hoef-Emden, K.; Melkonian, M. Revision of the Genus Cryptomonas (Cryptophyceae): A Combination of Molecular Phylogeny and Morphology Provides Insights into a Long-Hidden Dimorphism. Protist 2003, 154, 371–409. [Google Scholar] [CrossRef] [PubMed]
- Hoef-Emden, K. Revision of the genus Cryptomonas (Cryptophyceae) II: Incongruences between classical morphospecies concept and molecular phylogeny in smaller pyrenoid-less cells. Phycologia 2007, 46, 402–428. [Google Scholar] [CrossRef]
- Gusev, E.; Martynenko, N.; Kulizin, P.; Kulikovskiy, M. Molecular diversity of the genus Cryptomonas (Cryptophyceae) in Russia. Eur. J. Phycol. 2022, 57, 526–550. [Google Scholar] [CrossRef]
- Javornický, P.; Hindák, F. Cryptomonas frigoris spec. nova (Cryptophyceae), the new cyst-forming flagellate from the snow of the High Tatras. Biologia 1970, 25, 241–250. [Google Scholar] [PubMed]
- Gollerbakh, M.M.; Polyansky, V.I. Identification Guide for Freshwater Algae of the USSR; Sovetskaya Nauka: Moscow, Russia, 1953; Volume 6, 212p. [Google Scholar]
- Gollerbach, M.M.; Shtina, E.A. Soil Algae; Nauka: Saint-Petersburg, Russia, 1969; 228p. [Google Scholar]
- Matvienko, O.M.; Litvinenko, R.M. Identification Guide for Freshwater Algae of the Ukrainian SSR; Naukova Dumka: Kiev, Ukraine, 1977; Volume 3, 387p. [Google Scholar]
- Norton, J.R.; Davis, J.S. Soil Algae from North Central Florida. Fla. Sci. 1975, 38, 77–81. Available online: https://www.jstor.org/stable/24319630 (accessed on 14 September 2022).
- Hindák, F. Species composition and biomass of phototrophic edaphon in the ecosystem of a floodplain forest in southern Moravia. In Floodplain Forest Ecosystem. I. Before Water Management Measures; Penka, M., Vyskot, M., Klimo, E., Vašíček, F., Eds.; Academia, Publishing House of the Czechoslovak Academy of Sciences: Prague, Czechoslovakia, 1985; pp. 239–249. ISBN 0-444-99566-8. [Google Scholar]
- Paulsen, B.S.; Vieira, A.A.H.; Klaveness, D. Structure of extracellular polysaccharides produced by a soil Cryptomonas sp. (cryptophyceae). J. Phycol. 1992, 28, 61–63. [Google Scholar] [CrossRef]
- Choi, B.; Son, M.; Kim, J.I.; Shin, W. Taxonomy and phylogeny of the genus Cryptomonas (Cryptophyceae, Cryptophyta) from Korea. Algae 2013, 28, 307–330. [Google Scholar] [CrossRef]
- Martynenko, N.A.; Gusev, E.S.; Kulizin, P.V.; Guseva, E.E.; McCartney, K.; Kulikovskiy, M.S. A new species of Cryptomonas (Cryptophyceae) from the Western Urals (Russia). Eur. J. Taxon. 2020, 649, 1–12. [Google Scholar] [CrossRef]
- Martynenko, N.A.; Gusev, E.S.; Sterlyagova, I.N.; Kulikovskiy, M.S. Revealing hidden diversity in the Cryptomonas erosa clade (Cryptophyceae), with the description of two new species from acidic habitats. Phycologia 2022, 61, 184–194. [Google Scholar] [CrossRef]
- Gusev, E.; Podunay, Y.; Martynenko, N.; Shkurina, N.; Kulikovskiy, M. Taxonomic studies of Cryptomonas lundii clade (Cryptophyta: Cryptophyceae) with description of a new species from Vietnam. Fottea 2020, 20, 137–143. [Google Scholar] [CrossRef]
- Martynenko, N.A.; Gusev, E.S.; Kapustin, D.A.; Guseva, E.E.; Kulikovskiy, M.S. Cryptomonas cattiensis sp. nov. (Cryptophyceae: Cryptomonadales), a new species described from Vietnam. Phytotaxa 2020, 454, 127–136. [Google Scholar] [CrossRef]
- Gusev, E.; Karthick, B.; Martynenko, N.; Shkurina, N.; Kulikovskiy, M. Cryptomonas indica sp. nov. (Cryptophyceae: Cryptomonadales), a new species described from the Western Ghats, India. Phytotaxa 2021, 518, 261–270. [Google Scholar] [CrossRef]
- Blanc, L.; Maury-Lechon, G.; Pascal, J.-P. Structure, floristic composition and natural regeneration in the forests of Cat Tien National Park, Vietnam: An analysis of the successional trends. J. Biogeogr. 2000, 27, 141–157. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Anichkin, A.E. Cat Tien National Park—General information. In Structure and Functions of Soil Communities of a Monsoon Tropical Forest (Cat Tien National Park, Southern Vietnam); Tiunov, A.V., Ed.; KMK Scientific Press: Moscow, Russia, 2011; pp. 11–15. ISBN 978-5-87317-797-4. [Google Scholar]
- Kuznetsov, A.N.; Kuznetsova, S.P. Forest vegetation: Species composition and stand structure. In Structure and Functions of Soil Communities of a Monsoon Tropical Forest (Cat Tien National Park, Southern Vietnam); Tiunov, A.V., Ed.; KMK Scientific Press: Moscow, Russia, 2011; pp. 16–43. ISBN 978-5-87317-797-4. [Google Scholar]
- Cat Tien National Park. Available online: http://www.namcattien.org (accessed on 23 September 2022).
- Gusev, E.; Doan-Nhu, H.; Nguyen-Ngoc, L. Silica-scaled chrysophytes from Cat Tien National Park (Dong Nai Province, Vietnam). Nova Hedwig. Beih. 2017, 105, 347–364. [Google Scholar] [CrossRef]
- Gusev, E.S.; Guseva, E.E.; Dien, T.D.; Kulikovskiy, M.S. Flora of Silica-Scaled Chrysophytes (Chrysophyceae) of Two Provinces in Southern Vietnam. Inland Water Biol. 2022, 15, 205–216. [Google Scholar] [CrossRef]
- Glushchenko, A.; Genkal, S.I.; Kulikovskiy, M. Aulacoseira konstastinovii sp. nov. andAulacoseira kryloviisp. nov.—Two new centric diatoms from South-east Asia. Diatom Res. 2016, 31, 367–378. [Google Scholar] [CrossRef]
- Glushchenko, A.; Kulikovskiy, M.; Kociolek, J.P. New and interesting species from the genus Luticola (Bacillariophyceae) in waterbodies of Southeastern Asia. Nova Hedwig. Beih. 2017, 146, 157–173. [Google Scholar] [CrossRef]
- Glushchenko, A.; Kulikovskiy, M.; Kuznetsova, I.; Dorofeyuk, N.; Kociolek, J.P. New species and combinations in the genus Eunotia Ehrenberg 1837 (Bacillariophyceae: Eunotiaceae) from waterbodies of Southeastern Asia. Nova Hedwig. Beih. 2018, 147, 69–103. [Google Scholar] [CrossRef] [PubMed]
- Gusev, E.S. A new species of the genus Mallomonas (Synurophyceae), Mallomonas spinosa sp. nov., from Vietnam. Phytotaxa 2012, 66, 1–5. [Google Scholar] [CrossRef]
- Gusev, E.S.; Doan-Nhu, H.; Nguyen-Ngoc, L. Mallomonas cattiensis sp. nov. (Synurales, Chrysophyceae), a new species from Viet Nam. Phytotaxa 2015, 221, 188–192. [Google Scholar] [CrossRef]
- Gusev, E.S.; Doan-Nhu, H.; Nguyen-Ngoc, L.; Kapustin, D.A. Two new species of the genus Mallomonas from the Cat Tien National Park (Viet Nam): Mallomonas distinguenda and Mallomonas skvortsovii. Phytotaxa 2016, 273, 59–64. [Google Scholar] [CrossRef]
- Gusev, E.S.; Siver, P.A.; Shin, W. Mallomonas bronchartiana Compère Revisited: Two New Species Described from Asia. Cryptogam. Algologie 2017, 38, 3–16. [Google Scholar] [CrossRef]
- Gusev, E.S.; Čertnerová, D.; Škaloudová, M.; Škaloud, P. Exploring Cryptic Diversity and Distribution Patterns in the Mallomonas kalinae/rasilis Species Complex with a Description of a New Taxon—Mallomonas furtiva sp. nov. J. Eukaryot. Microbiol. 2018, 65, 38–47. [Google Scholar] [CrossRef]
- Gusev, E.S.; Kulizin, P.; Guseva, E.; Shkurina, N.; Kulikovskiy, M. Mallomonas lamii sp. nov. (Synurales, Chrysophyceae), a new species bearing large scales described from the tropics. Phytotaxa 2019, 423, 266–272. [Google Scholar] [CrossRef]
- Vadjunina, A.F.; Korchagina, Z.A. Methods of Studying the Physical Properties of Soils, 3rd ed.; Agropromizdat: Moscow, Russia, 1986; 416p. [Google Scholar]
- Arinushkina, E.V. Handbook for Chemical Soil Analysis, 2nd ed.; Moscow State University: Moscow, Russia, 1970; 488p. [Google Scholar]
- McFadden, G.I.; Melkonian, M. Use of Hepes buffer for microalgal culture media and fixation for electron microscopy. Phycologia 1986, 25, 551–557. [Google Scholar] [CrossRef]
- Marin, B.; Klingberg, M.; Melkonian, M. Phylogenetic Relationships among the Cryptophyta: Analyses of Nuclear-Encoded SSU rRNA Sequences Support the Monophyly of Extant Plastid-Containing Lineages. Protist 1998, 149, 265–276. [Google Scholar] [CrossRef]
- Kim, J.-H.; Boo, S.-M.; Shin, W.-G. Two Freshwater Cryptomonads New to Korea: Cryptomonas marssonii and C. pyrenoidifera. Algae 2007, 22, 147–152. [Google Scholar] [CrossRef]
- Hoef-Emden, K.; Marin, B.; Melkonian, M. Nuclear and Nucleomorph SSU rDNA Phylogeny in the Cryptophyta and the Evolution of Cryptophyte Diversity. J. Mol. Evol. 2002, 55, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef]
- Xia, X.; Xie, Z. DAMBE: Software Package for Data Analysis in Molecular Biology and Evolution. J. Hered. 2001, 92, 371–373. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Xie, Z.; Salemi, M.; Chen, L.; Wang, Y. An index of substitution saturation and its application. Mol. Phylogenetics Evol. 2002, 26, 1–7. [Google Scholar] [CrossRef]
- Xia, X.; Lemey, P. Assessing substitution saturation with DAMBE. In The Phylogenetic Handbook: A Practical Approach to DNA and Protein Phylogeny, 2nd ed.; Lemey, P., Salemi, M., Vandamm, A.-M., Eds.; Cambridge University Press: Cambridge, UK; pp. 615–630. [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef]
- Keller, A.; Schleicher, T.; Schultz, J.; Müller, T.; Dandekar, T.; Wolf, M. 5.8S-28S rRNA interaction and HMM-based ITS2 annotation. Gene 2009, 430, 50–57. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Schultz, J.; Maisel, S.; Gerlach, D.; Müller, T.; Wolf, M. A common core of secondary structure of the internal transcribed spacer 2 (ITS2) throughout the Eukaryota. Rna 2005, 11, 361–364. [Google Scholar] [CrossRef]
- Caisová, L.; Marin, B.; Melkonian, M. A Consensus Secondary Structure of ITS2 in the Chlorophyta Identified by Phylogenetic Reconstruction. Protist 2013, 164, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.J.; Johnston, J.S.; Cannone, J.J.; Gutell, R.R. Characteristics of the nuclear (18S, 5.8S, 28S and 5S) and mitochondrial (12S and 16S) rRNA genes of Apis mellifera (Insecta: Hymenoptera): Structure, organization, and retrotransposable elements. Insect Mol. Biol. 2006, 15, 657–686. [Google Scholar] [CrossRef]
- Wuyts, J.; Van de Peer, Y.; De Wachter, R. Distribution of substitution rates and location of insertion sites in the tertiary structure of ribosomal RNA. Nucleic Acids Res. 2001, 29, 5017–5028. [Google Scholar] [CrossRef]
- Byun, Y.; Han, K. PseudoViewer: Web application and web service for visualizing RNA pseudoknots and secondary structures. Nucleic Acids Res. 2006, 34, W416–W422. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Seibel, P.N.; Müller, T.; Dandekar, T.; Schultz, J.; Wolf, M. 4SALE—A tool for synchronous RNA sequence and secondary structure alignment and editing. BMC Bioinform. 2006, 7, 498. [Google Scholar] [CrossRef]
- Javornický, P. Cryptomonas ovata Ehrenberg (Cryptophyceae) and some related species. Algol. Studies. 2003, 110, 43–65. [Google Scholar] [CrossRef]
- Hoef-Emden, K.; Archibald, J.M. Cryptophyta (Cryptomonads). In Handbook of the Protists, 2nd ed.; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Margulis, L., Melkonian, M., Chapman, D.J., Corliss, J.O., Eds.; Springer: Cham, Switzerland, 2017; Volume 2, pp. 851–891. [Google Scholar] [CrossRef]
- Lichtlé, C. Effects of nitrogen deficiency and light of high intensity on Cryptomonas rufescens (Cryptophyceae) I. Cell and photosynthetic apparatus transformations and encystment. Protoplasma 1979, 101, 283–299. [Google Scholar] [CrossRef]
- Lichtlé, C. Effects of nitrogen deficiency and light of high intensity on Cryptomonas rufescens (Cryptophyceae) II. Excystment. Protoplasma 1980, 102, 11–19. [Google Scholar] [CrossRef]
- Santore, U.J. Light-and Electron-microscopic Observations of the Palmelloid) Phase in Members of the Genus Cryptomonas (Cryptophyceae). Arch. Für Protistenkd. 1978, 120, 420–435. [Google Scholar] [CrossRef]
- Kezlya, E.; Glushchenko, A.; Kociolek, J.P.; Maltsev, Y.; Martynenko, N.; Genkal, S.; Kulikovskiy, M. Mayamaea vietnamica sp. nov.: A new, terrestrial diatom (Bacillariophyceae) species from Vietnam. Algae 2020, 35, 325–335. [Google Scholar] [CrossRef]
- Kezlya, E.; Glushchenko, A.; Maltsev, Y.; Gusev, E.; Genkal, S.; Kuznetsov, A.; Kociolek, J.P.; Kulikovskiy, M. Placoneis cattiensis sp. nov.—A new, diatom (Bacillariophyceae: Cymbellales) soil species from Cát Tiên National Park (Vietnam). Phytotaxa 2020, 460, 237–248. [Google Scholar] [CrossRef]
- Kezlya, E.; Glushchenko, A.; Maltsev, Y.; Gusev, E.; Genkal, S.; Kociolek, J.P.; Kulikovskiy, M. Three New Species of Placoneis Mereschkowsky (Bacillariophyceae: Cymbellales) with Comments on Cryptic Diversity in the P. elginensis—Group. Water 2021, 13, 3276. [Google Scholar] [CrossRef]
- Foissner, W. Biogeography and dispersal of micro-organisms: A review emphasizing protists. Acta Protozool. 2006, 45, 111–136. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Taxa | Helix | ITS2 Length | K2P-Distances | |||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | SSU | LSU | psbA | ITS2 | |||
| 1 | C. erosa M0741 * | 0 | 2 | 3 | 364 | 0.040 | 0.134 | 0.030 | 0.269 |
| 2 | C. commutata M1975 | 2 | 2 | 2 | 343 | 0.048 | 0.126 | – | 0.244 |
| 3 | C. macilenta SYKOA Cr-009-19 | 0 | 0 | 4 | 416 | 0.040 | 0.140 | 0.026 | 0.313 |
| 4 | C. obovoidea M2811 ** | 0 | 2 | 2 | 370 | 0.054 | 0.155 | 0.030 | 0.273 |
| 5 | C. parmana SYKOA Cr-008-19 | 0 | 3 | 1 | 424 | 0.041 | 0.128 | 0.035 | 0.326 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martynenko, N.; Kezlya, E.; Gusev, E. Description of a New Species of the Genus Cryptomonas (Cryptophyceae: Cryptomonadales), Isolated from Soils in a Tropical Forest. Diversity 2022, 14, 1001. https://doi.org/10.3390/d14111001
Martynenko N, Kezlya E, Gusev E. Description of a New Species of the Genus Cryptomonas (Cryptophyceae: Cryptomonadales), Isolated from Soils in a Tropical Forest. Diversity. 2022; 14(11):1001. https://doi.org/10.3390/d14111001
Chicago/Turabian StyleMartynenko, Nikita, Elena Kezlya, and Evgeniy Gusev. 2022. "Description of a New Species of the Genus Cryptomonas (Cryptophyceae: Cryptomonadales), Isolated from Soils in a Tropical Forest" Diversity 14, no. 11: 1001. https://doi.org/10.3390/d14111001
APA StyleMartynenko, N., Kezlya, E., & Gusev, E. (2022). Description of a New Species of the Genus Cryptomonas (Cryptophyceae: Cryptomonadales), Isolated from Soils in a Tropical Forest. Diversity, 14(11), 1001. https://doi.org/10.3390/d14111001