Abstract
Loropetalum chinense var. rubrum is a common landscape plant. Because of the preference for the ‘red leaf’ trait, the ‘flower color’ trait was ignored in breeding, and the genetic diversity of L. chinense var. rubrum was replaced by a few genotypes. Therefore, the potential value of flower color traits was affected, and the genetic diversity was not fully exploited. In this study, we utilized a cultivar with mosaic-colored flowers and leaves and observed 15 phenotypic traits in its flower organs and its progeny, as well as a total of 60 different flower color materials. We analyzed the variation characteristics and diversity of flower colors, and qualitatively described and preliminarily classified flower colors using the ISCC-NBS and CIELab systems to explore the correlation with the flower color phenotype from the distribution and content of anthocyanins. The phenotypic diversity of the flower organs was enriched, and genetic differences in flower color were significant for the L. chinense var. rubrum, with most of the 15 phenotypic traits showing significant positive correlations, and the most critical phenotypes are sepal, petal and nectary color, as well as petal length. According to the CIELab color system and anthocyanin content and distribution characteristics, it is considered more accurate and convenient to classify the color phenotypes of L. chinense var. rubrum flowers into five categories. In this study, for the first time, the flower organ phenotype of L. chinense var. rubrum has been systematically analyzed, and it is explored as a reference method for breeding new cultivars of a diverse range of colors in L. chinense var. rubrum.
1. Introduction
Loropetalum chinense var. rubrum Yieh is a variety of Hamamelidaceae [1]. In contrast to the original species, this variety has red flowers and leaves, which greatly improves its ornamental value. It was first discovered in Changsha, Hunan Province, in 1942 [2]. The wild resources of L. chinense var. rubrum are distributed sporadically in the broad-leaved evergreen forests of Liuyang, Pingjiang, Liling and Luoxiao Mountains at altitudes of 100–400 m [3]. The city of Liuyang is a county-level city located in the northeast of Changsha, and it is known as ‘the hometown of L. chinense var. rubrum’ due to its large-scale planting and breeding [4]. L. chinense var. rubrum is also known as ‘The panda of plants’ by botanists due to its strong ecological adaptability, trimming resistance, easiness to shape, etc. [5]. L. chinense var. rubrum has many tree forms, such as scrubs, bulbs and small trees, and as its leaves are purple-red all year round, it has become a typical representative of plants with colorful leaves in landscape applications [6]. Moreover, L. chinense var. rubrum has bright flower colors, a large number of flowers, a long flowering period and can bloom 2–3 times a year; these characteristics have attracted a large amount of attention from landscape architects. At present, it is applied in garden cultivation not only in Changsha, Hunan, but also in most cities in southern China and some other parts of the world [7]. In recent years, the studies on L. chinense var. rubrum have mainly focused on cultivation and breeding [8,9,10], mechanisms of leaf color change [11,12,13], photosynthetic characteristics [14,15,16], etc. However, few studies have addressed phenotypic traits, especially in flower organs.
Plant phenotypic diversity is the result of the interaction between the environment and genetic diversity [17]. Studies on phenotypic diversity can clarify the structure, origin and variation of the population, and provide a scientific basis for the collection, conservation and genetic improvement of plant germplasm resources [18,19]. The genetic diversity of phenotypic traits has been proven to be an important scientific and effective method and has been successfully applied in Acorus calamus [20], ornamental peach [21], Hemerocallis [22] and other garden plants. The morphological diversity of floral organs is an important basis for genetic diversity, and its variation is of adaptive and evolutionary significance [23]. In the phenotypic analysis of the quantitative and qualitative characteristics of the flower organs of ornamental plants, it is found that the morphology of flower organs of different varieties of ornamental plants is highly diverse [24,25,26]. The wild and cultivated types of L. chinense var. rubrum also showed phenotypic diversity in the quantitative character of the flower [27,28]. The flower number of the wild type L. chinense var. rubrum was 4, approximately 50%, and that of the cultivated type was 5. The variation of the quantitative characters of flowers in recent cultivation was greater than that in early cultivation [29].
This indicated that L. chinense var. rubrum had a very plastic genetic space. The flower organs of L. chinense var. rubrum are very important ornamental features, and its flower color is a very important ornamental characteristic. Previous studies have shown that L. chinense var. rubrum can be divided into 3 groups, 15 types and 41 varieties according to the main characteristics of different flower colors, flower shapes, leaf shapes and branch densities [30], but they cannot provide a precise definition of flower color phenotypes or realize the standardization classification of the flower color phenotypes. At present, the phenotypic variation of flower organs, especially the phenotypic variation of flower color, in the progenies of L. chinense var. rubrum is still not clear. Additionally, flower color is the most intuitive phenotypic feature of flower organs and an important basis for variety classification [31,32,33]. Recently, the ISCC-NBS color names and CIELab system have been widely used in the classification of flower color phenotypes and have been scientifically validated in ornamental plants such as Paeonia rockii, Dendrobium, Chrysanthemum and Petunia hybrida [34,35,36,37]. The scientific and accurate classification of flower color is of great practical significance not only to the study of flower color formation mechanisms and breeding, but also to the classification of varieties and guidance of production. Therefore, exploring the variation degree of the phenotypic traits of flower organs of L. chinense var. rubrum and revealing its variation regularity are the key to accelerating the breeding of new varieties. The quantitative definition and classification of flower color are an important basis for variety classification, identification and breeding.
At present, cutting and grafting are the main methods of seedling propagation in L. chinense var. rubrum production. People prefer ‘Leaf color’ or ‘Flower color’, which makes a narrow genetic basis of L. chinense var. rubrum and the problem of relatively single varieties more and more serious. Therefore, it is very urgent and significant to study the floral organ phenotypic diversity of L. chinense var. rubrum. In this study, the phenotypic diversity of floral organs of the offspring of a self-selected chimera (L. chinense var. rubrum ‘Huayejimu1’) was analyzed and discussed; this is expected to provide a reference for the development of new varieties and resource types of L. chinense var. rubrum.
2. Materials and Methods
2.1. Plant Material
A total of 60 plant materials were used in this study. Among these, those numbered Lc1 were the female parents, an excellent variety recognized by the Hunan Forestry Bureau, which are named L. chinense var. rubrum ‘Huayejimu1’ (accession number is Xiang S-SV-LC-032-2012-1) and have ‘mosaic leaf’ characteristics. It is a unique variety with dual-colored ‘red + green’ leaves and dual-colored ‘white + red’ petals, which is stable and exhibits mottled color blocks. In particular, the high heterozygosity of its flower color and abundant genetic diversity mean that its progeny is diversified.
The other 59 plant materials are the progeny formed by the natural pollination of Lc1 material. These progenies are 9-year-old plants screened through sowing and seedling raising, and are numbered from Lc2 to Lc60. The flower organs of the 60 plant materials were used as the research object materials. The used plant materials were planted and preserved in the flower nursery of Hunan Agricultural University and Hunan Mid-subtropical Quality Plant Breeding and Utilization Engineering Technology Research Center.
2.2. Measurement of Phenotypic Traits of Flower Organs
The flowers in full bloom were selected for the analysis of phenotypic diversity. The 15 phenotypic traits of 60 L. chinense var. rubrum plants were investigated. The phenotypic traits were observed and measured on the basis of GB/T 19557.1-2004 (National standard) [38], and the flower color was determined using the RHS Colour Chart published by the British Royal Horticultural Society [39]. Measurements were repeated seven times for each index and averaged. The standard for determining specific phenotypic traits is shown in Table 1.

Table 1.
Morphological diversity indexes and description of L. chinense var. rubrum.
2.3. Determination of L*, a* and b* Parameters of CIELab Chromaticity System
CIELab color space consists of lightness (L*), redness (a*) and yellowness (b*). L* denotes the change in brightness from black to bright; a* represents the color change from green to red; b* represents the color change from blue to yellow. L*, a* and b* were measured directly by grating spectrometer (YS3010, 3 nh, China). The center position of the petals was measured with a D65 light source and a 10° angle of view, and each flower was measured seven times to get an average. According to the hue a* and b*, to calculate chroma C*, the formula is C* = (a*2 + b*2)1/2. The C* value represents the vertical distance to the L* axis, and the greater the distance, the greater the chroma.
2.4. Determination of Name of Flower Color Based on ISCC-NBS Method
The measured L*, a* and b* values were imported into R 4. 1. 2 (WU Vienna, Austria) software, using the Munsell Interpol model to convert them into H (Munsell Hue), V (Munsell Value) and C (Munsell Chroma) values, which were compared with the ISCC-NBS color name representation to qualitatively determine the flower color. Each flower was measured seven times, and the average value was taken for subsequent analysis.
2.5. Distribution and Content Determination of Anthocyanins in Flowers
Three petals were placed in a Petri dish half filled with distilled water. Using two razor blades, thin slices were cut quickly to create a temporary water container. Bubbles and folding were avoided, and excess moisture was siphoned off with a filter. The temporary waterboard was placed under a microscope (DMIL LED, Leica, Wetzlar, Germany) to observe the pigmentation distribution of the petals by color depth, and photos were taken.
Color depth indicates the brightness level of the color. The higher the content of anthocyanin, the more pigment distribution and the darker the color.
Anthocyanin was determined using the pH differential method for the cyanidin-3-glucoside content in plants [40]. The anthocyanin content was calculated using the formula TA = A × MW × 5 × 100 × V/e [41], where TA represents the total anthocyanin content (mg/100 g) and V represents the final volume (mL). A = pH 1.0 (A510–A700)–pH 4.5 (A510–A700). According to Wrolstad et al. [42], the molar absorptivity (e) was 26,900 and the molecular weight (MW) was 449.2. Anthocyanin content thresholds refer to the numerical range of anthocyanin content. Three measurements were performed for each biological replication.
2.6. Statistical Analysis
The experimental data were processed with Excel 2019. SPSS 22.0 was used to calculate the mean, maximum, minimum, standard deviation (SD), coefficient of variation (CV) and genetic diversity index (H’) of the phenotypic traits in the flower organs of L. chinense var. rubrum. CV is a normalized measure of the discrete degree of probability distribution, and the formula is CV = (SD/Mean) × 100%. H’ is based on the Shannon–Wiener Information Index, and the formula is H’ = −∑ Pi∙InPi. Pi is the percentage of the number of materials at the ith level of a phenotypic trait in the total number of materials. Origin 2021 (Origin, Northampton, MA, USA) software was used for principal component analysis and correlation analysis of flower color phenotypic traits. The flower color classification used was the European distance method and was analyzed by Origin 2021 software. The distribution of anthocyanins in petals, calyx and nectaries were plotted by Adobe Photoshop CC 2019 (Adobe, San Jose, CA, USA).
3. Results
3.1. Phenotypic Analysis of Flower Organs in L. chinense var. rubrum
3.1.1. Analysis of Genetic Diversity of Phenotypic Traits in Floral Organs
The genetic diversity of 60 flower organs of L. chinense var. rubrum was analyzed. The phenotypic data (Table 2 and Table 3) showed that the coefficient of variation (CV) of quantitative traits ranged from 5.24% to 32.55%, and the average of the CV was 15.77%; the coefficient of variation of anthocaulus length (AL) was the largest, and that of petal number (PN) was the smallest. The genetic diversity index (H’) of quantitative traits ranged from 1.38 to 2.00, and the mean value of H’ was 1.84. The H’ of flower number (FN) was the highest, and the H’ of the stamen number (SN) was the lowest, which indicated that the genetic difference in quantitative traits was large. The results of the frequency distribution of quality traits showed that the frequency distribution of quality traits in different distribution regions was unbalanced. The H’ value of quality traits varied from 1.02 to 1.57, among which the H’ value of the nectary color (NC) was the highest, followed by the H’ value of the petal color (PC) and sepal color (SC), and the H’ value of the petal form (PF) was the lowest. The results showed that there was rich diversity in flower color.
Table 2.
Genetic diversity analysis of quantitative characters in flower organs of L. chinense var. rubrum.
Table 3.
Frequency distribution and diversity of quality traits in flower organs of L. chinense var. rubrum.
3.1.2. Principal Component Analysis Based on Phenotypic Traits
Principal component analysis (PCA) was carried out on phenotypic traits in flower organs of 60 L. chinense var. rubrum (Table 4). According to the principle that the characteristic value is higher than 1, 6 principal components were screened, and the cumulative contribution rate was 79.55%. These 6 principal components could reflect the basic characteristics of 15 phenotypic traits. The contribution rate of the first principal component was 23.64%. The phenotypic traits with a larger eigenvector were CD, CLD and PL, which were mostly related to corolla and reflected the corolla characteristics of flower organs. The contribution rate of the second principal component was 16.30%. The traits with larger eigenvector values were SC, NC and PC. These phenotypic traits were all related to color and reflected the color characteristics of flower organs. The 3rd and 4th principal components reflect the quantitative characteristics of flower organs. The 5th and 6th principal components reflect the petal characteristics of flower organs. Similar results can be observed in Figure 1, where the cumulative contribution of principal components 1, 2, and 3 is 53.14%. Therefore, the main characteristic vectors of 3 principal components, namely, corolla characteristics, flower color characteristics and quantitative characteristics, can be used as important morphological indexes for the evaluation of new parent varieties of L. chinense var. rubrum.
Table 4.
Principal component analysis of phenotypic traits of L. chinense var. rubrum.
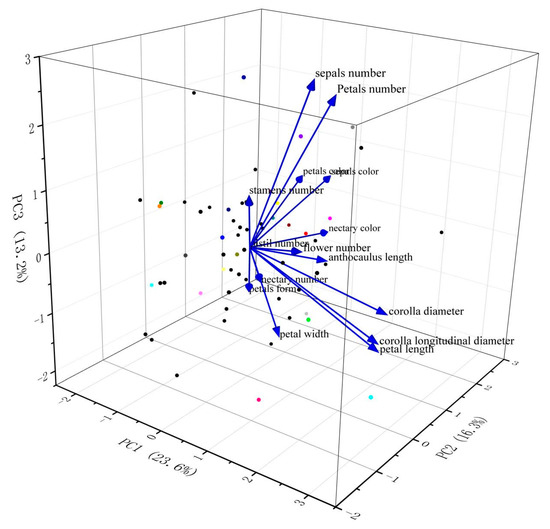
Figure 1.
Principal component analysis diagram of 60 L. chinense var. rubrum.
3.1.3. Correlation Analysis of Phenotypic Traits in Flower Organs
The phenotypic traits of L. chinense var. rubrum were correlated to different degrees (Figure 2). Among the 17 pairs of phenotypic traits, 7 pairs of phenotypic traits were highly significantly correlated, 2 pairs of phenotypic traits were extremely significantly correlated, 8 pairs of phenotypic traits were significantly correlated, and all of them were positively correlated. The three pairs of NC and SC, SC and PC, PC and NC showed a very significant correlation, which indicated that there was a close correlation between flower color traits of L. chinense var. rubrum. It can be further classified and analyzed for its color phenotype.
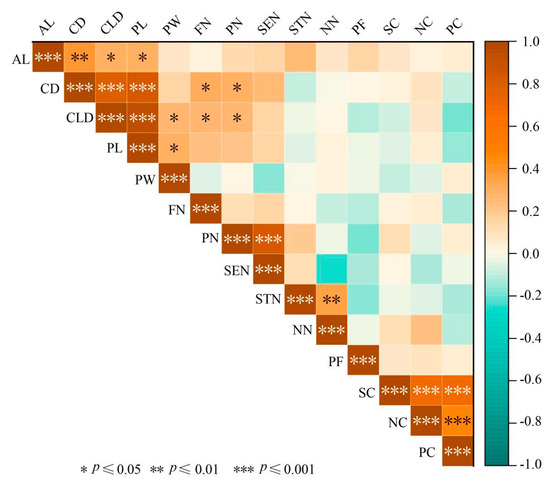
Figure 2.
Correlation coefficients of phenotypic traits in 60 plants of L. chinense var. rubrum. Note: Yellow-brown indicates a positive correlation; light blue indicates a negative correlation; the darker the color, the stronger the correlation.
3.2. Classification of Flower Color Phenotypes of L. chinense var. rubrum
3.2.1. Flower Color Naming and Classification Based on ISCC-NBS Method
We measured the flower color phenotype and chromaticity values of 60 samples of L. chinense var. rubrum from 1 mother and 59 progeny (attached list), and the measured L*, a* and b* values were converted into H (Hue), V (Value) and C (Chroma) values of the Munsell Color System (Appendix A Table A1). Then, combined with H, V and C values, the ISCC-NBS method was used to qualitatively describe the color of each sample, and finally, they were categorized into 22 colors (Table 5). After simplifying the V values and C values, except for the polychromatic samples, the flower colors of other samples were classified into five color groups: green-white, yellowish white, light pink, reddish purple and purple. However, according to the observation and analysis of the actual flower colors, the green-white and yellowish white colors were easily mixed and considered as white (35% of the total), and it is difficult to distinguish light pink, reddish purple or purple among the samples with similar flower colors (61.67% of the total). Therefore, it can be considered that the ISCC-NBS method does not particularly accurately classify the flower colors at the edge of the Munsell color system, and other more clear and concise classification methods need to be further explored.
Table 5.
The classification of L. chinense var. rubrum samples based on ISCC-NBS method of designating colors.
3.2.2. Flower Color Classification Based on CIELab Data Clustering Analysis
Based on the previous qualitative description of the ISCC-NBS method and the actual investigation experience, the flower colors were classified into six groups (Table 6): polychromatic, green-white, yellowish white, light pink, reddish purple and purple. We analyzed the CIELab parameter distribution range of these six groups of samples and found that (Table 6) the brightness of the green-white group is the greatest, and the distribution range of the L* value is 78.30–86.65; the brightness of the light pink group is the lowest, and its L* value distribution range is 62.85–63.95. The a* value of the purple group is slightly higher than that of the reddish purple, and the a* value of the reddish purple group is relatively concentrated; it also can be seen that the a* values of the reddish purple group and the purple group in the samples are relatively close and difficult to distinguish. The maximum b* value of the yellowish white group is 22.35, which also reflects the lack of a yellow group in L. chinense var. rubrum samples. According to the above analysis results, this classification method is not suitable for the color classification of L. chinense var. rubrum varieties.
Table 6.
The distribution range of flower color—L*, a* and b* parameters of each color group in L. chinense var. rubrum.
Therefore, we constructed a cluster analysis based on the L*, a* and b* values of the 60 samples. From the cluster diagram (Figure 3), it is clear that the sample branches can be clustered into five groups: I polychromatic, II purplish pink, III purplish red, IV green-white and V yellowish white. Among them, the number of samples classified as the purplish red group is the largest, followed by the green-white group. Compared with the six groups categorized using the ISCC-NBS method, the three groups of light pink, reddish purple and purple are simplified into two groups of purplish pink and purplish red. It is not difficult to see that the classification result is simpler.
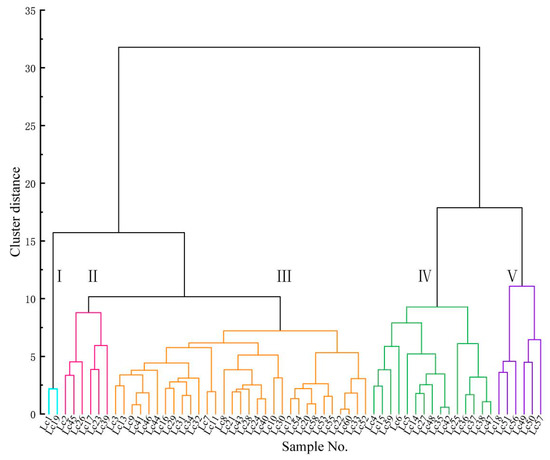
Figure 3.
Cluster analysis of flower color in 60 L. chinense var. rubrum based on L*, a* and b* parameters.
3.2.3. Flower Color Classification Based on L*, a* and b* Parameter Feature Analysis
According to the color grouping results of L*, a* and b* parameter clustering analysis, the distribution characteristics of the L*, a* and b* values of each group of colors were analyzed (Figure 4A). The analysis results show that the L* value is arranged from high to low in order of green-white, yellowish white, purplish pink, polychromatic and purplish red, and the brightness of the green-white group is greater than that of other color groups. Among the five groups, the green-white and yellowish white groups have overlapping areas in the a* value distribution, and the purplish pink and purplish red groups have overlapping areas. These L. chinense var. rubrum varieties are more closely distributed in the red axis area. Although the a* value of the polychromatic group is similar to the distribution of the purplish pink and purplish red groups, there is no overlapping area, indicating that the red part of the polychromatic group samples is different from the red part of other groups. The b* value of the yellowish white group is higher than that of the green-white group, and the difference in the distribution of the two groups of samples in the yellow axis area is also an important feature that can differentiate between the two groups. Since the polychromatic samples contain two colors, the b* value of this group completely overlaps with that of the green-white group, indicating that the light color in the polychromatic samples is similar to the green-white group.
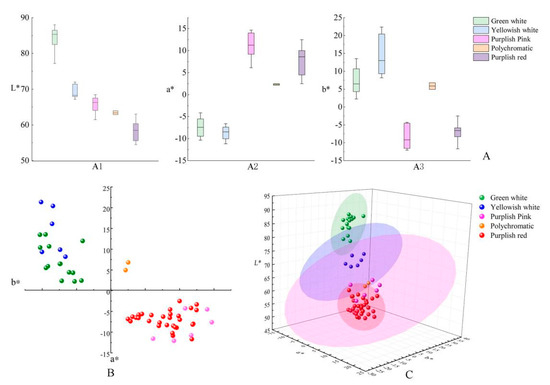
Figure 4.
The box plot, 2D and 3D distribution diagram of L*, a* and b* values of different color groups of L. chinense var. rubrum. Note: (A) The box plot of L*, a* and b* values, A1 box plot of L* value for different color groups; A2 box plot of a* value for different color groups; A3 box plot of b* value for different color groups. (B) The 2D distribution diagram of L*, a* and b* values of different color groups. (C) The 3D distribution diagram of L*, a* and b* values of different color groups.
In the two-dimensional (2D) distribution diagram of parameters a* and b* (Figure 4B) and the three-dimensional (3D) distribution diagram of L*, a* and b* (Figure 4C), the distribution of each color group is concentrated, and the boundary is obvious. It was further proved that the five color groups divided by the CIELab color system were more consistent with the actual flower color characteristics of L. chinense var. rubrum varieties than other methods.
3.2.4. Observation of the Distribution of Anthocyanins in Petals
The distribution of anthocyanins on the cross-section of petals was observed using a microscope (Figure 5); it was found that the anthocyanins in L. chinense var. rubrum are mainly distributed in sponge tissue cells, the approximate content can be preliminarily judged from the color depth, and the anthocyanin distribution in different flower color groups also presents different characteristics. The most obvious is that there is no macroscopic anthocyanin distribution in the petals of the green-white group and the yellowish white group (Figure 5a,b), so the petals are white, but the petals of the other three flower color groups (Figure 5c–e) of samples contain different amounts of anthocyanins. The anthocyanin content of the purplish red group (Figure 5c) is the most evenly distributed in the whole petal, fills sponge tissue cells and has a bright color. Second, for the purplish pink group (Figure 5d) with a lighter color, the anthocyanin content in the petals is lower, the anthocyanin distribution is relatively loose, it does not fill all sponge tissue cells and there are no anthocyanins in some cells. As a polychromatic group of samples (Figure 5e), the petals of the anthocyanins are more random, with densely distributed cell areas, loosely distributed cell areas and areas without anthocyanins. Moreover, the anthocyanin content in the cells containing anthocyanin is uneven, and the content difference between cells is obvious. From the distribution and content of anthocyanins in petal cells, these five kinds of flower color samples have obvious and prominent characteristics, which can better distinguish them.

Figure 5.
The distribution pattern of anthocyanins in flower of L. chinense var. rubrum. Note: (a) polychromatic group; (b) green-white group; (c) yellowish white group; (d) purplish red group; (e) purplish pink group.
In order to further accurately compare the difference in anthocyanin content in the petals of each group and help to confirm the results of the cross-sectional observation, the anthocyanin concentration of all samples was determined and is shown in Table 7. The anthocyanin content thresholds from high to low are purplish red, purplish pink, polychromatic, yellowish white, and green-white. The yellowish white and green-white samples contain minimal anthocyanins, and the threshold values of the two groups are partially overlapped. The content of the purplish pink and purplish red groups is obviously the highest, and the threshold values of the two groups are also partially overlapped. However, the threshold range of anthocyanin content in the purplish red group (97.06–334.67) is larger than that in the purplish pink group (99.43–264.96); in particular, the samples with a high anthocyanin content have a significantly higher value than the maximum value of the purplish pink group. For the mosaic polychromatic group samples with red and white petals, their anthocyanin content is in the middle range compared with the other groups, and the variation in anthocyanin content within the group is small. The component comparison results of these anthocyanin content determinations are consistent with the observation results of petal sections.
Table 7.
The threshold distribution of anthocyanin content in petals of different colors of L. chinense var. rubrum.
3.2.5. Observation of the Distribution of Anthocyanins in Calyx and Nectary
When we dissected and observed the flowers, we found an interesting phenomenon; that is, the colors of the calyx and nectary of petals of various colors are different. After the color classification, it was also confirmed that this phenomenon was consistent in each group and different among groups. As shown in Figure 6, the calyx and nectary of the green-white group and the yellowish white group with white petals are all green in different shades, while the calyx and nectary of the samples with red petals are all red. In the polychromatic group of various red and white petals, for example, the calyx and nectary of the flower of the mother sample Lc1 (Figure 6) are all green with a small amount of red, while the calyx and nectary of the purplish pink group samples have less green and more red. In the purplish red group samples (Figure 6), the green of their calyx and nectary almost disappears and changes to dark red.

Figure 6.
Analysis of calyx and nectary of different color groups of L. chinense var. rubrum. Note: Lc1 represents the polychromatic group; Lc5 represents the yellowish white group; Lc51 represents the green-white group; Lc26 represents the purplish pink group; Lc29 represents the purplish red group.
4. Discussion
4.1. Diversity Analysis of Flower Organ Phenotypic Traits of L. chinense var. rubrum
Compared with other horticultural crops, ornamental plants have been studied more in phenotypic determination and pigment analysis based on flower organs. As a major ornamental part and sexual reproduction organ, improving the variation of flower organs in offspring can help enrich flower color breeding. In this study, the offspring of a self-selected chimera (L. chinense var. rubrum ‘Huayejimu1’) had obvious variation in flower color, and the offspring had the mosaic flower color trait of the genetic mother parent, but there were also variations that separated different flower colors, showing the phenotypic diversity of flower organs. Therefore, in this study, 15 phenotypic traits of 60 flower organs of L. chinense var. Rubrum were analyzed through diversity analysis, principal component analysis and correlation analysis. In the diversity analysis, the coefficient of variation (CV) is a normalized measure of the dispersion of probability distribution, and the CV is more than 10%, which indicates that the difference in this character among different germplasms is significant [43]. The mean value of the CV of the flower organ phenotypic quantitative traits in L. chinense var. rubrum was 15.77%, and the largest value was 32.55%; the results of the frequency distribution of quality traits also showed a diverse range of flower colors. This also showed that the population had strong potential for diversity improvement. The diversity index (H’) is an important index for the evaluation of the diversity of germplasm resources [44]. The diversity index of the phenotypic traits of L. chinense var. rubrum was 1.02–2.00. The traits with a large to small variation coefficient were not consistent with the traits with a large to small diversity index. This was because the diversity index reflected the quantitative distribution of different grades of a trait, that is, the abundance and evenness of diversity, while the variation coefficient reflected the range in variation of the traits. Therefore, the performance of the coefficient variation and diversity index was not consistent for the same trait, indicating that the variation range was large. This is similar to the phenotypic results of Sesame, Rosa hybrid, Lotus and Chestnut [45,46,47,48]. Through principal component analysis, 6 principal components were screened, and the cumulative contribution rate was 79.55%. These 6 principal components could reflect the basic characteristics of 15 phenotypic traits, which reflected the genetic diversity of the floral organ phenotypic traits of L. chinense var. rubrum. As a flowering plant, flower color has a more obvious effect on the whole plant. In this study, the flower color feature vector was screened, which can provide an important reference value for breeding new varieties of L. chinense var. rubrum. Trait correlation can indirectly reflect the phenotype of another trait through the phenotype of one trait, improving selection efficiency and speeding up the breeding process [49]. The correlation analysis showed that the three pairs of phenotypic shapes related to the flower color of L. chinense var. rubrum showed a highly significant correlation, which indicated that the flower color traits were closely related to each other and provided a reference for the classification of flower color phenotypes. It can be seen from the above that among the varieties of L. chinense var. rubrum produced by L. chinense var. rubrum ‘Huayejimu1’ with flower color mosaicism, the most important one is that there are many flower color variations, and the differences are obvious. This unique flower color mosaicism can produce new flower color varieties efficiently with a certain degree of character separation when it is passed on to offspring.
4.2. Taxonomic Analysis of Flower Color Phenotype of L. chinense var. rubrum
The color measurement method of the Royal Horticultural Society Color Chart (RHSCC) has been applied to plant materials since 1776 and has been widely used in the color measurement of various plants [50]. Regarding the analysis of the flower color diversity of this ornamental plant, the flower color phenotype was only observed in a study report on the phenotypic analysis of 23 varieties of L. chinense var. rubrum published by Bao Zhiyi [51], but the flower color determination at that time was based on the R.H.S.C.C color card comparison method. At that time, they classified the flower color comparison results of all 23 samples into only two types of flower colors, 22 varieties were compared into a red purple group, and only one variety was different, with a few light pink flowers in summer. This result also shows that the traditional flower color phenotype analysis method cannot precisely analyze and classify the flower color phenotype of L. chinense var. rubrum. In order to make up for this shortcoming in the analysis of the flower color phenotype of L. chinense var. rubrum, the number of varieties with a flower color phenotype of 60 cultivated varieties was classified. The results showed that after the L*, a* and b* parameters were converted into H, V and C values, many colors of L. chinense var. rubrum could be accurately named using the ISCC-NBS method. However, the color classification results of this method often produced errors compared with the actual observation results. Therefore, we further analyzed the L*, a* and b* parameters, and used them for clustering and distribution analysis. Finally, according to the color classification, the 60 varieties were classified into five groups: polychromatic, yellowish white, green-white, purplish pink and purplish red. In order to verify the accuracy and applicability of this flower color classification result, in this study, the anthocyanin distribution observation and content determination of their petals and the color characteristics in the calyx and nectary were analyzed to verify that the flower color classification classified into five groups is more accurate and simple. It was also found that there was no yellow color in the L. chinense var. rubrum flowers. In the past, studies on the color of ornamental plants, including color rendering mode, pigment chemistry, gene function and breeding strategies, mainly focused on some traditional famous flowers in China. In peony, orchid, lily and other plants, flower color classification, anthocyanin distribution and flower color content have been analyzed, and the related research on the anthocyanin metabolism pathway is also more in-depth [35,52,53,54]. These achievements are the basis for the research of new flower color breeding. It can also be found from this study that the flower color of L. chinense var. rubrum is not only red or purplish red. By using this special variety L. chinense var. rubrum, we can obtain white flowers similar to the flower of L. chinense and obtain the purplish pink and purplish red flowers similar to the flower of L. chinense var. rubrum. In addition, we can obtain polychrome flowers mixed with the flower color of L. chinense and L. chinense var. rubrum. With the increase in the breeding work in L. chinense var. rubrum over the last decade, the variation in flower color has gradually diversified. In the future, we can consider selecting and breeding more varieties with different colors from these special materials to obtain new germplasm more quickly. It is expected that the classification results of this study can make up for the shortcoming in the analysis of the flower color phenotype of L. chinense var. rubrum and provide a reference for future breeding work to create more diversified varieties.
5. Conclusions
In this study, the phenotypic characteristics of flower organs of 60 varieties of L. chinense var. rubrum were comprehensively and systematically analyzed for the first time. We found that the phenotypic traits of varieties were quite different, and the 59 progenies were considered as different varieties. Moreover, from the analysis results of the 15 phenotypic traits, it can be seen that the variation among the germplasms of the petal, nectary and sepal color is the highest and the most highly correlated. According to the more suitable CIELAB method and the anthocyanin content and distribution, the 22 flower colors of these varieties after naming can be divided into five color groups, which were polychromatic, yellowish white, green white, purplish pink and purplish red. It was also found that there was no yellow in the flower colors. It must be mentioned that this unique variety of L. chinense var. rubrum ‘Huayejimu1′ with dual-colored ‘red + green’ leaves and dual-colored ‘white + red’ petals has generated a large number of variations in the genetic process of its offspring, which are mainly reflected in the color characteristics of flower organs, thus, greatly saving time for flower color breeding to cultivate more varieties of flower colors. In the future, we can speed up breeding work by applying the naming and classification method to enrich the flower color, enrich the flower color resources of L. chinense var. rubrum and increase the flower’s ornamental value.
Author Contributions
L.Z., X.Y., Y.L. and L.X. (Lu Xu) collected the sample conceived and designed the study; X.Z. and D.Z. observed and measured the phenotypic traits of the flower organs; W.L., L.X. (Lili Xiang) and Y.Y. performed the data analysis; L.Z., X.Y., Y.L. and L.X. (Lu Xu) wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The work is funded by the Forestry Science and Technology Innovation Foundation of Hunan Province for Distinguished Young Scholarship (XLKJ202205), The Found of Changsha Municipal Science and Technology Bureau (KQ2202227), Key Research and Development Program of Science and Technology Department of Hunan Province (2021NK2008), Hunan Provincial Education Department Teaching reform Project (2021JGYB101), Open Project of Horticulture Discipline of Hunan Agricultural University (2021YYXK001), Hunan Provincial Natural Science Youth Foundation Project (2020JJ5264), Hunan Agricultural University Teaching reform research project (XJJG-2020-071), National Key Research and development program (2019YFD1100400).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated/analyzed during this study are included in this article.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Appendix A
Table A1.
The flower color measured value based on CIELab with Munsell and color classification in L. chinense var. rubrum cultivars.
Table A1.
The flower color measured value based on CIELab with Munsell and color classification in L. chinense var. rubrum cultivars.
| Plant Code | CIELab Color Coordinate | Munsell Color System | Color Group According to CIELab | |||||
|---|---|---|---|---|---|---|---|---|
| L* | a* | b* | C* | H | V | C | ||
| Lc1 | 62.85 | 2.14 | 4.95 | 5.39 | 7.45YR | 6.18 | 0.87 | Polychromatic |
| Lc19 | 63.95 | 2.52 | 6.79 | 7.24 | 8.08YR | 6.29 | 1.15 | Polychromatic |
| Lc4 | 78.30 | −9.16 | 5.87 | 10.88 | 1.60G | 7.77 | 1.5 | Green-white |
| Lc14 | 86.46 | −4.28 | 2.43 | 4.92 | 2.44G | 8.61 | 0.64 | Green-white |
| Lc27 | 85.00 | −5.28 | 2.23 | 5.73 | 4.59G | 8.46 | 0.79 | Green-white |
| Lc35 | 83.34 | −5.51 | 4.31 | 6.99 | 0.39G | 8.29 | 0.89 | Green-white |
| Lc42 | 83.36 | −6.07 | 4.43 | 7.52 | 0.83G | 8.29 | 0.97 | Green-white |
| Lc48 | 86.15 | −7.15 | 2.37 | 7.53 | 6.40G | 8.58 | 1.07 | Green-white |
| Lc59 | 82.51 | −9.44 | 5.49 | 10.92 | 2.25G | 8.2 | 1.48 | Green-white |
| Lc5 | 88.00 | −7.35 | 6.47 | 9.79 | 9.70GY | 8.77 | 1.19 | Yellowish white |
| Lc6 | 77.19 | −8.99 | 10.66 | 13.94 | 7.55GY | 7.65 | 1.82 | Yellowish white |
| Lc15 | 79.87 | −7.42 | 6.45 | 9.83 | 9.66GY | 7.93 | 1.28 | Yellowish white |
| Lc25 | 87.53 | −4.11 | 11.98 | 12.67 | 1.24GY | 8.72 | 1.41 | Yellowish white |
| Lc36 | 86.65 | −8.99 | 10.66 | 13.94 | 7.74GY | 8.63 | 1.68 | Yellowish white |
| Lc37 | 85.62 | −10.29 | 10.41 | 14.64 | 8.79GY | 8.52 | 1.83 | Yellowish white |
| Lc38 | 85.30 | −10.33 | 13.47 | 16.98 | 7.18GY | 8.49 | 2.09 | Yellowish white |
| Lc47 | 85.99 | −9.51 | 13.55 | 16.56 | 6.73GY | 8.56 | 2 | Yellowish white |
| Lc18 | 68.13 | −10.03 | 9.34 | 13.71 | 9.01GY | 6.72 | 1.93 | Yellowish white |
| Lc49 | 68.40 | −11.15 | 22.35 | 24.98 | 4.35GY | 6.74 | 3.35 | Yellowish white |
| Lc50 | 71.39 | −8.43 | 20.40 | 22.07 | 2.63GY | 7.05 | 2.86 | Yellowish white |
| Lc51 | 67.93 | −6.61 | 8.17 | 10.51 | 7.04GY | 6.70 | 1.40 | Yellowish white |
| Lc56 | 71.96 | −7.39 | 9.88 | 12.33 | 6.73GY | 7.11 | 1.62 | Yellowish white |
| Lc57 | 67.16 | −8.55 | 16.11 | 18.24 | 4.72GY | 6.62 | 2.40 | Yellowish white |
| Lc2 | 67.50 | 14.01 | −4.53 | 14.72 | 1.80RP | 6.66 | 3.89 | Purplish Pink |
| Lc17 | 61.46 | 9.18 | −12.08 | 15.17 | 3.31P | 6.05 | 3.84 | Purplish Pink |
| Lc23 | 64.07 | 11.69 | −10.76 | 15.89 | 5.93P | 6.31 | 4.06 | Purplish Pink |
| Lc26 | 66.14 | 14.68 | −7.61 | 16.53 | 9.49P | 6.52 | 4.27 | Purplish Pink |
| Lc39 | 66.39 | 6.14 | −11.58 | 13.11 | 0.94P | 6.55 | 3.46 | Purplish Pink |
| Lc45 | 68.46 | 10.81 | −4.28 | 11.63 | 0.86RP | 6.76 | 3.17 | Purplish Pink |
| Lc3 | 56.83 | 11.98 | −8.00 | 14.41 | 7.80P | 5.58 | 3.58 | Purplish red |
| Lc7 | 55.59 | 12.46 | −4.67 | 13.30 | 1.46RP | 5.46 | 3.32 | Purplish red |
| Lc8 | 62.60 | 7.08 | −7.70 | 10.46 | 5.02P | 6.16 | 2.68 | Purplish red |
| Lc9 | 58.10 | 9.08 | −8.47 | 12.42 | 5.83P | 5.71 | 3.1 | Purplish red |
| Lc10 | 63.00 | 12.49 | −3.32 | 12.92 | 2.60RP | 6.2 | 3.37 | Purplish red |
| Lc11 | 57.29 | 11.66 | −4.18 | 12.39 | 1.62RP | 5.63 | 3.13 | Purplish red |
| Lc12 | 56.20 | 4.08 | −6.03 | 7.28 | 2.48P | 5.52 | 1.84 | Purplish red |
| Lc13 | 59.27 | 11.89 | −8.17 | 14.43 | 7.57P | 5.83 | 3.65 | Purplish red |
| Lc16 | 55.36 | 9.03 | −10.86 | 14.12 | 3.96P | 5.44 | 3.44 | Purplish red |
| Lc20 | 55.30 | 3.15 | −6.28 | 7.03 | 0.31P | 5.43 | 1.8 | Purplish red |
| Lc21 | 61.60 | 8.58 | −6.54 | 10.79 | 6.92P | 6.06 | 2.77 | Purplish red |
| Lc22 | 60.77 | 3.66 | −6.52 | 7.48 | 1.17P | 5.97 | 1.98 | Purplish red |
| Lc24 | 58.96 | 8.66 | −5.66 | 10.35 | 7.88P | 5.79 | 2.61 | Purplish red |
| Lc28 | 60.32 | 8.62 | −4.55 | 9.75 | 9.40P | 5.93 | 2.49 | Purplish red |
| Lc29 | 56.35 | 9.56 | −8.93 | 13.08 | 5.80P | 5.53 | 3.23 | Purplish red |
| Lc30 | 61.29 | 9.99 | −2.51 | 10.30 | 2.77RP | 6.03 | 2.67 | Purplish red |
| Lc31 | 54.47 | 8.15 | −8.29 | 11.63 | 5.25P | 5.35 | 2.85 | Purplish red |
| Lc32 | 54.66 | 10.02 | −6.98 | 12.21 | 7.54P | 5.37 | 3.02 | Purplish red |
| Lc33 | 59.26 | 4.04 | −5.82 | 7.08 | 2.79P | 5.82 | 1.82 | Purplish red |
| Lc34 | 55.57 | 6.98 | −8.47 | 10.98 | 3.97P | 5.46 | 2.71 | Purplish red |
| Lc40 | 58.45 | 7.61 | −6.26 | 9.85 | 6.53P | 5.74 | 2.48 | Purplish red |
| Lc41 | 58.79 | 9.34 | −8.79 | 12.83 | 5.78P | 5.78 | 3.21 | Purplish red |
| Lc43 | 60.20 | 9.84 | −6.10 | 11.58 | 8.25P | 5.92 | 2.95 | Purplish red |
| Lc44 | 58.47 | 11.78 | −11.69 | 16.60 | 5.42P | 5.75 | 4.09 | Purplish red |
| Lc46 | 59.26 | 8.96 | −10.24 | 13.61 | 4.49P | 5.83 | 3.39 | Purplish red |
| Lc52 | 63.01 | 2.85 | −7.38 | 7.91 | 0.17P | 6.18 | 2.22 | Purplish red |
| Lc53 | 55.87 | 5.89 | −5.91 | 8.34 | 5.41P | 5.48 | 2.08 | Purplish red |
| Lc54 | 57.39 | 4.51 | −5.62 | 7.20 | 3.92P | 5.64 | 1.81 | Purplish red |
| Lc55 | 55.37 | 5.55 | −7.32 | 9.19 | 3.31P | 5.43 | 2.28 | Purplish red |
| Lc58 | 55.52 | 2.48 | −6.82 | 7.26 | 1.45P | 5.43 | 1.97 | Purplish red |
| Lc60 | 61.00 | 4.01 | −6.58 | 7.71 | 1.80P | 6 | 2.02 | Purplish red |
References
- Creech, J.L. On the distribution of Loropetalum chinense. Am. Hortic. Mag. 1960, 39, 236. [Google Scholar]
- Tang, Q.R. Study on the Genetic Diversity among Loropetalum chinense var. rubrum Introductions and Changes of Physiology and Bioehemistry during Its Leaf Colour Transformation. Ph.D. Dissertation, Hunan Agricultural University, Changsha, China, 2001. [Google Scholar]
- Huang, R.K.; Yang, C.K.; He, Z.H.; LI, J.C.; Liu, Y.Q. Resources investigation of Loropetalum chinense var. rubrum. Hunan Agric. Sci. 1998, 4, 44–45. [Google Scholar]
- Shao, Z.X.; Hou, W.; Long, X.Z.; Yang, G.D.; Chen, C.H.; Zeng, X.J.; Hou, B.X.; Yu, G.F.; Wu, W.W. The formation and development of the geography symbol product Loropetalum chinense var.rubrum in Liuyang City. Hunan For. Sci. Technol. 2007, 2, 71–73. [Google Scholar]
- Liang, S.L.; Tian, H. “Panda in plants”-Loropetalum chinense var. rubrum. China Flower Hortic. 2003, 22, 40. [Google Scholar]
- Hou, B.X. The development and prospect of the characteristic flower industry of Loropetalum chinense var. rubrum. Technol. Mark. (Landsc. Eng.) 2006, 11, 40–43. [Google Scholar]
- Gawel, N.J.; Johnson, G.R.; Sauve, R. Identification of genetic diversity among Loropetalum chinense var. rubrum introductions. J. Environ. Hortic. 1996, 14, 38–41. [Google Scholar] [CrossRef]
- Wang, W.T. Preliminary study on annual cultivation and conservation of Loropetalum chinense var. rubrum. Hunan For. Sci. Technol. 2004, 31, 67–68. [Google Scholar]
- Hou, B.X.; Yu, G.F.; Song, Q.G.; Yi, A.Q. Cultivation and management techniques of Loropetalum chinense var. rubrum. Hunan For. Sci. Technol. 2003, 1, 34–35. [Google Scholar]
- Li, H.W.; Duan, Z.G.; Feng, J.T.; Liu, J.J. Characteristics and cultivation and management techniques of Loropetalum chinense var. rubrum. Agric. Dev. Equip. 2017, 8, 185. [Google Scholar]
- Tang, Q.R.; Chen, D.F.; Chen, Y.Y.; Zhang, H.Z.; Zhou, P.H. Changes of physiology and biochemistry during leaf color transformation in Loropetalum chinense var. rubrum. Sci. Silvae Sin. 2006, 2, 111–115. [Google Scholar]
- Li, Y.L. Studies on Genetic and Biological Characteristics of Flower Leaf Bud of Loropetalum chinense var. Rubrum. Master’s Thesis, Hunan Agricultural University, Changsha, China, 2009. [Google Scholar]
- Chen, Q.R.; Cai, W.Q.; Zhang, X.; Zhang, D.M.; Li, W.D.; Xu, L.; Yu, X.Y. The comparative studies on phytochemicals of leaf coloration of Loropetalum chinense var. rubrum. Acta Hortic. Sin. 2021, 48, 1969–1982. [Google Scholar]
- Chen, X.L.; Chen, J.H.; Hou, B.X.; Chen, G. Comparative analysis on photosynthetic characteristics of Loropetalum chinense var. rubrum. J. Cent. South Univ. For. Technol. 2010, 30, 119–121. [Google Scholar]
- Zhang, Z.Z. Photosynthetic physiological characteristics of Loropetalum chinense var. rubrum and its application in landscape. Qinghai Sci. Technol. Agric. For. 2011, 2, 17–19. [Google Scholar]
- Liu, J.; Zhang, L.; Li, W.D.; Li, T.; Li, Y.L.; Yu, X.Y. Comparative analysis of photosynthetic physiological characteristics of several Loropetalum chinense lines with different flower colors. Non Wood For. Res. 2020, 38, 245–251. [Google Scholar]
- Wani, G.A.; Shah, M.A.; Tekeu, H.; Reshi, Z.A.; Atangana, A.R.; Khasa, D.P. Phenotypic variability and genetic diversity of phragmites australis in Quebec and Kashmir reveal contrasting population Structure. Plants 2020, 9, 1392. [Google Scholar] [CrossRef] [PubMed]
- Bigard, A.; Berhe, D.T.; Maoddi, E.; Sire, Y.; Boursiquot, J.M.; Ojeda, H.; Peros, J.P.; Doligez, A.; Romieu, C.; Torregrosa, L. Vitis vinifera L. fruit diversity to breed varieties anticipating climate changes. Front. Plant Sci. 2018, 9, 455. [Google Scholar] [CrossRef]
- Manco, R.; Basile, B.; Capuozzo, C.; Scognamiglio, P.; Forlani, M.; Rao, R.; Corrado, G. Molecular and phenotypic diversity of traditional european plum (Prunus domestica L.) germplasm of southern Italy. Sustainability 2019, 11, 4112. [Google Scholar] [CrossRef]
- Wei, G.M.; Wang, S.M.; Luo, L.S.; Leng, J.G.; Hou, Y.H. Phenotypic trait diversity of acorus calamus germplasm resources. Guizhou Agric. Sci. 2002, 50, 15–20. [Google Scholar]
- Zhang, B.B.; Cai, Z.X.; Shen, Z.J.; Yan, J.; Ma, R.J.; Yu, M.L. Diversity Analysis of phenotypic characters in germplasm resources of ornamental peaches. Sci. Agric. Sin. 2021, 54, 2406–2418. [Google Scholar]
- Duan, J.J.; Ling, Z.; Jia, M.L.; Song, Z.Q.; Zhang, C.; Cao, D.M. Phenotypic diversity analysis of wild resources of hemerocallis in Shanxi Province. J. Plant Resour. Environ. 2021, 30, 29–38. [Google Scholar]
- Chen, J.Y. Ornamental Plants (Diversity); Chen, J.Y., Ed.; Beijing Forestry University, Chinese Society of Horticulture: Beijing, China, Selected Essays by Professor Chen Junyu; China Agricultural Science and Technology Press: Beijing, China, 1997; pp. 73–77. [Google Scholar]
- Wang, J.; Xu, L.F.; Wang, L.; Qi, X.Y.; Song, M.; Cao, Y.W.; He, G.R.; Tang, Y.C.; Yang, P.P.; Ming, J. The numerical classification of flower color phenotype in lily. Acta Hortic. Sin. 2022, 49, 571–580. [Google Scholar]
- Liu, Y.P.; Wu, F.F.; He, D.; Zhuang, Y.; Kong, D.Z. Numerical classification of lotus cultivars based on flower color phenotype. J. Zhejiang Univ. (Agric. Life Sci.) 2020, 46, 319–326. [Google Scholar]
- Wang, C.; Rong, L.Q.; Feng, L.J.; Liu, H.Q. Preliminary study of mechanism on diversity of floral organ variants in Anemone rivularis var. flore-minore. Acta Hortic. Sin. 2017, 44, 89–96. [Google Scholar]
- Hou, B.X.; Lin, F.; Yi, A.Q.; Yu, G.F.; Wang, X.M.; Song, Q.A. Study of the dissected structure of Loropetalum chinense and Loropetalum chinense var. Rubrum. Chin. Wild Plant Resour. 2004, 23, 47–49. [Google Scholar]
- Wang, Y. Research progress of Loropetalum chinense var. rubrum germplasm resources and their genetic characteristics. J. Hunan Univ. Arts Sci. (Nat. Sci. Ed.) 2007, 19, 66–68. [Google Scholar]
- Hou, B.X.; Lin, F.; Yu, G.F.; Yi, A.Q. Study of the number variation of outward characteristic of the Loropetalum chinense and Loropetalum chinense var. rubrum. J. Plant Genet. Resour. 2003, 4, 203–206. [Google Scholar]
- Hou, B.X.; Lin, F.; Li, W.P.; Wang, X.M.; Yu, G.F.; Song, Q.A. Classification system of Loropetalum chinense var. rubrum. For. Res. 2003, 16, 430–433. [Google Scholar]
- Gonnet, J.F. CLElab measurement, a precise communication in flower colour: An example with carnation (dianthus caryophyllus) cultivars. J. Hortic. Sci. 1993, 68, 499–510. [Google Scholar] [CrossRef]
- Sun, W.; Li, C.H.; Wang, L.S.; Dai, S.L. Study on the determination part of flower color of chrysanthemum tongue. J. Hortic. 2010, 37, 777–784. [Google Scholar]
- Wang, F.; Yang, S.H.; Liu, X.Y.; Cui, J.P.; Chang, Z.H.; Ge, H. Flower color diversity of rose germplasm resources and its relationship with anthocyanins. J. Hortic. 2017, 44, 1125–1134. [Google Scholar]
- Guo, X.; Cheng, F.Y.; Zhong, Y.; Chen, X.Y.; Tao, X.W. The quantitative classification of flower color phenotype in Paeonia rockii (flare tree peony). Acta Hortic. Sin. 2022, 49, 86–99. [Google Scholar]
- Yin, H.T.; Yin, J.M.; Liao, Y.; Lu, S.J.; Li, C.H. Phenotype classification based on flower color, pigment distribution and epidermal cell shape of Dendrobium hybrids. Acta Hortic. Sin. 2021, 48, 1907–1920. [Google Scholar]
- Hong, Y.; Bai, X.X.; Sun, W.; Jia, F.W.; Dai, S.L. Quantitative classification of flower color phenotypes of chrysanthemum cultivars. J. Hortic. 2012, 39, 1330–1340. [Google Scholar]
- Li, Y.Q.; Liu, X.Y.; Guo, X.Y.; Pan, J.S.; Tang, D.Q. Analyses of flower color phenotype and flower pigments of Petunia hybrida new germplasms. Subtrop. Plant Sci. 2021, 50, 378–387. [Google Scholar]
- Jiang, X.K. Studies on Test Guideline to New Vaarities on Distinctness Uniformity and Stability of Amorphophallus. Master’s Thesis, Southwest University, Chongqing, China, 2012. [Google Scholar]
- Ren, X.Y. Study on Mutation and Identification of Hibiscus Syriacus Seeds by Colchicine and EMS. Master’s Thesis, Central South University of Forestry & Technology, Changsha, China, 2019. [Google Scholar]
- Zhao, B.T.; Zhao, W.; Liu, L.W. Advances in the analytical methods of anthocyanin com pounds. J. Northwest A F Univ. (Nat. Sci. Ed.) 2014, 42, 180–188. [Google Scholar]
- Romero, I.; Sanchez-Ballesta, M.T.; Maldonado, R.; Isabel escribano, M.; Merodio, C. Anthocyanin, antioxidant activity and stress-induced gene expression in high CO2-treated table grapes stored at tow temperature. J. Plant Physiol. 2008, 165, 522–530. [Google Scholar] [CrossRef]
- Wrolstad, R.; Culbertson, J.; Cornwell, C.; Mattick, L. Detection of adulteration in blackberry juice concentrates and wines. Assoc. Off. Anal. Chem. 1982, 65, 1417–1423. [Google Scholar] [CrossRef]
- Su, Q.; Yang, Y.H.; Tian, M.; Zhang, J.Z.; Mao, L.Y.; Tang, Y.W. Bu, Z.Y.; Lu, Z.S. Phenotypic diversity analysis and comprehensive evaluation of 49 waterlily resources. Southwest China J. Agric. Sci. 2019, 32, 2670–2681. [Google Scholar]
- Petruccelli, R.; Ganino, T.; Ciaccheri, L.; Maselli, F.; Mariotti, P. Phenotypic diversity of traditional cherry Accessions present in the tuscan region. Sci. Hortic. 2013, 150, 334–347. [Google Scholar] [CrossRef]
- Lv, W.; Han, J.N.; Wen, F.; Ren, G.X.; Wang, R.P.; Liu, W.P. Phenotypic Diversity analysis of sesame germplasm resources. J. Plant Genet. Resour. 2020, 21, 234–242, 251. [Google Scholar]
- Wang, L.F.; Xu, J.J.; Huang, X.X.; Li, S.B.; Cheng, X.M. Phenotypic Traits and comprehensive evaluation of 57 modern Chinese rose germplasm resources. J. Southwest For. Univ. 2022, 42, 83–90. [Google Scholar]
- Yuan, X. Study on Genetic Diversity of Phenotype Traits of Lotus Germplasm Resources and Classification Based on Flower Color Phenotype. Master’s Thesis, Henan Agricultural University, Changsha, China, 2021. [Google Scholar]
- Li, Y.; Zhang, S.H.; Guo, Y.; Zhang, X.F.; Wang, G.P. Catkin phenotypic diversity and cluster analysis of 211 Chinese chestnut germplasms. Sci. Agric. Sin. 2020, 53, 4667–4682. [Google Scholar]
- Rui, W.J.; Wang, X.M.; Zhang, Q.N.; Hu, X.Y.; Hu, X.H.; Fu, X.J.; Gao, Y.M.; Li, J.S. Genetic diversity analysis of 353 tomato germplasm resources by phenotypic traits. Acta Hortic. Sin. 2018, 45, 561–570. [Google Scholar]
- Tucker, A.O.; Maciarello, M.J. A survey of colored harts for biological description. Taxon 1991, 40, 201–214. [Google Scholar] [CrossRef]
- Bao, Z.Y.; Zhang, H.; Bao, J.S.; Huang, Z. Genetic diversity of Loropetalum chinense var. rubrum was analyzed by RAPD. J. Zhejiang Univ. (Agric. Life Sci.) 2003, 29, 665–670. [Google Scholar]
- Bi, M.M.; Cao, Y.W.; Song, M.; Tang, Y.C.; He, G.R.; Yang, Y.; Yang, P.P.; Xu, L.F.; Ming, J. Advances in flower color of Lilium. Acta Hortic. Sin. 2021, 48, 2073–2086. [Google Scholar]
- Yu, P.C.; Tan, P.Y.; Gao, L.; Jia, G.X. Analysis of the development of flower color based on hybridization breeding process in Lilium OT hybrids. Acta Hortic. Sin. 2021, 48, 1885–1894. [Google Scholar]
- Zou, H.Z.; Zhou, L.; Han, L.L.; Lv, J.H.; Wang, Y. Changes of carotenoid components and expression of the related genes during petal coloring of Paeonia delavayi. Acta Hortic. Sin. 2021, 48, 1934–1944. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).