

Genome-Based Taxa Delimitation (GBTD): A New Approach

Abstract
:1. Introduction
2. Materials and Methods
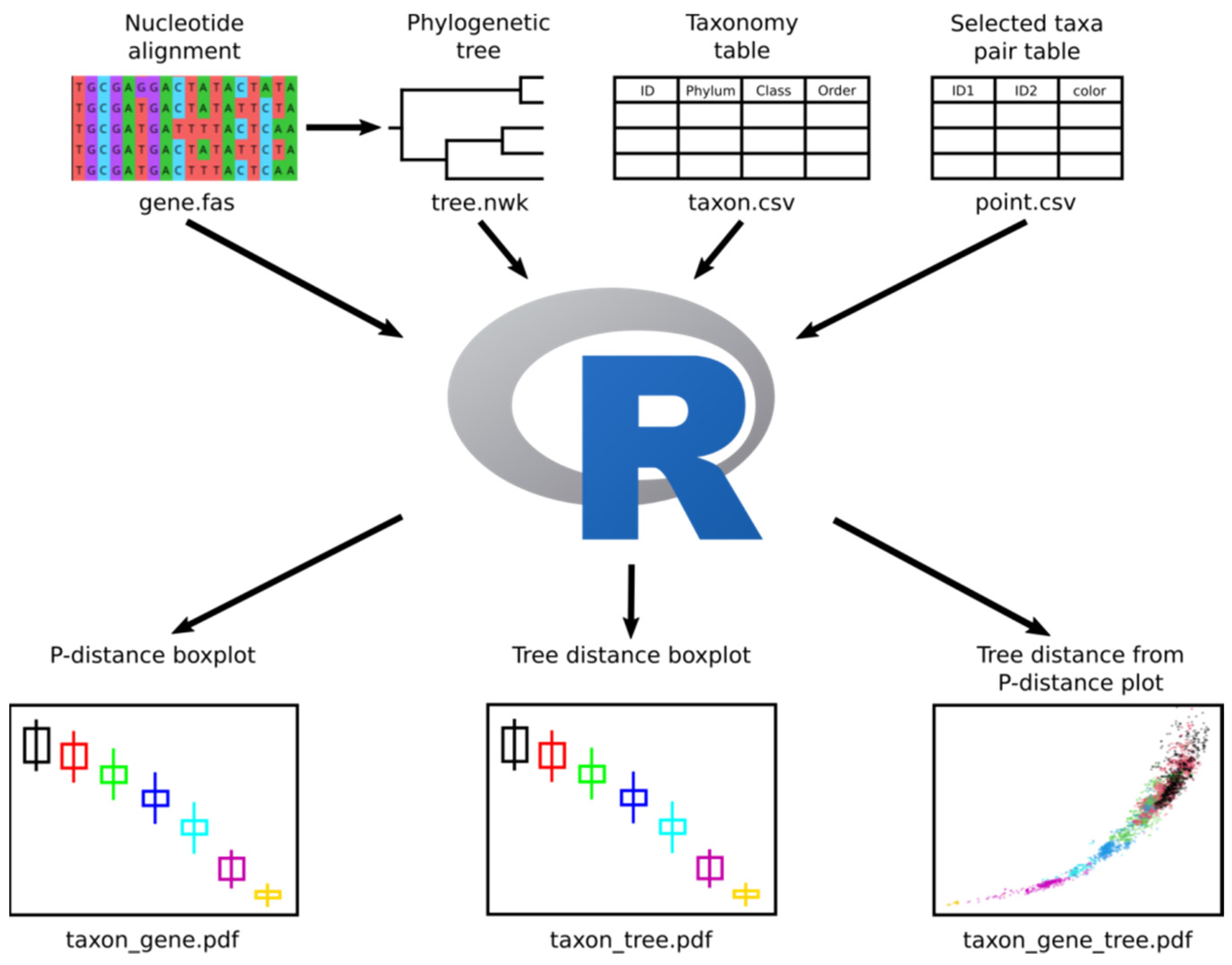
2.1. New Method Description
2.2. Sample Collection, DNA Extraction and Sequencing
2.3. Genome Assembly and Annotation
2.4. Mitogenome Alignment and Phylogeny Reconstruction
3. Results
4. Discussion
5. Conclusions
- Transfer Paracanthobdella livanowi into the genus Acanthobdella, with dissolution of both the genus Paracanthobdella and the family Paracanthobdellidae.
- Members of the genus Baicaloclepis should be synonymized with the genus Glossiphonia.
- Though the Piscicola and Codonobdella samples show little genetic divergence, there are significant morphological distinctions between these genera. Because the Piscicolidae samples are underrepresented in the studied dataset, careful reinvestigation of their position is required.
- Abolish the family Haemopidae by moving its representatives to the family Hirudinidae. Additionally, more research is required with the extended set of taxa, including the poorly described Whitmania.
- With growing evidence, it has becomes obvious that oligochaetes of the genera Amynthas and Metaphire should be considered one genus. Since the former genus was described more than a century prior to the latter, the Code of Zoological Nomenclature retains priority for the genus Amynthas.
- Regarding the splitting of the Eisenia nordenskioldi samples into different species, nothing can be said with confidence based solely on the genetic divergence. More careful morphological investigation is required.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hebert, P.; Cywinska, A.; Shelley, B.; de Waard, J. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Pons, J.; Barraclough, T.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.; Hazell, S.; Kamoun, S.; Sumlin, W.; Vogler, A. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapli, P.; Lutteropp, S.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate Poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics 2017, 33, 1630–1638. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Kaygorodova, I.; Bolbat, N.; Bolbat, A. Species delimitation through DNA barcoding of freshwater leeches of the Glossiphonia genus (Hirudinea: Glossiphoniidae) from Eastern Siberia, Russia. J. Zoolog. Syst. Evol. Res. 2020, 58, 1437–1446. [Google Scholar] [CrossRef]
- Remigio, E.A.; Hebert, P.D.N. Testing the utility of partial COI sequences for phylogenetic estimates of gastropod relationships. Mol. Phylogenet. Evol. 2003, 29, 641–647. [Google Scholar] [CrossRef]
- Saetang, T.; Marrone, F.; Vecchioni, L.; Maiphae, S. Morphological and molecular diversity patterns of the genus Tropodiaptomus Kiefer, 1932 (Copepoda, Calanoida, Diaptomidae) in Thailand. Sci. Rep. 2022, 12, 2218. [Google Scholar] [CrossRef]
- Phillips, A.; Dornburg, A.; Zapfe, K.; Anderson, F.; James, S.; Erséus, C.; Lemmon, E.; Lemmon, A.; Williams, B. Phylogenomic analysis of a putative missing link sparks reinterpretation of leech evolution. Genome Biol. Evol. 2019, 11, 3082–3093. [Google Scholar] [CrossRef] [Green Version]
- Galtier, N.; Nabholz, B.; Glémin, S.; Hurst, G.D. Mitochondrial DNA as a marker of molecular diversity: A reappraisal. Mol. Ecol. 2009, 18, 4541–4550. [Google Scholar] [CrossRef]
- Moore, S. Mitochondrial-gene trees versus nuclear-gene trees, a reply to Hoelzer. Evolution 1997, 51, 627–629. [Google Scholar] [CrossRef] [PubMed]
- Bolbat, A.; Vasiliev, G.; Kaygorodova, I. The first mitochondrial genome of the relic Acanthobdella peledina (Annelida, Acanthobdellida). Mitochondrial DNA B Resour. 2020, 5, 3300–3301. [Google Scholar] [CrossRef] [PubMed]
- Epstein, V.M. Acanthobdella livanowi sp. n.; a new species of ancient leeches (Archihirudinea) from Kamchatka. Proc. USSR Acad. Sci. 1966, 168, 955–958. [Google Scholar]
- Livanow, N. Acanthobdella peledina Grube, 1851. Zool. Jahrbücher. Anat. 1906, 22, 636–866. [Google Scholar]
- Lukin, E. Leeches of Fresh and Brackish Waters. Fauna of the USSR. (Vol. 1); Nauka: Leningrad, Russia, 1976; pp. 195–206. [Google Scholar]
- Holmquist, C. A Fish Leech of the genus Acanthobdella found in North America. Hydrobiologia 1974, 44, 2–3. [Google Scholar] [CrossRef]
- Hauck, A.; Fallon, M.; Burger, C. New host and geographical records for the leech Acanthobdella peledina Grube 1851 (Hirudinea, Acanthobdellidae). J. Parasitol. 1979, 65, 989. [Google Scholar] [CrossRef]
- Utevsky, S.; Sokolov, S.; Shedko, M. New records of the chaetiferous leech-like annelid Paracanthobdella livanowi (Epshtein, 1966) (Annelida: Clitellata: Acanthobdellida) from Kamchatka, Russia. Syst. Parasitol. 2013, 84, 71–79. [Google Scholar] [CrossRef]
- Epstein, V.M. worms—Annelida. In Identification Guide to Parasites of Freshwater Fish in the Soviet Union; Bauer, O., Ed.; Nauka: Leningrad, Russia, 1987; Volume 3, pp. 340–372. [Google Scholar]
- R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.R-project.org (accessed on 22 September 2022).
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 22 September 2022).
- Bolger, A.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Chevreux, B.; Wetter, T.; Suhai, S. Genome sequence assembly using trace signals and additional sequence information. In Proceedings of the Computer Science and Biology: Proceedings of the German Conference on Bioinformatics (GCB), Hannover, Germany, 4–6 October 1999; GBF-Braunschweig: Braunschweig, Germany, 1999; pp. 45–56. [Google Scholar]
- Milne, I.; Stephen, G.; Bayer, M.; Cock, P.; Pritchard, L.; Cardle, L.; Shaw, P.; Marshall, D. Using Tablet for visual exploration of second-generation sequencing data. Brief. Bioinform. 2013, 14, 193–202. [Google Scholar] [CrossRef]
- BLASTn Suite. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE=BlastSearch (accessed on 22 September 2022).
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Reichert, A.; Rothbauer, U.; Mörl, M. Processing and editing of overlapping tRNAs in human mitochondria. J. Biol. Chem. 1998, 273, 31977–31984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BLASTp Suite. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch (accessed on 22 September 2022).
- Tseemann/Barrnap: Bacterial Ribosomal RNA Predictor—GitHub. Available online: https://github.com/tseemann/barrnap (accessed on 2 September 2022).
- Katoh, K.; Standley, D. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.D.; Schrempf, M.D. Woodhams, A.; von Haeseler, R. Lanfear IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, S.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.; Ho, S.; Phillips, M.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS ONE 2006, 4, e88. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.; Xie, D.; Baele, G.; Suchard, M. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Dong, Y.; Jiang, J.; Qui, J. Three new species of earthworms belonging to the genera Amynthas and Metaphire (Oligochaeta: Megascolecidae) from Yunnan, China. J. Nat. His. 2019, 53, 1961–1974. [Google Scholar] [CrossRef]
- Seto, A.; Endo, H.; Minamiya, Y.; Matsuda, M. The complete mitochondrial genome sequences of Japanese earthworms Metaphire hilgendorfi and Amynthas yunoshimensis (Clitellata: Megascolecidae). Mitochondrial DNA B Resour. 2021, 6, 965–967. [Google Scholar] [CrossRef]
- Hussain, M.; Liaqat, I.; Mubin, M.; Nisar, B.; Shahzad, K.; Durrani, A.; Zafar, U.; Afzaal, M.; Ehsan, A.; Rubab, S. DNA barcoding: Molecular identification and phylogenetic analysis of pheretimoid earthworm (Metaphire sp. and Amynthas sp.) based on mitochondrial partial COI gene from Sialkot, Pakistan. J. Oleo Sci. 2022, 71, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Chang, C.-H.; Chih, W.-J. Two new earthworm species of the genus Amynthas (Oligochaeta: Megascolecidae) from central Taiwan, with comments on some recent species assignments in Amynthas and Metaphire. Zootaxa 2019, 4658, 101–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielecki, A.; Cichocka, J.M.; Jeleń, I.; Swiątek, P.; Płachno, B.J.; Pikuła, D. New data about the functional morphology of the chaetiferous leech-like annelids Acanthobdella peledina (Grube, 1851) and Paracanthobdella livanowi (Epshtein, 1966) (Clitellata, Acanthobdellida). J. Morphol. 2014, 275, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Siddall, M.; Burreson, E. Phylogeny of leeches (Hirudinea) based on mitochondrial cytochrome c oxidase subunit I. Mol. Phylogenet. Evol. 1998, 9, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Apakupakul, K.; Siddall, M.; Burreson, E. Higher level relationships of leeches (Annelida: Clitellata: Euhirudinea) based on morphology and gene sequences. Mol. Phylogenet. Evol. 1999, 12, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Trontelj, P.; Sket, B.; Steinbrück, G. Molecular phylogeny of leeches: Congruence of nuclear and mitochondrial rDNA data sets and the origin of bloodsucking. J. Zoolog. Syst. Evol. Res. 1999, 37, 141–147. [Google Scholar] [CrossRef]
- Tessler, M.; de Carle, D.; Voiklis, M.; Gresham, O.; Neumann, J.; Cios, S.; Siddall, M. Worms that suck: Phylogenetic analysis of Hirudinea solidifies the position of Acanthobdellida and necessitates the dissolution of Rhynchobdellida. Mol. Phylogenet. Evol. 2018, 127, 129–134. [Google Scholar] [CrossRef]
- Phillips, A.J.; Siddal, M.E. Poly-paraphyly of Hirudinidae: Many lineages of medicinal leeches. BMC Evol. Biol. 2009, 9, 246. [Google Scholar] [CrossRef] [Green Version]
- Richardson, L.R. A contribution to the systematics of the hirudinid leeches, with description of new families, genera and species. Acta Zool. Acad. Sci. Hungricae 1969, 15, 97–149. [Google Scholar]
- Lukin, E.I.; Epstein, V.M. Leeches of subfamily Toricinae nov. subfam. and their geographic distribution. Proc. USSR Acad. Sci. 1960, 131, 478–481. [Google Scholar]
- Shekhovtsov, S.; Golovanova, E.; Ershov, N.; Poluboyarova, T.; Berman, D.; Bulakhova, N.; Szederjesi, T.; Peltek, S.; Schrader, S. Phylogeny of the Eisenia nordenskioldi complex based on mitochondrial genomes. Eur. J. Soil Biol. 2020, 96, 103137. [Google Scholar] [CrossRef]
- Envall, I.; Källersjö, M.; Erséus, C. Molecular evidence for the non-monophyletic status of Naidinae (Annelida, Clitellata, Tubificidae). Mol. Phylogenet. Evol. 2006, 40, 570–584. [Google Scholar] [CrossRef] [PubMed]
- Erseus, C.; Wetzel, M.; Gustavsson, L. ICZN rules—A farewell to Tubificidae (Annelida, Clitellata). Zootaxa 2008, 1744, 66–68. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sampling Location and Coordinates or Data Accession | GenBank Number |
|---|---|---|
| Acanthobdella peledina | River Pitea, Sweden; 66.449778, 18.03977 | MZ562997 |
| River Iya, Irkutsk Region, Russia; 53.804354, 99.558638 | OM203184 | |
| River Middle Rassokha, Irkutsk Region, Russia; 58.001986, 109.466141 | OM203186 | |
| Lake Bolshoy Kharbey, Nenets AD, Russia; 67.556111, 62.889722 | OM117616 | |
| Lake Labynkyr, Sakha (Yakutia), Russia; 62.498315, 143.598658 | OM203185 | |
| River Yana, Sakha (Yakutia), Russia; 67.553945, 133.367242 | OM214536 | |
| Paracanthobdella livanowi | Lake Azhabachje, Kamchatka, Russia; 56.146114, 161.802687 | OM117614 |
| OM117615 | ||
| Codonobdella sp. | Lake Baikal, Russia; 53.240811, 107.234195 | MZ202177 |
| Baicaloclepsis grubei | Lake Baikal, Russia; 53.240811, 107.234195 | OM257165 |
| Baicaloclepsis echinulata | Lake Baikal, Russia; 53.012873, 106.920606 | OM257166 |
| Erpobdella octoculata | SRA accession: SRX9009198 | OM257408 |
| Glossiphonia complanata | SRA accession: SRX8928147 | OM039422 |
| Haemopis sanguisuga | SRA accession: SRX9009141 | OM234778 |
| Haemopis sanguisuga | SRA accession: SRX9009400 | OM234779 |
| Lumbriculus variegatus | SRA accession: SRX9009164 | OM062609 |
| Piscicola geometra | SRA accession: SRX9009199 | BK059172 |
| Theromyzon tessulatum | SRA accession: SRX8928146 | OM039423 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolbat, A.; Bukin, Y.; Kaygorodova, I. Genome-Based Taxa Delimitation (GBTD): A New Approach. Diversity 2022, 14, 948. https://doi.org/10.3390/d14110948
Bolbat A, Bukin Y, Kaygorodova I. Genome-Based Taxa Delimitation (GBTD): A New Approach. Diversity. 2022; 14(11):948. https://doi.org/10.3390/d14110948
Chicago/Turabian StyleBolbat, Alexander, Yurij Bukin, and Irina Kaygorodova. 2022. "Genome-Based Taxa Delimitation (GBTD): A New Approach" Diversity 14, no. 11: 948. https://doi.org/10.3390/d14110948