Systematic Paleontology
Superfamily Bostrichoidea Latreitte, 1802.
Family Bostrichidae Latreille, 1802.
Subfamily Alitrepaninae Peng, Jiang, Engel & Wang, 2022.
Genus Poinarinius Legalov, 2018.
Type species. Poinarinius burmaensis Legalov, 2018.
Alitrepanum Peng, Jiang, Engel & Wang, 2022, syn. nov.
Type species. Alitrepanum aladelicatum Peng, Jiang, Engel & Wang, 2022.
Diagnosis. Body subcylindrical, usually covered with long erect or semierect setae; length 1.7–4.9 mm. Head hypognathous, visible from above, wider than anterior margin of pronotum. Mandible quite large, curved. Galea with dense, long, erect setae. Labrum almost semicircular, with dense setae. Eyes convex, finely faceted. Frons convex or impressed in middle, densely or finely punctate, sometimes transversally-rugose, usually covered with more or less dense erect setae, sometimes with tubercle in the middle. Vertex weakly convex or weakly impressed in middle, punctate, or longitudinally-rugose, sometimes with two tubercles. Antennae quite short, 8-segmented. First antennomere long-conical or suboval. Second antennomere conical. Third antennomeres conical or suboval. Fourth antennomere conical, wide-conical, or suboval. Fifth antennomere wide-conical. Antennal club formed with three terminal transverse antennomeres. Pronotum concave at anterior margin. Disk usually weakly convex, punctate, sometimes longitudinally-rugose. Scutellum trapezoidal. Elytra subcylindrical or suboval. Elytral intervals from wide to narrow, flat or convex, wider, subequal or narrow than striae. Elytra lacking distinct elytral declivity and lobes or with more or less distinct elytral declivity and ones. Elytral declivity almost vertical or oblique. Elytra sometimes with two transverse or longitudinal lobes, or two spines before elytral declivity; with longitudinal lobes or double spines on sides of elytral declivity; with on sides of elytral declivity or before one. Metanepisterna quite wide. Metaventrite 2.4–3.2 times as long as the metacoxal cavity length. Procoxae elongate and widely separated. Metatrochanter triangular, partially separating femur from coxa. Profemora distinctly thickened. Meso- and metafemora weakly thickened. Protibiae curved, with sixth spines, lacking spurs. Meso- and metatibiae flattened, with two apical spurs. Mesotibial spurs equal in length, quite long. Metabibiae with long spur, almost equal in length to the first tarsomere and quite short one. Tarsi with five narrow tarsomeres. Protarsi subequal or longer than protibiae. Meso- and metatarsi longer than meso- and metatibiae. Claws free, laminate at base.
Notes. The main differences between the genus Poinarinius and the genus Alitrepanum were the 9- and 10-segmented antennae and the structure of the apices of the elytra. The study of the material showed that Poinarinius burmaensis and Alitrepanum aladelicatum have 8-segmented antennae, and the structure of the apex of the elytra can be different, from elytra without a elytral declivity to elytra with one and with spines or lobes. Therefore, Alitrepanum Peng, Jiang, Engel & Wang, 2022, syn. nov. is synonymous with Poinarinius Legalov, 2018. The body is covered with more or less dense long erect or semierect setae, 8-segmented antennae with transverse antennal club, concave at anterior margin pronotum, elongate and widely separated procoxae, curved protibiae, with sixth spines, lacking spurs, meso- and metatibiae with two apical spurs, metabibiae with two spurs unequal in length and protarsi subequal or longer than protibiae are important characteristics of the species of the genus.
Poinarinius aristovi Legalov & Háva,
sp. n. (
Figure 1A–F).
LSIDurn:lsid:zoobank.org:act:396688E6-E5AA-48E5-8645-28C614316B24.
Description. Male. Body brown, length 2.4–3.7 mm. Integument covered with sparse, long, erect setae. Head spherical, densely punctate, 1.1–1.5 times as wide as pronotum apex width. Frons impressed in middle, finely punctate, with setae. Pronotum 0.7–1.6 times as long as wide at apex, 0.6–1.8 times as long as wide in middle, 0.7–1.6 times as long as wide at base. Disk weakly convex, finely and sparsely. Sides weakly rounded, lacking erect setae. Elytra suboval, 2.0–3.4 times as long as the pronotum, 1.9–2.6 times as long as wide at the base, 1.7–2.3 times as long as wide in the middle, 1.8–2.4 times as long as the apical fourth. Sides of elytra without erect setae. Elytral intervals wide, flat, 3.5–4.0 times wide as striae. Striae not deep, weak. Elytral declivity absent. Metaventrite about 2.4 times as the long as the metacoxal cavity length. First ventrite about 1.4 times as long as the metacoxal cavity length. Second ventrite about 0.9 times as long as the first ventrite. Third ventrite about 1.1 times as long as the second ventrite. Fourth ventrite is 1.0–1.2 times as long as the third ventrite. Fifth ventrite 0.7–1.1 times as long as the fourth ventrite. Metabibial long spur slightly shorter than the fifth tarsomere. Metabibial short, about 0.6 times as long as the fifth tarsomere. Protarsi about 1.5 times as long as the protibiae. Mesotarsi slightly longer than the mesotibiae. Metatarsi about 1.3 times as long as the metatibiae. Protarsi: first tarsomere about 2.7 times as long as the second tarsomere; second tarsomere about 1.2 times as long as the third tarsomere; fourth tarsomere about 0.7 times as long as the third tarsomere; fifth tarsomere about 2.6 times as long as the fourth tarsomere, slightly longer than the third and fourth tarsomeres combined. Mesotarsi: first tarsomere about 1.8 times as long as the second tarsomere; second tarsomere about 1.2 times as long as the third tarsomere; fourth tarsomere subequal to the third tarsomere; fifth tarsomere about 2.4 times as long as the fourth tarsomere, about 1.2 times as long as the third and fourth tarsomeres combined. Metatarsi: first tarsomere about 4.7 times as long as the second tarsomere; second tarsomere subequal to the third tarsomere; fourth tarsomere about 0.9 times as long as the third tarsomere; fifth tarsomere about 2.3 times as long as the fourth tarsomere, slightly longer than the third and fourth tarsomeres combined. Female. Body length 2.6–4.9 mm. Integument covered with long erect setae. Head spherical, densely punctate, about 1.2 times as wide as the pronotum apex width. Frons weakly impressed in middle, with dense erect setae. First antennomere oval, about 1.6 times as long as wide in the middle. Second antennomere conical, about 0.9 times as long as wide at the apex, about 1.3 times as long as and about 0.7 times as narrow as the first antennomere. Third antennomere long-conical, about 1.7 times as long as wide at the apex, 0.5 times as long as and about 0.7 times as narrow as the second antennomere. Fourth antennomere conical, about 0.9 times as long as wide at apex, about 0.6 times as long as and about 1.1 times as wide as third antennomere. Fifth antennomere wide-conical, about 0.7 times as long as wide at apex, about 0.9 times as long as and about 1.1 times as wide as the fourth antennomere. Sixth antennomere wide-conical, about 0.5 times as long as wide at apex, about 0.8 times as long as and about 1.1 times as wide as the fifth antennomere. Pronotum about 0.7 times as long as wide at the apex, about 0.6 times as long as wide in the middle, about 0.7 times as long as wide at the base. Sides with erect setae. Elytra about 3.3 times as long as the pronotum, about 2.1 times as long as wide at the base, about 1.8 times as long as wide in the middle, about 2.2 times as long as the apical fourth. Sides of elytra with erect setae. Metaventrite 2.4–2.6 times as long as metacoxal cavity length. First ventrite about 1.5 times as long as the metacoxal cavity length. Second ventrite subequal to the first ventrite. Third ventrite 1.0–1.3 times as long as the second ventrite. Fourth ventrite 0.8–1.1 times as long as the third ventrite. Fifth ventrite 1.1–1.4 times as long as the fourth ventrite.
Material examined. Holotype—male (ISEA), no. MA2016/2. Paratypes: male (ISEA), no. MA2016/11; male (ISEA), no. MA2018/14; male (ISEA), no. MA2020/11; male (ISEA), no. MA2020/13; male (ISEA), no. MA2020/15; male (ISEA), no. MA2020/21; male (ISEA), no. MA2020/22; male (ISEA), no. MA2020/23; male (ISEA), no. MA2020/26; male (ISEA), no. MA2020/28; male (ISEA), no. MA2020/36; male (ISEA), no. MA2020/37; male (ISEA), no. MA2020/38; female (ISEA), no. MA2020/25; female (ISEA), no. MA2020/27; female (ISEA), no. MA2020/39; female (JHAC), no. JH2022/7; female (JHAC), JH2022/10.
Etymology. The species is named after the late paleoentomologist Danil S. Aristov (Moscow).
Comparison. The new species differs from other species of the genus (excluding P. antonkozlovi sp. nov.) in the elytra lacking elytral declivity and lobes. It is distinguished from P. antonkozlovi sp. nov. in the punctate pronotum.
Poinarinius antonkozlovi Legalov & Háva,
sp. n. (
Figure 1G,H).
LSIDurn:lsid:zoobank.org:act: 20F616EF-F921-4EB7-9418-41E5B1284936.
Description. Female. Body dark brown, length 2.7–2.9 mm. Head slightly wider than the pronotum apical width. Frons impressed, densely punctate, with dense, long, erect setae. Pronotum about 1.1 times as long as wide at the apex, slightly narrower than wide in the middle, slightly longer than wide at the base. Disk weakly convex, coarse, longitudinally-rugose. Sides weakly rounded, with long erect setae. Elytra subcylindrical, about 2.1 times as long as the pronotum, about 1.7 times as long as wide at the base, about 2.0 times as long as wide in the middle, about 2.4 times as long as the apical fourth. Sides of elytra with very sparse, long, erect setae. Elytral intervals narrow, flattened, punctate, 1.5–2.0 times wide as striae. Striae deep. Fifth interval without lobate convexity. Legs with sparse, long, erect setae. Metabibiae with long spur, which is almost equal in length to the first tarsomere and quite short one. Protarsi subequal in length to protibiae. Mesotarsi about 1.3 times as long as the mesotibiae. Metatarsi about 1.2 times as long as the metatibiae. Tarsi longer than tibiae, with five narrow tarsomeres. Protarsi: first tarsomere about 1.4 times as long as the second tarsomere; second tarsomere about 1.3 times as long as the third tarsomere; fourth tarsomere about 0.8 times as long as the third tarsomere; fifth tarsomere about 2.7 times as long as the fourth tarsomere, slightly shorter than the third and fourth tarsomeres combined. Mesotarsi: first tarsomere about 1.3 times as long as the second tarsomere; second tarsomere about 1.4 times as long as the third tarsomere; fourth tarsomere about 0.6 times as long as the third tarsomere; fifth tarsomere about 1.7 times as long as the fourth tarsomere, about 0.6 times as long as the third and fourth tarsomeres combined. Metatarsi: first tarsomere about 2.2 times as long as the second tarsomere; second tarsomere about 1.1 times as long as the third tarsomere; fourth tarsomere about 0.7 times as long as the third tarsomere; fifth tarsomere about 2.8 times as long as the fourth tarsomere, about 1.1 times as long as the third and fourth tarsomeres combined.
Material examined. Holotype—female (ISEA), no. MA2020/24. Paratypes: female (ISEA), no. MA2018/15; female (JHAC), no. JH2022/5.
Etymology. In honor of Anton Kozlov (Moscow), who helped us in the study.
Comparison. The new is very similar to P. aristovi sp. nov., but it is easily distinguished by transverse rugose pronotum.
Poinarinius perkovskyi Legalov & Háva,
sp. n. (
Figure 2A).
LSIDurn:lsid:zoobank.org:act:10D44B00-6920-4A28-9198-89E319409CFF.
Description. Male. Body brown, length 3.0 mm. Integument almost glabrous. Frons weakly impressed in the middle, finely and sparsely punctate, without setae. First antennomere of club about 3.0 times as wide as the length. Second antennomere of club about 4.3 times as wide as the length. Third antennomere of club about 3.6 times as wide as the length. Pronotal disk weakly convex, finely and sparsely punctate. Sides weakly rounded, lacking erect setae. Elytra subcylindrical, about 2.9 times as long as the pronotum. Sides of elytra without erect setae. Elytral intervals wide, flat, about 3.3 times as wide as the striae. Striae not deep. Elytral declivity almost vertical. Elytra with two spines before elytral declivity. Metaventrite 3.0 times as long as the metacoxal cavity length. Metanepisterna 3.1 times as long as wide in the middle. Abdomen convex. First ventrite about 0.8 times as long as the metacoxal cavity length. Second ventrite equal to the first ventrite. Third ventrite slightly longer than the second ventrite. Fourth ventrite slightly longer than the third ventrite. Fifth ventrite about 1.3 times as long as the fourth ventrite. Protibiae curved, with sixth spines, lacking spurs. Meso- and metatibiae flattened, with two apical spurs. Mesotibial spurs equal in length, quite long. Metabibiae with a long spur and quite a short one. Protarsi about 1.4 times as long as protibiae.
Mesotarsi about 1.9 times as long as the mesotibiae. Metatarsi about 1.6 times as long as the metatibiae. Tarsi longer than tibiae, with five narrow tarsomeres. Protarsi: first tarsomere about 2.3 times as long as the second tarsomere; second tarsomere about 0.8 times as long as the third tarsomere; fourth tarsomere about 0.5 times as long as the third tarsomere; fifth tarsomere about 2.2 times as long as the fourth tarsomere, slightly shorter than the third and fourth tarsomeres combined. Mesotarsi: first tarsomere about 2.0 times as long as the second tarsomere; second tarsomere about 0.9 times as long as the third tarsomere; fourth tarsomere about 0.5 times as long as the third tarsomere; fifth tarsomere about 2.9 times as long as the fourth tarsomere, slightly shorter than the third and fourth tarsomeres combined. Metatarsi: first tarsomere about 2.8 times as long as the second tarsomere; second tarsomere about 0.9 times as long as the third tarsomere; fourth tarsomere about 0.5 times as long as the third tarsomere; fifth tarsomere about 3.7 times as long as the fourth tarsomere, about 1.2 times as long as the third and fourth tarsomeres combined.
Material examined. Holotype—male (ISEA), no. MA2020/4.
Etymology. In honor of Evgeny E. Perkovsky (Kiev) who contributed to the study of amber faunas of the Cretaceous and the Eocene.
Comparison. The new differs from P. aladelicatus in almost vertical elytral declivity. It is very similar to P. lesnei sp. nov., but is distinguished by the elytra with two spines before elytral declivity.
Poinarinius aladelicatus (Peng, Jiang, Engel & Wang, 2022),
comb. nov. (
Figure 2B–F).
Redescription. Body red-brown, brown or black-brown, length 1.7–2.7mm. Integument covered with erect and semierect setae. Head slightly wider than the pronotum apical width. Frons convex, densely punctate, covered with rare erect setae. Vertex weakly impressed in the middle. First antennomere suboval, about 1.7 times as long as wide at the apex. Second antennomere conical, about 1.3 times as long as wide at the apex, about 0.6 times as long as and about 0.8 times as narrow as the first antennomere. Third and fourth antennomeres suboval. Third antennomere about 1.8 times as long as wide at the apex, about 0.6 times as long as and about 0.4 times as narrow as the second antennomere. Fourth antennomere equal to the third antennomere. Fifth antennomere wide-conical about 0.8 times as long as wide at the apex, about 0.6 times as long as and about 1.3 times as wide as the fourth antennomere. Sixth antennomere about 0.6 times as long as wide at the apex, about 3.0 times as long as and about 4.0 times as wide as the fifth antennomere. Seventh antennomere subequal to about 0.5 times as long as wide at the apex, slightly shorter and subequal in width to the sixth antennomere. Eighth antennomere about 0.7 times as long as wide at the apex, subequal in length, and 0.7 times as narrow as the seventh antennomere. Pronotum 0.9–1.0 times as long as wide at the apex and in the middle, 1.0–1.1 times as long as wide at the base. Disk weakly flattened, densely punctate and longitudinally rugose. Elytra subcylindrical, 2.3–2.5 times as long as the pronotum, 1.8–2.3 times as long as wide at the base, 1.8–2.2 times as long as wide in the middle, 1.8–2.1 times as long as the apical fourth. Elytral intervals narrow, convex, equal in width or narrower than the striae. Striae deep, with large rounded punctation. Elytra with distinct elytral declivity, without lobes. Edges of elytral declivity serrated. Metaventrite about 2.5 times as long as the metacoxal cavity length. First ventrite slightly shorter than the metacoxal cavity length. Second ventrite slightly longer than the first ventrite. Third ventrite slightly shorter than the second ventrite. Fourth ventrite subequal to the third ventrite. Fifth ventrite slightly shorter than the fourth ventrite. Protarsi subequal in length to the protibiae. Mesotarsi about 1.5 times as long as the mesotibiae. Metatarsi about 1.3 times as long as the metatibiae. Protarsi: first tarsomere about 1.3 times as long as the second tarsomere; second tarsomere about 1.1 times as long as the third tarsomere; fourth tarsomere about 0.8 times as long as the third tarsomere; fifth tarsomere about 2.3 times as long as the fourth tarsomere, subequal to the third and fourth tarsomeres combined. Mesotarsi: first tarsomere about 1.8 times as long as the second tarsomere; second tarsomere subequal to the third tarsomere; fourth tarsomere about 0.8 times as long as the third tarsomere; fifth tarsomere about 2.3 times as long as the fourth tarsomere, subequal to the third and fourth tarsomeres combined. Metatarsi: first tarsomere about 1.6 times as long as second tarsomere; second tarsomere about 1.1 times as long as third tarsomere; fourth tarsomere about 0.8 times as long as third tarsomere; fifth tarsomere about 3.0 times as long as the fourth tarsomere, about 1.3 times as long as the third and fourth tarsomeres combined.
Material examined. Male (ISEA), no. MA2018/30; male (ISEA), no. MA2018/20; male (ISEA), no. MA2019/26; male (ISEA), no. MA2020/1; male (ISEA), no. MA2020/2; male (ISEA), no. MA2020/3; male (ISEA), no. MA2020/11; male (ISEA), no. MA2020/18; male (ISEA), no. MA2020/19; ex. (ISEA), no. MA2020/16; ex. (ISEA), no. MA2020/17; ex. (ISEA), no. MA2020/30; ex. (ISEA), no. MA2020/31; male (ISEA), no. MA 2018/24; (JHAC), no. JH2022/1; ex (JHAC), no. JH2022/2; ex. (JHAC), no. JH2022/3.
Remarks. This species is one of the most common Bostrichidae in Burmese amber. Peng et al. (2022) indicated 10-segmented antennae in the original description. We studied the antennae of our specimens, which were 8-segmented. It is likely that some antennomeres were torn during fossilization and Peng et al. (2022) mistakenly took them for different segments.
Poinarinius zahradniki Legalov & Háva,
sp. n. (
Figure 2G,H).
LSIDurn:lsid:zoobank.org:act:76AEDE38-90AC-4ED5-8F25-37279B695F09.
Description. Female. Body brown, length 3.3 mm. Head about 1.1 times as wide as the pronotum apical width. Frons convex, densely punctate, with tubercle in the middle, covered with sparse erect setae. Vertex with two tubercles. Pronotum about 1.3 times as long as wide at the apex, equal in wide in the middle, about 1.1 times as long as wide at the base. Disk weakly convex, densely and finely punctate. Elytra cylindrical, about 2.1 times as long as the pronotum, about 1.9 times as long as wide at the base, in the middle, and at the apical fourth. Sides of elytra with sparse, erect setae. Elytral intervals narrow, weakly convex, subequal in width to the striae. Striae deep. Elytra with distinct oblique elytral declivity, without lobes. Edges of elytral declivity even, quite high. Metaventrite about 3.2 times as long as the metacoxal cavity length. Protarsi subequal in length to the protibiae. Protarsi: first tarsomere about 1.6 times as long as the second tarsomere; second tarsomere subequal to the third tarsomere; fourth tarsomere about 0.8 times as long as the third tarsomere; fifth tarsomere about 2.0 times as long as the fourth tarsomere, slightly shorter than the third and fourth tarsomeres combined.
Material examined. Holotype—female (ISEA), no. MA2018/27.
Etymology. In honor of Petr Zahradník (Prague), specialist in Bostrichidae and Ptinidae.
Comparison. The new is similar to P. aladelicatus but differs in the larger body (3.3 mm) and even higher edges of the elytral declivity.
Poinarinius burmaensis Legalov, 2018.
Material examined. Holotype—female, (ISEA), no. MA2018/3; female (JHAC), JH2021/1; female (JHAC), no. JH2021; ex. (JHAC), no. JH2022.
Remarks. Description see in Legalov (2018).
Poinarinius borowskii Legalov & Háva,
sp. n. (
Figure 3A–C,E).
LSIDurn:lsid:zoobank.org:act:31607A3B-BEA8-4B43-9DF3-EC637F4BDDDF.
Description. Body brown, length 3.8–4.5 mm, covered with long quite dense erect setae. Frons flattened, densely punctate, covered with sparse erect setae. Pronotum about 1.1 times as long as wide at the apex, subequal in wide in the middle, about 1.3 times as long as wide at the base. Disk weakly convex, sparsely, and finely punctate. Elytra subcylindrical, about 2.6 times as long as the pronotum, about 2.4 times as long as wide at the base, 2.0 times as long as wide in the middle, and about 2.2 times as long as wide at the apical fourth. Disk and sides of elytra with erect setae. Elytral intervals wide, flat, about 5.0 times as wide as the striae. Striae weak. Elytra with weak elytral declivity and with double spines on the sides of elytral declivity. Protarsi about 1.1 times as long as the protibiae. Mesotarsi about 1.4 times as long as the mesotibiae. Protarsi: first tarsomere about 2.3 times as long as the second tarsomere; second tarsomere subequal to the third tarsomere; fourth tarsomere about 0.5 times as long as the third tarsomere; fifth tarsomere about 3.0 times as long as the fourth tarsomere, equal to the third and fourth tarsomeres combined. Mesotarsi: first tarsomere about 2.0 times as long as the second tarsomere; second tarsomere subequal to the third tarsomere; fourth tarsomere about 0.7 times as long as the third tarsomere; fifth tarsomere about 2.0 times as long as the fourth tarsomere, about 0.8 times as long as the third and fourth tarsomeres combined. Metatarsi: first tarsomere about 2.3 times as long as the second tarsomere; second tarsomere subequal to the third tarsomere; fourth tarsomere about 0.5 times as long as the third tarsomere; fifth tarsomere about 3.3 times as long as the fourth tarsomere, about 1.1 times as long as the third and fourth tarsomeres combined.
Material examined. Holotype—female (ISEA), no. MA2018/8. Paratypes: male (JHAC), no. JH2022/9; female (ISEA), no. MA2018/25.
Etymology. In honor of Jerzy Borowski (Warszawa), specialist in Bostrichidae and Ptinidae.
Comparison. The new is distinguished from P. burmaensis in the punctate pronotum. It differs from P. cretaceus sp. nov. in the elytra with double spines on the sides of the elytral declivity.
LSIDurn:lsid:zoobank.org:act:D15AFE9F-0574-44D4-BFF4-CD70EE9D3C99.
Description. Female. Body dark brown, length 3.1 mm, covered with long, erect setae. Head about 1.1 times as wide as the pronotum apical width. Frons weakly convex, densely punctate, with tubercle in middle, covered with quite dense erect setae. Vertex with two tubercles. Pronotum about 0.8 times as long as wide at the apex and in the middle, 0.9 times as long as wide at the base. Disk weakly flattened, finely punctate. Sides weakly rounded, with long erect setae. Elytra subcylindrical, about 2.5 times as long as the pronotum, about 1.7 times as long as wide at the base, about 1.5 times as long as wide in the middle, about 1.6 times as long as the apical fourth. Sides of elytra with long, erect setae. Elytral intervals flattened, punctate, 1.4–1.6 times as the striae. Striae deep. Elytra with double spines on the sides of the weak elytral declivity. Metaventrite 2.5 times as long as the metacoxal cavity length. First ventrite slightly longer than the metacoxal cavity length. Second ventrite equal to the first ventrite. Third ventrite about 1.2 times as long as the second ventrite. Protarsi about 1.3 times as long as the protibiae. Mesotarsi about 2.3 times as long as the mesotibiae. Metatarsi about 1.6 times as long as the metatibiae. Protarsi: first tarsomere 2.0 times as long as the second tarsomere; second tarsomere subequal to the third tarsomere; fourth tarsomere 0.7 times as long as the third tarsomere; fifth tarsomere about 2.3 times as long as the fourth tarsomere, slightly shorter than the third and fourth tarsomeres combined. Mesotarsi: first tarsomere 1.8 times as long as second tarsomere; second tarsomere about 1.1 times as long as third tarsomere; fourth tarsomere about 0.8 times as long as third tarsomere; fifth tarsomere about 2.7 times as long as fourth tarsomere, about 1.2 times as long as than third and fourth tarsomeres combined. Metatarsi: first tarsomere about 1.7 times as long as the second tarsomere; second tarsomere subequal to the third tarsomere; fourth tarsomere about 0.6 times as long as the third tarsomere; fifth tarsomere about 2.6 times as long as the fourth tarsomere, slightly shorter than the third and fourth tarsomeres combined.
Material examined. Holotype—female (ISEA), no. MA2018/9; Paratypes: female (JHAC, no. JH2022/6; female (JHAC, no. JH2022/8.
Etymology. In memory dedicated to Pierre Lesne (*9.4.1871-†10.11.1949)—the famous expert of the family Bostrichidae.
Comparison. The new differs from P. perkovskyi sp. nov. in the elytra with two transverse lobes before the elytral declivity.
Poinarinius cretaceus Legalov & Háva,
sp. n. (
Figure 4A–E).
LSIDurn:lsid:zoobank.org:act:3479B170-66A1-4163-95AE-22893E9E1DCE.
Description. Male. Body red-brown, length 2.8–3.2 mm. Integument covered with erect and semierect long setae. Head about 1.2 times as wide as the pronotum apical width. Frons convex, sparsely punctate, covered with rare erect setae. Vertex weakly convex. First antennomere long conical, weakly curved, about 2.3 times as long as wide at the apex. Second and third antennomeres conical. Second antennomere about 1.4 times as long as wide at the apex, about 0.4 times as long as and about 0.7 times as narrow as the first antennomere. Third antennomere subequal in the length and width, about 0.6 times as long as and about 0.8 times as narrow as the second antennomere. Fourth and fifth antennomeres are wide-conical. Fourth antennomere about 0.6 times as long as wide at the apex, about 0.6 times as long as and about 1.1 times as wide as the third antennomere. Fifth antennomere about 0.7 times as long as wide at the apex, about 1.2 times as long as and equal in width to the fourth antennomere. Sixth antennomere about 0.7 times as long as wide at the apex, about 2.7 times as long as and about 2.7 times as wide as the fifth antennomere. Seventh antennomere about 0.4 times as long as wide at the apex, equal in length and about 2.9 times as wide as the sixth antennomere. Eighth antennomere about 0.3 times as long as wide at the apex, slightly shorter, and slightly narrower than the seventh antennomere. Pronotum about 0.9 times as long as wide at the apex, in the middle, and at the base. Disk weakly convex, densely and finely punctate, with quite long setae. Elytra subcylindrical, 2.2–2.6 times as long as the pronotum, 1.6–2.0 times as long as wide at the base, about 1.6 times as long as wide in the middle, 1.6–1.7 times as long as the apical fourth. Elytral intervals narrow, convex, about 0.7 times as narrow as the striae. Striae deep, with large rounded punctation. Elytra with two longitudinal lobes before weak elytral declivity. Metaventrite about 1.8 times as long as the metacoxal cavity length. Metanepisterna about 3.8 times as long as wide in the middle. First ventrite about 0.6 times as long as the metacoxal cavity length. Second ventrite slightly longer than the first ventrite. Third ventrite slightly shorter than the second ventrite. Fourth ventrite about 1.6 times as long as the third ventrite. Fifth ventrite about 0.9 times as long as the fourth ventrite. Metabibial long spur about 2.5 times as long as the short one. Protarsi about subequal in length to the protibiae. Mesotarsi about 1.6 times as long as the mesotibiae. Metatarsi about 1.5 times as long as the metatibiae. Protarsi: first tarsomere about 1.6 times as long as the second tarsomere; second tarsomere about 1.3 times as long as the third tarsomere; fourth tarsomere about 0.5 times as long as the third tarsomere; fifth tarsomere about 3.0 times as long as the fourth tarsomere, subequal to the third and fourth tarsomeres combined. Mesotarsi: first tarsomere about 2.0 times as long as the second tarsomere; second tarsomere about 1.2 times as long as the third tarsomere; fourth tarsomere about 0.8 times as long as the third tarsomere; fifth tarsomere about 2.3 times as long as the fourth tarsomere, subequal to the third and fourth tarsomeres combined. Metatarsi: first tarsomere about 2.1 times as long as the second tarsomere; second tarsomere about 1.2 times as long as the third tarsomere; fourth tarsomere about 0.7 times as long as the third tarsomere; fifth tarsomere about 2.8 times as long as the fourth tarsomere, slightly longer than the third and fourth tarsomeres combined.
Material examined. Holotype—male (ISEA), no. MA2018/4. Paratypes: male (ISEA), no. MA2020/5; male (ISEA), no. MA2020/33.
Etymology. From the Cretaceous period.
Comparison. The new is similar to P. borowskii sp. nov., but differs in the elytra with two longitudinal lobes before the elytral declivity.
Key to species of the genusPoinarinius.
1. Elytra lacking distinct elytral declivity and lobes (
Figure 1A,B,E–G)………………2
— Elytra with more or less distinct elytral declivity (
Figure 2A–D,F–H,
Figure 3A,B,D,E, and
Figure 4A,B,D,F)……………………………………………………………………………..3
2. Pronotum punctate (
Figure 1A,B,E,F)……………………………..
P.
aristovi sp. nov.— Pronotum transverse rugose (
Figure 1G)……………………
P.
antonkozlovi sp. nov.4. Elytra with two transverse lobes before elytral declivity (
Figure 3D and
Figure 4F)…………………………………………………………………………
P.
lesnei sp. nov.— Elytra with two spines before elytral declivity (
Figure 2A)…..
P.
perkovskyi sp. nov.5. Elytra with distinct elytral declivity, without lobes (
Figure 2B–D,F–H)………….6
— Elytra with weak elytral declivity, but with lobes or spines (
Figure 3A,B,E and
Figure 4A,B,D)………………………………………………………………………………………7
6. Body smaller (1.7–2.7 mm). Edges of elytral declivity serrated, lower (
Figure 2B–D,F)………………………………………………………………………..
P.
aladelicatus— Body larger (3.3 mm). Edges of elytral declivity even, higher (
Figure 2G,H)………………………………………………………………….
P.
zahradniki sp. nov.7. Pronotum transverse rugose. Elytra with longitudinal lobes on the sides of very weak elytral declivity…………………………………………………………P. burmaensis
— Pronotum punctate……………………………………………………………………..8
8. Elytra with double spines on sides of elytral declivity (
Figure 3A,B,E)…………………………………………………………………
P.
borowskii sp. nov.—Elytra with two longitudinal lobes before elytral declivity (
Figure 4A,B,D)…………………………………………………………………
P.
cretaceus sp. nov.
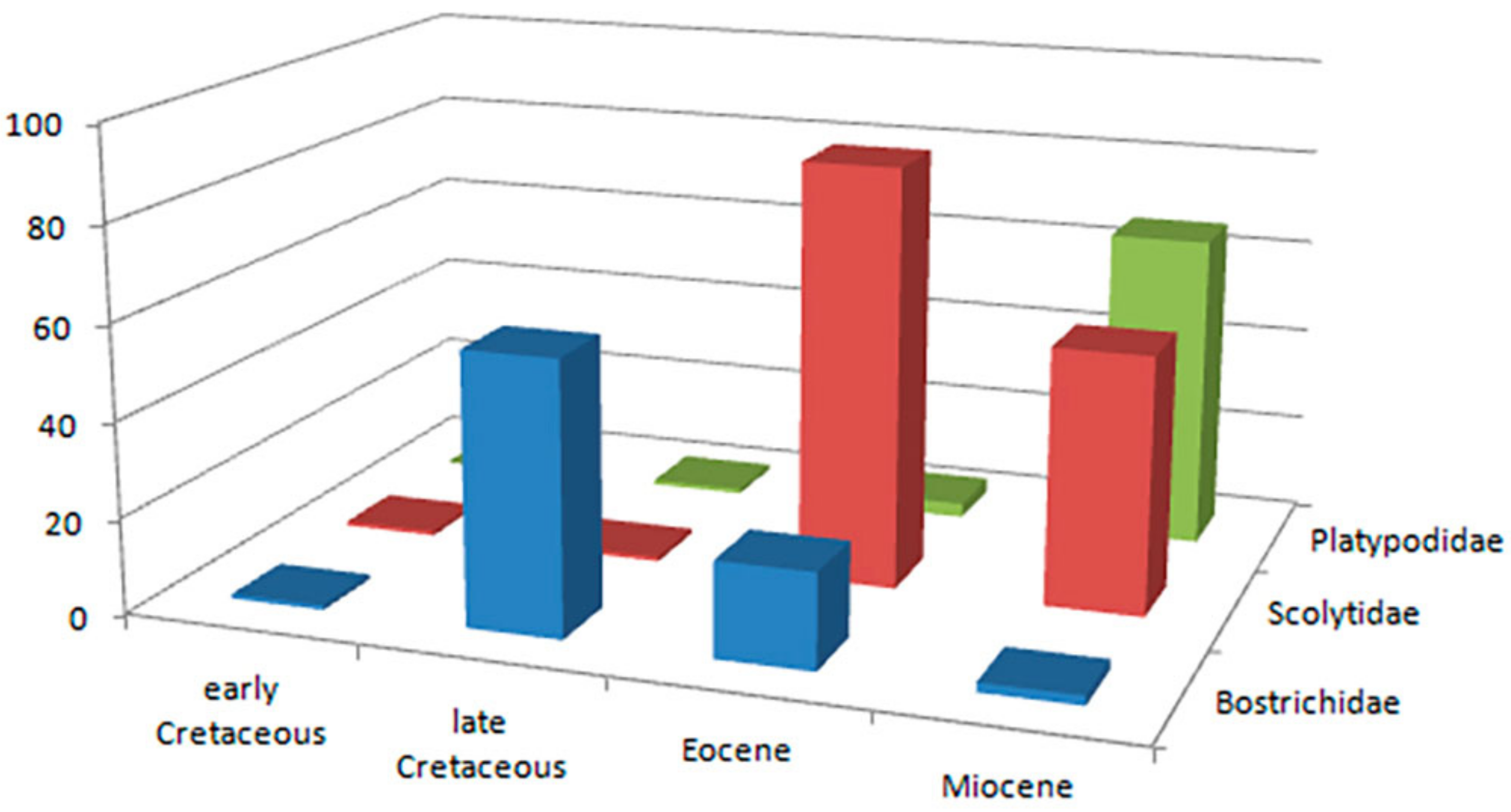
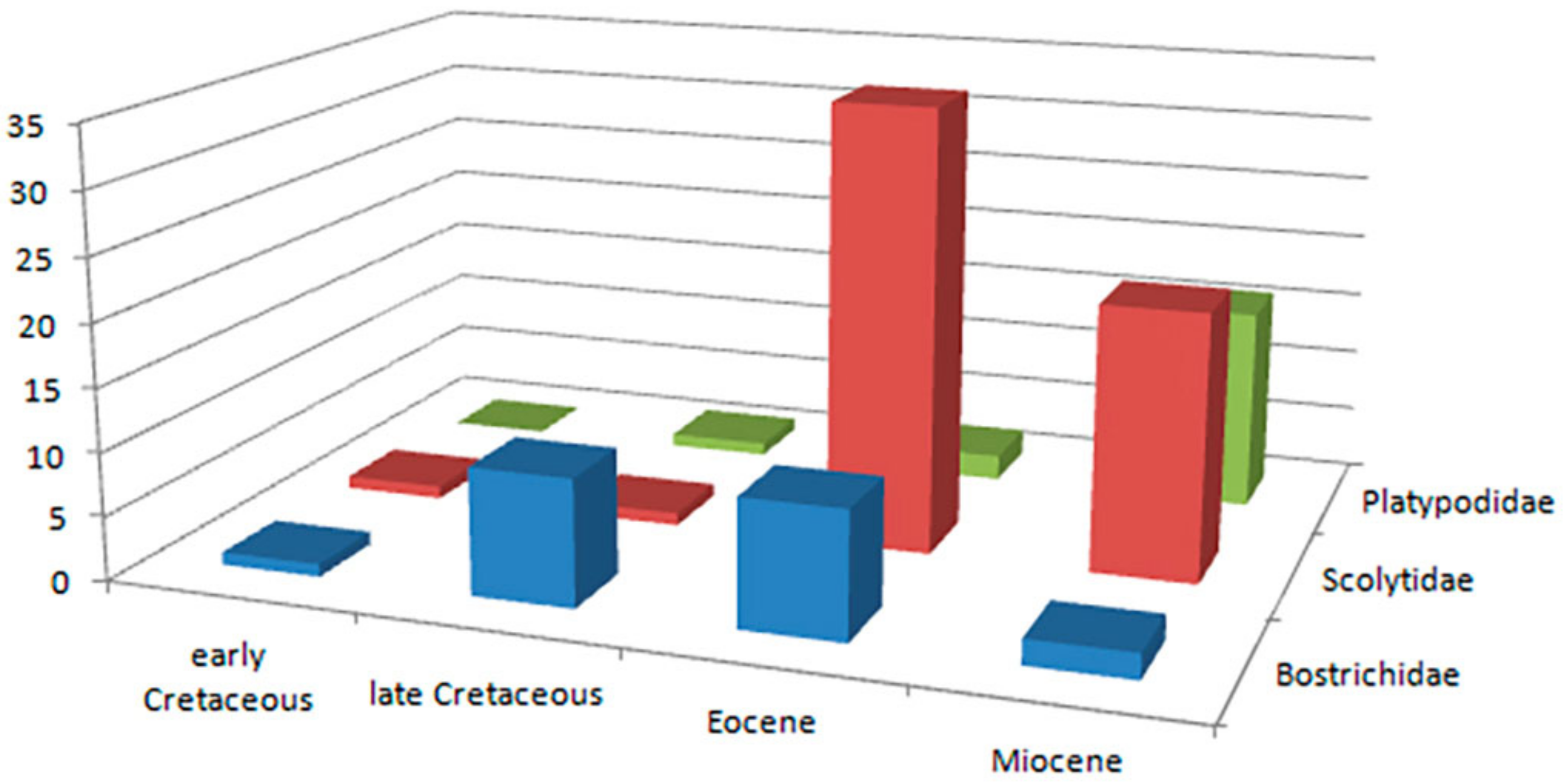
List of the fossil Bostrichidae
Polycaoninae.
Genus Cretolgus Legalov & Háva, 2020.
C.
minimus Legalov & Háva, 2020—Burmese amber, 1 ex. [
9].
Dinoderinae.
Genus Stephanopachys Waterhouse, 1888.
S.
vetus Peris, Delclòs et Perrichot, 2014—French amber, 1 ex. [
7].
S.
electron Zahradník & Háva, 2015—Baltic amber, 1 ex. [
11].
S.
ambericus Zahradník & Háva, 2015—Baltic amber, 1 ex. [
11].
Genus Dinoderus Stephens, 1830.
D.
cuneicollis Wickham, 1913—Florissant, 1 ex. [
14].
Genus? Rhizopertha Stephens, 1830.
Rh. sp.—Baltic amber, 3 ex. [
40].
Genus ? Prostephanus Lesne, 1898.
P. sp.—Mexican amber, 1 ex. [
16].
Alitrepaninae Peng, Jiang, Engel & Wang, 2022.
Genus Poinarinius Legalov, 2018.
P. aladelicatus (Peng, Jiang, Engel & Wang, 2022)—Burmese amber, 19 ex. [
10] and presented data.
P. antonkozlovi Legalov & Háva, sp. n.—Burmese amber, 3 ex.
P. aristovi Legalov & Háva, sp. n.—Burmese amber, 19 ex.
P. borowskii Legalov & Háva, sp. n.—Burmese amber, 3 ex.
P. burmaensis Legalov, 2018—Burmese amber, 4 ex.
P. cretaceus Legalov & Háva, sp. n.—Burmese amber, 3 ex.
P. lesnei Legalov & Háva, sp. n.—Burmese amber, 3 ex.
P. perkovskyi Legalov & Háva, sp. n.—Burmese amber, 1 ex.
P. zahradniki Legalov & Háva, sp. n.—Burmese amber, 1 ex.
Lyctinae.
Genus?Lyctus Fabricius, 1792.
L. sp.—Baltic amber? ex. [
41].
Bostrichinae.
Bostrichini.
Genus ? Bostrichus O.F. Mueller, 1764.
B. sp.—Baltic amber? ex. [
41,
42,
43].
Genus Amphicerus LeConte, 1861.
A.
sublevis Wickham, 1914—Florissant, 1 ex. [
15].
Genus Discoclavata Poinar, 2013.
D.
dominicana Poinar, 2013—Dominican amber, 1 ex. [
17].
Apatini.
Genus?Apate Fabricius 1775.
A. sp.—Baltic amber, ? ex. [
42,
43].
Genus Protapate Wickham, 1912.
P.
contorta Wickham, 1912—Florissant, 1 ex. [
13].
Xyloperthini.
Genus Xylobiops Casey, 1898.
X.
lacustre Wickham, 1912—Florissant, 2 ex. [
13,
14].




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





