Abstract
Curculio glandium females associated with the pedunculate oak were investigated in order to fill the knowledge gap on acorn structure preferences and shell-hardening influence on ovipositional behavior and fecundity. Shell solidification progression of weekly harvested acorns was measured using penetration and force–deformation tests along with fruit mass within a time frame covering weevils’ reproduction period. Captured females were offered regular acorns (uncracked, soft enough to drill into) and older seeds (with natural cracks) for behavior recordings and preferences tests. Young acorns and fruits ripe enough to be too hard for females to drill (artificially pierced and untouched) were used for egg output assessment throughout the shell-hardening progression. Experiments revealed that naturally cracked acorns were chosen significantly more often, which resulted in reduced drilling-phase duration. Egg number did not differ significantly before and after the threshold of acorn hardening; however, having passed it, females significantly more frequently deposited eggs in artificially pierced seeds. Results indicate the opportunistic nature of female preferences. Oviposition in cracked or pierced acorns facilitates the process and lowers competition for relatively shortly available soft and healthy seeds, the first of which reach impenetrability in the third week of August.
Keywords:
ethogram; force test; phenology; Palaearctic; plant–insect interactions; opportunistic behavior; Poland; Quercus; shell 1. Introduction
Curculio glandium Marsham (Coleoptera: Curculionidae: Curculionini) is a carpophagus weevil associated with a variety of oak species (Quercus spp.) that is widely distributed in Europe [1,2,3,4,5]. In the central part of the continent, its main host plant is the pedunculate oak (Q. robur L., Fagaceae), the seeds of which are shelter and a food supply for eggs and larvae, respectively. This weevil species is distinct among Curculionidae for possessing proportionally longer snouts (rostrum) [6], which act as the females’ drill used for the creation of egg galleries. The very process of drilling and its mechanical implications on the rostrum were elaborated by Matsumura et al. [7].
Despite its frequent occurrence and importance as an oak pest, some reproductive aspects of C. glandium ecology seem not to be entirely recognized yet. This is somewhat peculiar inasmuch as the acorn weevil is capable of reducing germination rate by up to 70–90%, mainly by cotyledon destruction [8,9]. In Europe, the species coexists with other pre-dispersal seed parasites, namely C. pellitus (Boheman), C. venosus (Gravenhorst) and C. elephas (Gyllenhal), also limiting acorn production [10,11]. Our recent studies revealed that when C. glandium co-occurs with other Curculio spp. and competes for limited resources, there is a certain degree of phenological inter-specific segregation. Curculio glandium starts oviposition relatively late (i.e., starting in the third week of July) and females drill and lay eggs into considerably grown (but not fully matured) acorns. Furthermore, the discussed species turned out to be the only Curculio weevil known so far to create radial galleries (harboring significantly higher numbers of eggs, compared to co-occurring C. venosus and C. pellitus) instead of simple egg holes [12]. The most recently proven peculiarity of C. glandium reproduction is females’ ability to oviposit, also into fallen acorns with natural cracks. Fractures allow for prolonged (up to mid-September) egg deposition in acorns which might have been already too hard for females to drill in a regular manner. This, with the highest probability, should be considered an adaptation to unfavorable environmental conditions [13], such as variable production of regularly infested, young acorns [14,15,16,17].
Seed shell solidification is one of the innate mechanisms developed by some plants as a means of resistance against invertebrate herbivores [18,19]. A phenology-focused relationship between the natural seed hardening and reproduction of curculionids has been examined in the case of C. nucum L. associated with hazelnut Corylus avellana L. varieties differentiated in terms of seed-ripening period. The research performed in Italy indicated that the vulnerability of nuts to nut weevil parasitism was strictly related to the shell-hardening timing—the more rapid the hardening, the more hampered the oviposition of C. nucum females; however, no relation was found to the shell thickness [20]. Analogous results were obtained during research conducted on hickory (Carya illinoinensis (Wangenh.) K. Koch) infested by the pecan weevil C. caryae (Horn)—the early maturing cultivar was damaged significantly less than the later maturing ones [21]. Other observations on C. caryae as well as on the filbert weevil C. occidentis L. also indicated that females did not oviposit in fully mature fruits of pecans and the Garry oak (Q. garryana Douglas ex Hook.), respectively [17,22]. In the case of C. glandium, no data on acorn shell hardness’ effect on reproduction are available, despite the fact that oak fruits are also nutritious food source for humans [23,24].
The above-cited results, in the context of C. glandium females’ tendency to drill highly time-consuming, radial tunnels in already quite grown acorns combined with their capability of also exploiting fallen, ripe fruits with fractures, prompted us to look at: (i) How is natural acorn hardening related to reproduction of acorn weevils? (ii) When exactly are the pedunculate oak fruits too hard to be drilled into? (iii) How does hardening affect ovipositional behavior, acorn preferences and reproductive capacity of females? Based on what we already observed in nature, we hypothesize that females are likely to seize the opportunity to shorten the drilling phase and to deposit eggs in naturally cracked and/or artificially pierced acorns.
Laboratory experiments performed within this research were designed to answer the above questions by 1. estimating natural hardness progression accompanying acorn growth; 2. providing a detailed description of females’ ovipositional activities and their duration depending on acorn condition; 3. learning about females’ preferences for cracked or uncracked acorns as ovipositional sites; and 4. evaluating egg output and fecundity change during the season in females offered three types of acorns, i.e., typically utilized—possible to drill into (I); too hard for effective drilling—untouched (II); and fractured (i.e., naturally cracked/artificially pierced) (III).
2. Material and Methods
2.1. Experiment 1. Acorn Development and Hardness Change during Reproduction of C. glandium
2.1.1. Differences along the Season
In order to analyze hardening progression during oak fruit development, we performed a series of acorn harvests. Fifteen undamaged fruits originating from the same tree were collected once a week from 23 July to 3 September. The period is the time frame covering C. glandium reproduction in Poland, which usually starts in the last week of July and ends in early September [12]. Acorns were picked up early in the morning, measured and then proceeded to further analysis in the Laboratory of Rheological Measurements (Institute of Food Sciences, Warsaw University of Life Sciences—SGGW). The hardness of each acorn was examined using a TA-HD plus texture analyzer equipped with a 5 kg load cell (Stable Micro Systems, Surrey, UK). The acorn was placed on the heavy-duty platform (HDP/90) and subjected to a penetration test using a needle probe (P/2N) with constant speed of 1 mm·s−1 and penetration distance of up to 5 mm. The force–deformation tests were carried out along the Z-axis of acorns (Figure 1) and the following parameters were extracted from the penetration curve: fracture force (N) and deformation (mm) of acorn shell (Force 1, Distance 1) and force at penetration distance of 5 mm (N, Force 2). Applied forces caused both shell and cotyledon penetration and deformation.
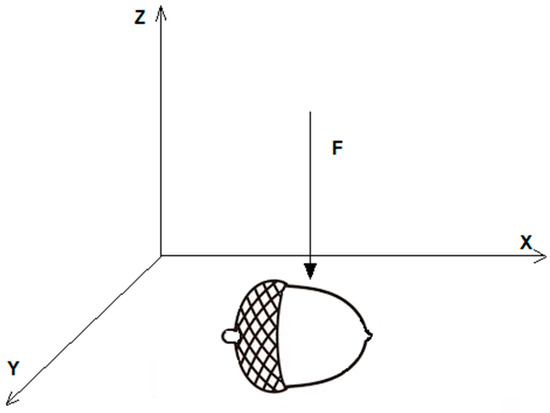
Figure 1.
The position of an acorn and penetration probe (P/2N) during force–deformation test.
Additionally, the mass of each acorn was determined using an electronic balance with 0.01 g sensitivity.
2.1.2. Mother Tree Differences
Additionally, at a point close to acorn growth termination (27th of August), we collected 15 acorns from three different oak trees growing in the same area to see if there were any physical differences in fruits between particular oaks. The applied measuring procedure was as described above (Section 2.1.1).
2.2. Experiment 2. Female Ovipositional Behavior on Acorns
Individuals of C. glandium to be used in video recordings were collected from oaks (>60 years old) growing in a University campus park in Warsaw, Poland. Species identity was confirmed based on the key by Smreczyński [25]. Weevils were kept in same-sex groups in plastic containers for 48 h prior to the experiment, and fed on an apple. In order to make females more motivated for oviposition during the experiment, no acorns were given at that stage. Then, on the day of the experiment, insects were placed in plastic containers with access to acorns and apple pieces. The acorns provided were checked beforehand for the presence of markings (i.e., punctures, discoloration) suggesting that they had already been utilized by other weevil females or moths.
Video recordings took place under laboratory conditions with no sunlight, but in a controlled photoperiod (adjusted to actual conditions on a particular day) and stable temperature (c. 21 °C). Recordings were made for 48 h after weevils were released, including during the night (when the red light was turned on). We used two IP (internet protocol) cameras (Day/Night megapixel IP camera, Novus, Poland) connected with a computer and an external disc. All of the recordings were analyzed manually by watching each video. Footage was stored on an external drive, so we were able to watch them multiple times if needed. Experimental arenas were made of transparent plastic. Video registrations were set to capture the behavior of individual female in optimal conditions: without other competing females or disruptive males. In this configuration, females could freely present ovipositional behavior: drilling egg channels and egg deposition. In order to assess whether the presence of cracks in the acorn shell affected oviposition behavior and duration, some of the females (n = 30) were allowed to exploit young, undestroyed acorns, while others (n = 30) had access to older and harder but cracked fruits.
While analyzing behavior, the following categories were under consideration and their durations (in minutes) registered: (1) initial climbing on acorn, (2) exploration, (3) biting the surface of acorn, (4) creating egg channel (shallow and deep drilling), (5) locating ovipositor in egg channel, (6) egg deposition: successful—eggs probably laid; unsuccessful—eggs probably not laid, problems with ovipositor or egg placed on acorn surface, (7) walking around, (8) leaving the acorn. Further on, an ethogram was prepared.
2.3. Experiment 3. Female Choice Test—Cracked vs. Uncracked Acorns
Forty-five females placed singly into plastic containers with apple piece as a food source were offered two acorns, naturally cracked and uncracked, to drill in and deposit eggs. After 24 h, fruits were removed and dissected with a surgical blade in order to check which one was chosen. Selected acorns were determined by the presence of eggs in egg galleries.
2.4. Experiment 4. Changes in Egg Output during the Season and Female Fecundity
Selected females (n = 12) were kept individually in plastic containers with constant access to food (apple). Every day, starting from 27th of July (when weevils mate in the natural conditions) to 17th of September (with modification explained later on), females were offered two acorns of the same type (i.e., with no visible signs of occupation by other weevils or Cydia moths). Acorns used were fresh and were collected just before offering to females; moreover, they were picked straight from a cohort of 10 trees to avoid tree specificity bias. Each acorn represented a growth phase which females would encounter if they foraged in natural conditions. After 24 h, utilized fruits were removed and new ones provided. In the third week of August, we realized that females stopped laying eggs—most likely because the acorns became too hard to be drilled into. Thus, we facilitated oviposition continuation. Starting from 23 August (the date we adopted as the moment that acorns hardened, determined based on two-day oviposition cessation), one of the two offered acorns was pierced with a minute entomological needle, resulting in a hole resembling a natural egg channel entrance.
Every day, all of the utilized fruits were dissected and the number of eggs was noted. During the weekends (i.e., Saturdays and Sundays), each female was provided with a male as company to ensure the production of new eggs.
2.5. Statistical Analysis
Results of the four experiments were statistically processed using Statistica software (v. 13.0, StatSoft Inc., Tulsa, OK, USA). Normality of distributions (Shapiro–Wilk test, α = 0.05) and homogeneity of variance (Levene’s test, α = 0.05) have been checked in all data obtained (experiments 1–4). Consequently:
- a parametric, one-way ANOVA with a post-hoc Tukey’s HSD test was used to analyze significant differences (p < 0.05) in hardness and masses of acorns; the effects of oak fruit development during the season and mother tree differences on four variables (Force 1, Force 2, Deformation 1, Mass) were checked (Experiment 1);
- a non-parametric Mann–Whitney U-test was used to determine significant differences (p < 0.05) in the amount of time spent on creating egg channels in cracked and uncracked acorns (Experiment 2);
- a non-parametric Cochran Q test was used to settle significant differences (p < 0.05) in the choice of cracked (coded as 0) or uncracked (coded as 1) acorns as ovipositional sites (Experiment 3);
- a non-parametric Kruskal–Wallis ANOVA with a post-hoc Dunn’s test was performed to evaluate whether females differed significantly (p < 0.05) in reproductive capacity from 29 July to 20 August and, after the hardening of acorns, from 23 August to 17 September (Experiment 4);
- a non-parametric Mann–Whitney U-test was applied to assess the significant difference (p < 0.05) in the number of eggs laid into two typical acorns given to females prior to fruit hardening, as well as in the number of eggs laid in pierced or unpierced fruits after hardening (Experiment 4).
3. Results
3.1. Experiment 1. Acorn Development and Hardness Change during Reproduction of C. glandium
The hardness of each acorn’s shell and cotyledon was determined using the penetration test. Figure 2 presents a typical force–deformation curve recorded during the mechanical test. The first peak on the curve indicates the puncture of the shell. The values of force and deformation (Force 1, Deformation 1) recorded at this point indicate the shell hardness. The second maximum on the penetration curve (Force 2) is related to the hardness of acorn cotyledon.
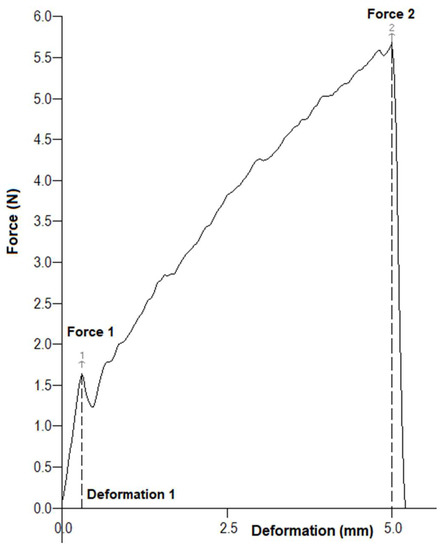
Figure 2.
The force–deformation curve of an acorn: Force 1 and Deformation 1—force and deformation of material at the moment of penetration of the shell, Force 2—force recorded at deformation of 5 mm.
The changes in shell hardness (Force 1) of acorns harvested from the 23th of July to the 3rd of September are presented in Figure 3a. In this period, the gradual increase in shell hardness, from 1.85 to 2.87 N, was observed. The shell hardness did not differ for acorns harvested on 30th of July and 6th of August as well as for the samples investigated on 20th and 27th of August (F(6) = 144.66, p < 0.0001). However, the significant threshold occurred between 13th and 20th of August (92.2% of total hardness) (F(6) = 233.6, p < 0.0001). During the first 21 days of the experiment, the hardness of acorn shells increased by about 30% and the mean mass increased by more than twice in the same period. The next 21 days of the oak nuts’ maturation resulted in an increase in shell hardness and acorn mass only by about 18% and 29%, respectively (Figure 3a).
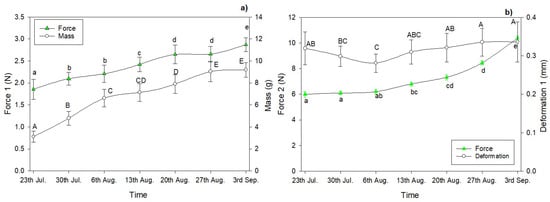
Figure 3.
Changes in shell hardness (Force 1, Force 2), deformation at fracture (Deformation 1) and average mass of acorns harvested from 23 July to 3 September: different letters indicate significant differences (Tukey’s HSD, p < 0.05) in shell hardness—Force 1 (a) or Force 2 (b); different capital letters indicate significant differences (Tukey’s HSD, p < 0.05) between masses (a) or Deformation 1 (b) of acorns.
Figure 3b shows values of the deformation required for the acorn shell to fracture (Deformation 1). This mechanical parameter (referred to as strain) describes the resistance of material to deformation. Slight fluctuations in the values of deformation 1 at the puncture of the shell were observed.
The force obtained at constant deformation of acorns (penetration depth 5 mm, Force 2) also increased during the investigated period of maturation but the rate of the changes was different than observed for acorn shells (Figure 3b). The hardness of cotyledon significantly determines the value of penetration by Force 2. This parameter increased by about 13% during the first 21 days of the experiment (from 23th of July to the 13th of August). However, the hardness of acorn cotyledon increased by about 50% in the period from 13th of August to 3rd of September (the next 21 days of maturation).
Results of force–deformation tests of acorns harvested on 27 August from three different oak trees showed significant differentiation of plant material (F(2) = 458.3, p < 0.0001). Hardness of the acorn shell varied from 1.87 to 2.65 N but the deformation at the puncture site of the acorn shell did not differ between trees (F(2) = 0.43, p = 0.651). However, the mean mass of acorns (3.00–9.02 g) varied significantly between the trees (F(2) = 607.1, p < 0.0001) (Table 1).

Table 1.
Selected mechanical parameters and an average mass of acorns from three oak trees.
3.2. Experiment 2. Female Ovipositional Behavior on Acorns
Based on recordings, an ethogram of C. glandium females’ oviposition activities was created (Table 2 and Table 3, Figure 4). Initially, when the acorn is placed in the arena, a female explores its surface. As she chooses a spot, usually just under the cupula, she slowly starts chewing through the external layer. After breaking it, the drilling starts. The first phase (i.e., shallow drilling) is characterized by various movements around the hole. The female does not insert the whole rostrum, but only up to half of its length. At this stage, females often stop drilling and abandon acorns if disturbed. This part can last more than one hour, but with cracked acorns, it is less time-consuming.
Table 2.
Ethogram of C. glandium female during oviposition.
Table 3.
Data on duration (minutes) of particular events during oviposition by C. glandium females offered cracked and uncracked oak fruits.
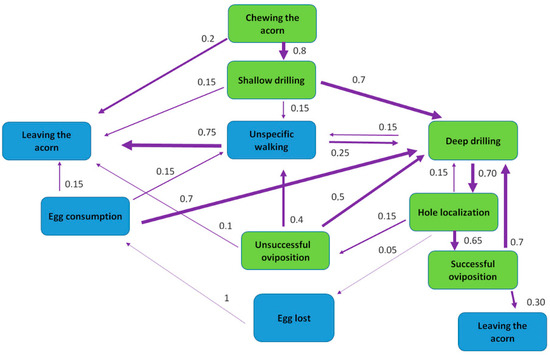
Figure 4.
Flow chart scheme representing the sequence of a typical female behavior during oviposition (n = 27): green color—ovipositional activities, blue—non-related behavior; arrow thickness indicates frequency of behavior; numbers stand for the probability of turning one behavior into another.
The second phase of drilling (i.e., deep drilling) is characterized by entering almost the entire snout into the egg channel. Deep drilling is performed in one position, which is rather still. In total, the drilling process can last up to 197 min or can be much shorter (2 min) when the acorn is cracked (Table 3, Figure 4). When the egg channel is ready, the female turns 180° and tries to locate its entrance with an ovipositor. It usually takes a few seconds, but many times we observed problems with the process. If a female was not able to find the hole, they may have given up on oviposition or gone back to drilling again. In a few cases (c. 5% of total ovipositions), we saw females placing an egg on the acorn surface and consuming it afterwards. If the ovipositor was correctly placed inside the fruit, the female stood still for egg laying, which usually took around 40 s. After the whole process was completed, females usually did not leave the acorn but continued drilling.
At the beginning of the breeding season, when acorns are abundant and accessible (soft enough to drill into), females place up to four eggs into one acorn. Sometimes they leave the acorn after the second oviposition. As the season develops, acorns become harder and mostly impossible for females to drill. This leads to more intense competition events. Therefore, females probably deposit more eggs into the same acorn (M.R.: personal observations). Detailed durations of events during oviposition are shown in Table 3. The analysis of the obtained results showed a significant difference (p < 0.05) in time taken to create channels in uncracked and cracked acorns (U(n1 = 30, n2 = 30) = 3.00, Z = 6.60, p < 0.001). Females spent significantly more time drilling channels in the uncracked acorns vs. cracked ones (Figure 5).
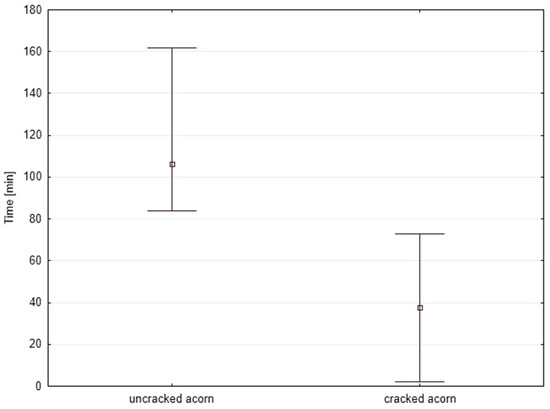
Figure 5.
Time spent by females on drilling egg channels in uncracked vs. cracked acorns: squares—average time (median), whiskers—minimal and maximal duration.
3.3. Experiment 3. Female Choice Test—Cracked vs. Uncracked Acorns
Statistical analysis showed that females (n = 45) significantly more often chose cracked (93%) rather than uncracked acorns (7%) (42 vs. 3 cases) for oviposition (Q (1) = 33.8, p < 0.001).
3.4. Experiment 4. Changes in Egg Output during the Season and Female Fecundity
The sum total of eggs deposited by 12 females throughout 51 days of the experiment ranged from 48 to 75, with a mean of 64.8 and a standard deviation of 7.7.
The number of eggs deposited by females before acorns hardened did not differ significantly between the two offered acorns (U(n1 = 204, n2 = 204) = 2046, Z = −0.3179, p = 0.751). However, after acorns became too hard for females to effectively drill into (green field on the graph), significantly more females laid eggs in artificially pierced acorns than in the untouched, hard ones (U(n1 = 240, n2 = 240) = 8338, Z = 13.47, p < 0.001). No significant difference (p > 0.05) was found in the number of eggs (daily fecundity) laid by females in both periods, i.e., before fruit hardening on August 23rd (0–3 eggs) (H(11) = 9.10, p = 0.612) and after (0–4 eggs) (H(11) = 4.90, p = 0.936) (Figure 6).
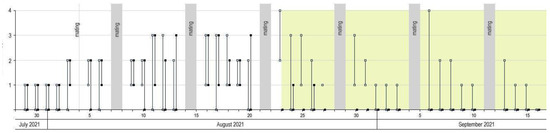
Figure 6.
Acorn selection and daily egg output range (min–max) of C. glandium females tested during Experiment 4. White background—pre-hardening period: empty squares—acorn one, filled squares—acorn two; green background—post-hardening period: empty squares—pierced hard acorns, filled squares—unpierced hard acorns.
4. Discussion
Acorns are still only studied in a limited capacity in many aspects despite being a human food source known for their nutritional value and very low levels of toxic contaminants [23,24]. Here, we present the results that can give an insight into acorns’ growth dynamics and the consequences for weevils relying on them to complete the life cycle. These fruits, while growing, not only grow in size and mass, but their thickness also increases, thereby the hardness, too. The latter seems to be a crucial factor for acorn-dependent invertebrates, mainly seed predators. Curculio glandium, the weevil focused on in this contribution, is an example of a species that adjusts its strategy depending on changing situations. Females of this beetle have a short time to drill and lay eggs inside acorns, and as days go by, there are less and less suitable resources.
The first experiment answered the question of how acorn shell hardness progressed during growth. To the best of our knowledge, this kind of data has not been available in the literature so far. Force tests and mass measurement results indicated that acorn hardness and weight advanced rather similarly in time; still, thresholds of significant changes were not entirely at the same points (Figure 3a). However, force values required to rupture the shell (hardness of shell) were positively correlated with acorn mass (Pearson’s correlation coefficient: r = 0.867). This is in agreement with results obtained by Valentini et al. [26] for hazelnuts during compression of nuts along the y-axis. Further, Koyuncu et al. [27] observed that the fracture force of walnut increased with shell thickness, nut weight and volume of the kernel. Our results indicated that the hardness of the shell and cotyledon were not linear during an acorns’ growth as measured in seven-day intervals (Figure 3a,b). The increase in shell hardness at different rates during nut growth was noted by Valentini et al. [19] too. Authors reported that at the beginning of the nut growth, the shell was soft and penetration force values reached 3–6 N, depending on the cultivar. Rapid increases in the force up to 20–40 N were observed when nuts reached 80% of their final size. The hardening process of the nut’s shell depends on the stage of growth and the cultivar of the nut. Guide et al. [20] observed that force values during the puncture test of the nut increased from 2.52 to 42.66 N for TGL variety, while for the Daria cultivar, they ranged from 2.52 to 3.69 in the harvest period from 21 June to 5 July.
The most important point, in the context of C. glandium reproduction, is the fact that oviposition cessation point (23 August) followed a significant change in oak fruit hardness. The threshold level of hardness was 92.2% of the maximal hardness and occurred between days 21 and 28 (20 August) of weekly measurements (Figure 3 and Figure 5). Therefore, a conclusion can be drawn that in the study area, the third week of August is the point at which acorn resources naturally start to shrink and some females may be forced to search for alternatives, i.e., fruits with damaged shells.
The second experiment was an attempt to describe in detail the ovipositional behavior of C. glandium on whole and cracked oak fruits. According to video footage, females’ strategies change during the reproductive period and can be seen as a result of alternating physical properties of acorns. At the beginning of the egg-laying term (end of July in central Poland), females have access to young, still growing, and soft acorns. In such fruits, weevils are capable of creating egg tunnels with their elongated rostrum. Such behavior is rather time-consuming, as it usually takes, on average, 107 min until the egg is laid (Table 3). For comparison, Eucryptorrhynchus brandti Harold, an Asian weevil reproducing in solid materials (trunk of tree Ailanthus altissima (Mill.) Swingle (Simaroubaceae)), excavates the egg cavity for around 20 min [28]; however, the species does not drill channels of radial pattern, which is a considerable distinction, reflected in the duration differentiation.
Additionally, if an initial egg output is rather low (one or two eggs per fruit), then females leave the acorn and search for another one. Thus, even though weevils spend a lot of time drilling, a low number of eggs are deposited. This might suggest that females try to infest as many fruits as possible, most likely to avoid competition within their own offspring. As the season progresses and acorns harden, females have less resources for egg deposition. In such situations, they seem to search for acorns with shells somehow weakened or damaged. The latter are usually easily accessible, as some oak fruits crack naturally while growing [29]. This allows females to lay eggs into normally unavailable, too hard fruits. Video recordings indicate that exploiting cracked acorns allows for saving a considerable amount of time; excavation in cracked acorns takes 41 min on average, which is 2.6 times faster than in the case of whole seeds (Table 3).
In the third experiment, we aimed at learning females’ preferences for oak fruits: would they prefer undamaged fresh acorns over the damaged older ones? The significant majority of females (42 of 45) chose cracked acorns, which can be seen as typical opportunistic behavior. Additionally, this preference was confirmed by our previous observations made in the natural environment, when C. glandium females oviposited on fallen acorns (mostly not matured, abscissed ones) until mid-September [13]. Opportunistic behavior might be characteristic for C. glandium solely, as other species coexisting on oaks (C. venosus and C. pellitus) in Poland have not been observed using naturally cracked acorns for oviposition [12]. Additionally, C. elephas does not seem to exploit this niche too, even if it faces similar resource issues (M.R.: personal observation, Raul Bonal: personal communication). It is unknown if acorn weevil females detect any chemical compounds to determine the fruit quality; it is also unknown if they release any semiochemicals that could attract other females.
Finally, the last experiment let us know how progressively hardening acorns influenced the oviposition process and egg output (Figure 6). According to our observations, acorns became too hard for females to effectively drill in the third week of August, when the oviposition ceased. We adopted the date of 23rd August as the acorn-hardening threshold. Before that date, Mann–Whitney’s U-test revealed no difference in the number of eggs laid in two typical, soft acorns offered to females. After the 23 August, however, a significant majority of females gave up trying to drill into too hard acorns and utilized fruits pierced by us with a fine needle, thus once more confirming our hypothesis on opportunism of females. Of course, one should bear in mind that the acorn-hardening threshold date is not strictly the same for every season and every location due to, e.g., differentiated duration in seed maturation depending on local conditions and innate features of plants; measurements showed that the plant of acorn origin was also an important variable. Indeed, the hardness and mass of acorns were significantly different depending on the tree (Table 1). We instead point to the fact that once acorns reach a stage of impenetrability, females are eager to utilize alternative ovipositional sites.
Additionally, in the last experiment, we aimed to determine the fecundity of females throughout the whole breeding season. Some species in the Curculio genus are known for reproductive diapause. This is the case for European species associated with oaks (C. glandium, C venosus and C. pellitus), as the imago is present and active from May, but insects start reproduction no sooner than July [12,30]. A similar phenomenon was described in a Nearctic species, C. sayi (Gyllenhall), associated with Castanea trees in the United States. According to field research, this weevil emerges from the soil twice a year—the first period being May/June, the second August/September. In addition, visible progress in reproductive organ development between the two periods was noted [31]. Our data show that within 51 days of reproduction activity, C. glandium females are capable of laying, on average, 64.8 eggs (n = 12; min–max: 49–75). This number is slightly higher than what other research presents. A study from the United Kingdom mentions 50 eggs as C. glanium female fecundity; still, the sample size was lower (n = 5) and beetles were dissected at the point in the season after which they could have produced more eggs, if still alive [32]. Fertility of other curculionids is notably higher—e.g., females of the large pine weevil Hylobius abietis (L.) deposit, on average, 70 eggs during one season [33]. A species closely related to Curculio spp., the strawberry blossom weevil Anthonomus rubi Herbst, has a mean fecundity of 157.6 eggs during a period of 71.6 days [34], whereas a rhynchocepharid, the red palm weevil Rhynchophorus ferrugineus (Olivier), can lay from 112 to 315 eggs [35].
Despite slight differences in egg number scopes before (0–3) and after (0–4) acorn hardening (which with the highest probability should be attributed to the advancement of breeding season), no significant differences in the egg output were revealed. This is another indication that, in favorable conditions, females are capable of laying eggs longer than they would in nature if only whole acorns were available.
Hereby presented observations are accordant with those made on C. occidentis, known for egg deposition solely into fresh, green acorns of Garry oak, which in very mild climatic conditions of southeastern Vancouver Island, Canada, is possible until 21 September. Females did not oviposit in mature, brown acorns and their egg number range was higher than that in C. glandium (0–6 vs. 0–4) [17]. The first pecan weevils had begun oviposition around 18 August (Luisiana, USA) and according to Calcote, the process continued until the pecan shells were impenetrable; however, the author did not closely link shell hardness and oviposition cessation [22]. In our view, the two should be linked. Another study on pecan development’s relation to C. caryae oviposition indicated that in the case of previously mentioned early-maturing cultivars, shell solidification occurred between the 11th and the 17th of August [21].
Daily fecundity of our females was rather low (2.4 eggs on average), most likely due to creation of time-consuming egg channels. On the other hand, it is common to observe eight or even more than ten eggs per acorn under natural conditions (M.R.: personal observations). This could be an outcome of a lack of ovipositional pheromones, as no Curculio species has been described as leaving chemical markings after successful oviposition. This is unlike females of the pepper weevil A. eugenii Cano, known for anal secretion deposition after laying eggs in flower buds [36,37]. Therefore, in C. glandium, high egg numbers in nature can come from either a female’s own mistakes, also considered self-superparasitism (female comes back and adds eggs to already exploited fruit/seed) [38], or from the superparasitism by other females. The latter is highly possible, as there is a limitation of resources (acorns) and, towards the end of the season, competition among females rises, as it was revealed in C. elephas associated with Q. ilex L. in Spain [39]. Superparasitism, defined as the reuse of seeds already exploited for oviposition, has been observed in another curculionid: the rice weevil Sitophilus oryzae (L.), which is also considered neither to secrete, nor to react to ovipositional chemical compounds [38]. In the absence of detailed data on both superparasitism and marking pheromones in C. glandium, it would be worth exploring these issues in further research.
5. Conclusions
Acorn hardening is a factor limiting the reproduction of weevils, otherwise females would continue to lay eggs for much longer period in the season, considerably boosting population size. This means that the quality of the resource (acorns) seems to be more important than the quantity of it, as even in mast years, the number of accessible fruits might not be very high. This in turn leads to increased competition between females, also forcing them to look for alternatives. At the same time, females are opportunists—when given a choice, they prefer broken or otherwise damaged acorns as ovipositional sites.
Author Contributions
Conceptualization, M.R.; methodology, M.R. and E.J.; software, M.R. and M.C.; formal analysis, M.C. and E.J.; investigation, M.R. and H.M.; resources, M.R.; data curation, M.R. and E.J.; writing—original draft preparation, M.R., H.M. and E.J.; writing—review and editing, H.M.; visualization, M.R., E.J. and M.C.; supervision, M.R.; project administration, M.R. All authors have read and agreed to the published version of the manuscript.
Funding
The work of M.R. was funded in part by the National Science Center, Poland, decision no. 2021/05/X/NZ9/00988.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
M.R. would like to thank and acknowledge Henryk Reut on his 99th birthday.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bonal, R.J.; Espelta, M.; Vogler, A.P. Complex selection on life-history traits and the maintenance of variation in exaggerated rostrum length in acorn weevils. Oecologia 2011, 167, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Dobrosavljevic, J.; Markovic, C.; Milanovic, S.; Vujicic, P.; Srbulovic, B.; Bojic, S. Influence of Curculio glandium (Marsham, 1802)(Coleoptera, Curculionidae) on Turkey Oak (Quercus cerris L., 1753) (Fagales, Fagaceae) Acorn Germination. In Proceedings of the IX International Agricultural Symposium “Agrosym 2018”, Sarajevo, Bosnia and Herzegovina, 4–7 October 2018. [Google Scholar]
- Espelta, J.M.; Bonal, R.; Sánchez-Humanes, B. Pre-dispersal acorn predation in mixed oak forests: Interspecific differences are driven by the interplay among seed phenology, seed size and predator size. J. Ecol. 2009, 97, 1416–1423. [Google Scholar] [CrossRef]
- Muñoz, A.; Bonal, R.; Espelta, J.M. Acorn–weevil interactions in a mixed-oak forest: Outcomes for larval growth and plant recruitment. Forest Ecol. Manag. 2014, 322, 98–105. [Google Scholar] [CrossRef]
- Wanat, M.; Mokrzycki, T. The Checklist of the Weevils (Coleoptera: Curculionoidea) of Poland Revisited. Ann. Zool. 2018, 68, 1–48. [Google Scholar] [CrossRef]
- Hughes, J.; Vogler, A.P. Ecomorphological adaptation of acorn weevils to their oviposition site. Evolution 2004, 58, 1971–1983. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Jafarpour, M.; Reut, M.; Shams Moattar, B.; Darvizeh, A.; Gorb, S.N.; Rajabi, H. Excavation mechanics of the elongated female rostrum of the acorn weevil Curculio glandium (Coleoptera; Curculionidae). Appl. Phys. A 2021, 127, 348. [Google Scholar] [CrossRef]
- Csóka, G.; Csókáné Hirka, A. Direct effects of carpophagous insects on the germination ability and early abscission of oak acorns. Acta Silv. Lignaria Hung. 2006, 2, 57–67. [Google Scholar]
- Oltean, I.; Stana, A. Curculio glandium, a pest which reduces the quality of the seed at Quercus petraea in Hilly Tree From Os Almaş, Ds Zalău. Bull. USAMV-CN 2007, 63, 123–126. [Google Scholar]
- Venner, S.; Pélisson, P.F.; Bel-Venner, M.C.; Débias, F.; Rajon, E.; Menu, F. Coexistence of insect species competing for a pulsed resource: Toward a unified theory of biodiversity in fluctuating environments. PLoS ONE 2011, 6, e18039. [Google Scholar] [CrossRef]
- Pélisson, P.F.; Bel-Venner, M.C.; Rey, B.; Burgevin, L.; Martineau, F.; Fourel, F.; Lecuyer, C.; Menu, F.; Venner, S. Contrasted breeding strategies in four sympatric sibling insect species: When a proovigenic and capital breeder copes with a stochastic environment. Funct. Ecol. 2012, 26, 198–206. [Google Scholar] [CrossRef]
- Reut, M.; Chrabąszcz, M.; Moniuszko, H. Timing Is Everything. Temporal and Spatial Niche Segregation in Curculio spp. (Coleoptera: Curculionidae) Associated with Oak Trees. Insects 2021, 12, 687. [Google Scholar] [CrossRef] [PubMed]
- Reut, M.; Moniuszko, H. Atypical Oviposition in Curculio glandium Marsham (Coleoptera: Curculionidae)—A Response to Unfavorable Environmental Conditions? Coleopt. Bull. 2022, 76, 36–38. [Google Scholar] [CrossRef]
- Addicott, F.T. Abscission; University of California Press: Berkeley, CA, USA, 1982; pp. 1–355. [Google Scholar]
- Bogdziewicz, M.; Espelta, J.M.; Muñoz, A.; Aparicio, J.M.; Bonal, R. Effectiveness of predator satiation in masting oaks is negatively affected by conspecific density. Oecologia 2018, 186, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Koenig, W.D.; Knops, J.M. The behavioral ecology of masting in oaks. In Oak Forest Ecosystems: Ecology and Management for Wildlife; Mc Shea, W.J., Healy, W.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2002; pp. 129–148. [Google Scholar]
- Rohlfs, D.A. A Study of Acorn Feeding Insects: Filbert Weevil (Curculio Occidentis (Casey)) and Filbertworm (Cydia Latiferreana (Walsingham)) on Garry Oak (Quercus Garryana) (Dougl.) in the Southeastern Vancouver Island Area. Doctoral Dissertation, University of British Columbia, Vancouver, BC, Canada, 1999. [Google Scholar]
- Holtz, B.A. Plant protection for pistachio. HortTechnology 2002, 12, 626–632. [Google Scholar] [CrossRef]
- Valentini, N.; Moraglio, S.T.; Rolle, L.G.C.; Tavella, L.; Botta, R. Nut and kernel growth and shell hardening in eighteen hazelnut cultivars (Corylus avellana L.). Hort. Sci. 2015, 42, 149–158. [Google Scholar] [CrossRef]
- Guidone, L.; Valentini, N.; Rolle, L.; Me, G.; Tavella, L. Early nut development as a resistance factor to the attacks of Curculio nucum (Coleoptera: Curculionidae). Ann. Appl. Biol. 2007, 150, 323–329. [Google Scholar] [CrossRef]
- Calcote, V.R.; Hyder, D.E. Pecan weevil preference for various pecan cultivars. J. Econ. Entomol. 1981, 74, 223–226. [Google Scholar] [CrossRef]
- Calcote, V.R. Pecan Weevil: Feeding and initial oviposition as related to nut development. J. Econ. Entomol. 1975, 68, 4–6. [Google Scholar] [CrossRef]
- Rakić, S.; Povrenović, D.; Tešević, V.; Simić, M.; Maletić, R. Oak acorn, polyphenols and antioxidant activity in functional food. J. Food Eng. 2006, 74, 416–423. [Google Scholar] [CrossRef]
- Sekeroglu, N.; Ozkutlu, F.; Kilic, E. Mineral composition of acorn coffees. Indian J. Pharm. Educ. Res. 2017, 51, 504–507. [Google Scholar] [CrossRef]
- Smreczyński, S. The Key for Identification of Polish Insects. Beetles–Coleoptera. Part XIX (98f). Weevils–Curculionidae. Subfamily–Curculioninae; Państwowe Wydawnictwo Naukowe: Warsaw, Poland, 1972; p. 195. [Google Scholar]
- Valentini, N.; Rolle, L.; Stévigny, C.; Zeppa, G. Mechanical behaviour of hazelnuts used for table consumption under compression loading. J. Sci. Food Agric. 2006, 86, 1257–1262. [Google Scholar] [CrossRef]
- Koyuncu, M.A.; Ekinci, K.; Savran, E. Cracking characteristics of walnut. Biosyst. Eng. 2004, 87, 305–311. [Google Scholar] [CrossRef]
- Zhang, G.Y.; Ji, Y.C.; Wen, X.J.; Li, Q.; Ren, Y.; Wen, J.B. Oviposition behaviour of Eucryptorrhynchus brandti (Coleoptera: Curculionidae: Cryptorrhychinae) on Ailanthus altissima (Mill.) Swingle (Sapindales: Simaroubaceae). Biocontrol Sci. Technol. 2017, 27, 1153–1167. [Google Scholar] [CrossRef]
- Bystrowski, C.; Wójcik, G. Use of neonicotinoid insecticides for the protection of pedunculate oak (Quercus robur L.) acorns in the seed orchard in the Lezajsk Forest District. Leśne Pr. Badaw. 2009, 70, 271. [Google Scholar] [CrossRef]
- Pelisson, P.F.; Bernstein, C.; Francois, D.; Menu, F.; Venner, S. Dispersal and dormancy strategies among insect species competing for a pulsed resource. Ecol. Entomol. 2013, 38, 470–477. [Google Scholar] [CrossRef]
- Keesey, I.W. The Chemical Ecology of the Lesser Chestnut Weevil: Behavioral and Electrophysiological Responses of Curculio sayi (Coleoptera: Curculionidae) to Host-Plant Volatile Organic Compounds. Doctoral Dissertation, University of Missouri, Columbia, MO, USA, 2011. [Google Scholar]
- Forrester, G.J. The Population Ecology of Acorn Weevils and Their Influence on Natural Regeneration of Oak. Doctoral Dissertation, University of London, London, UK, 1990. [Google Scholar]
- Doležal, P.; Kleinová, L.; Davídková, M. Adult Feeding Preference and Fecundity in the Large Pine Weevil, Hylobius abietis (Coleoptera: Curculionidae). Insects 2021, 12, 473. [Google Scholar] [CrossRef]
- Easterbrook, M.A.; Fitzgerald, J.D.; Pinch, C.; Tooley, J.; Xu, X.M. Development times and fecundity of three important arthropod pests of strawberry in the United Kingdom. Ann. App. Biol. 2003, 143, 325–331. [Google Scholar] [CrossRef]
- Faleiro, J.; Rangnekar, P.A.; Satarkar, V.R. Age and fecundity of female red palm weevils Rhynchophorus ferrugineus (Olivier) (Coleoptera: Rhynchophoridae) captured by pheromone traps in coconut plantations of India. Crop Prot. 2003, 22, 999–1002. [Google Scholar] [CrossRef]
- Addesso, K.M.; McAuslane, H.J.; Stansly, P.A.; Schuster, D.J. Host-marking by female pepper weevils, Anthonomus eugenii. Entomol. Exp. Appl. 2007, 125, 269–276. [Google Scholar] [CrossRef]
- Addesso, K.M.; Alborn, H.T.; Bruton, R.R.; McAuslane, H.J. A multicomponent marking pheromone produced by the pepper weevil, Anthonomus eugeni (Coleoptera: Curculionidae). Chemoecology 2021, 31, 247–258. [Google Scholar] [CrossRef]
- Campbell, J.F. Influence of seed size on exploitation by the rice weevil, Sitophilus oryzae. J. Insect Behav. 2002, 15, 429–445. [Google Scholar] [CrossRef]
- Bonal, R.; Muñoz, A. Seed growth suppression constrains the growth of seed parasites: Premature acorn abscission reduces Curculio elephas larval size. Ecol. Entomol. 2008, 33, 31–36. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).