
Are Sunken Warships Biodiversity Havens for Corals?

 , ,
, , (This article belongs to the Section Marine Diversity)
Abstract
:1. Introduction
2. Materials and Methods
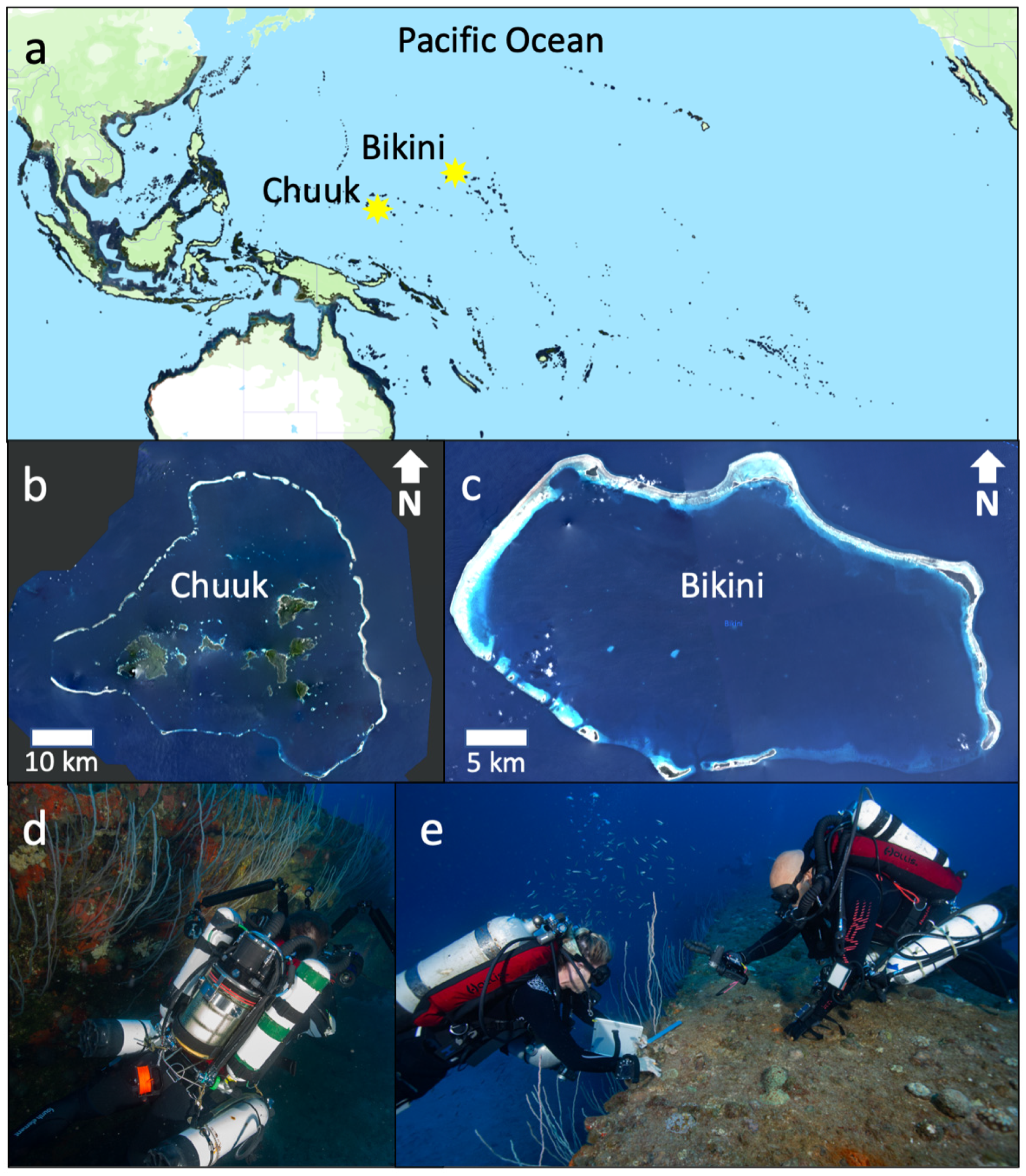
2.1. Field Sites
2.2. Biodiversity Surveys
2.3. Image Analysis
2.4. Data Analysis
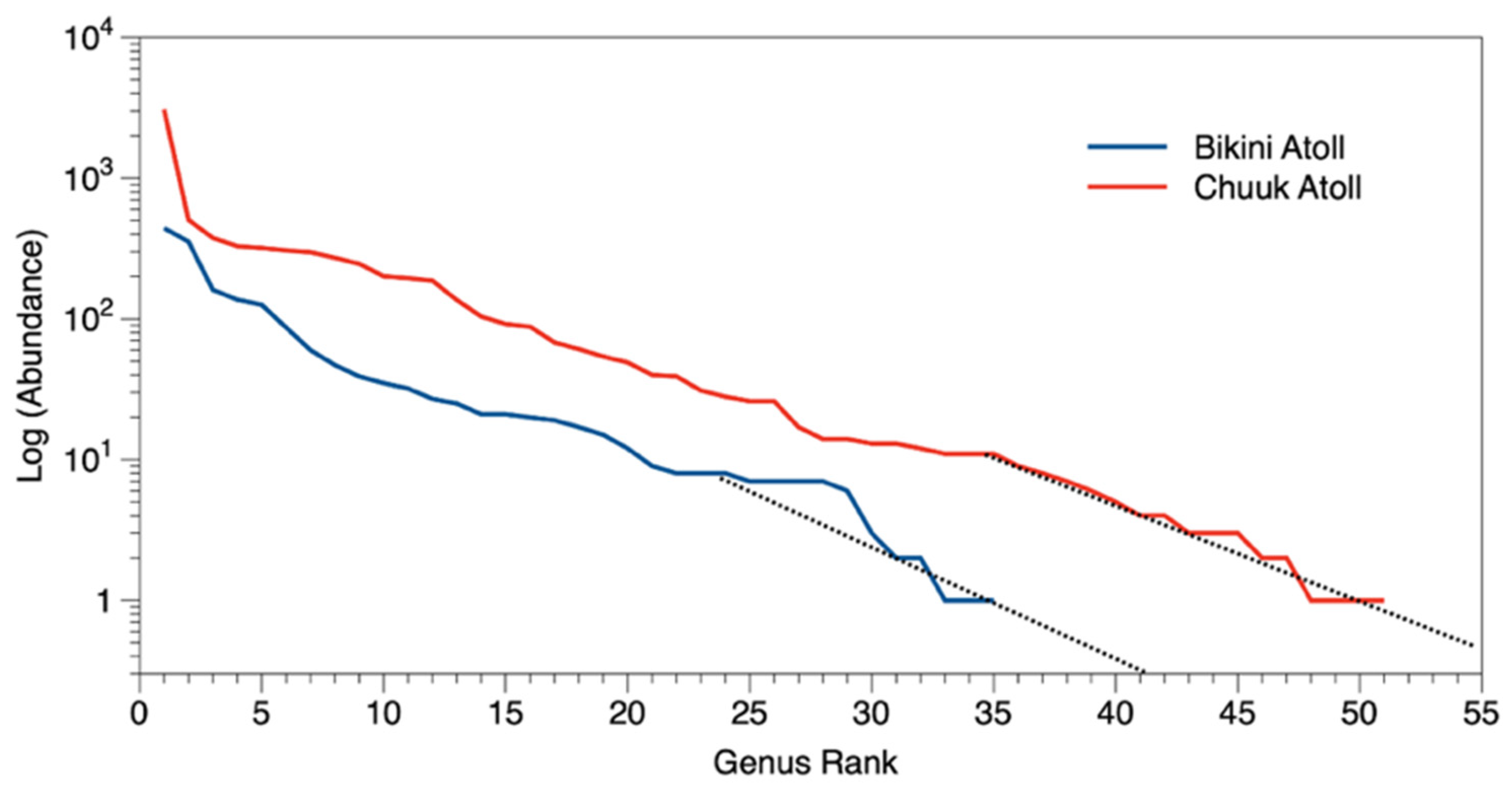
3. Results
3.1. Coral Diversity
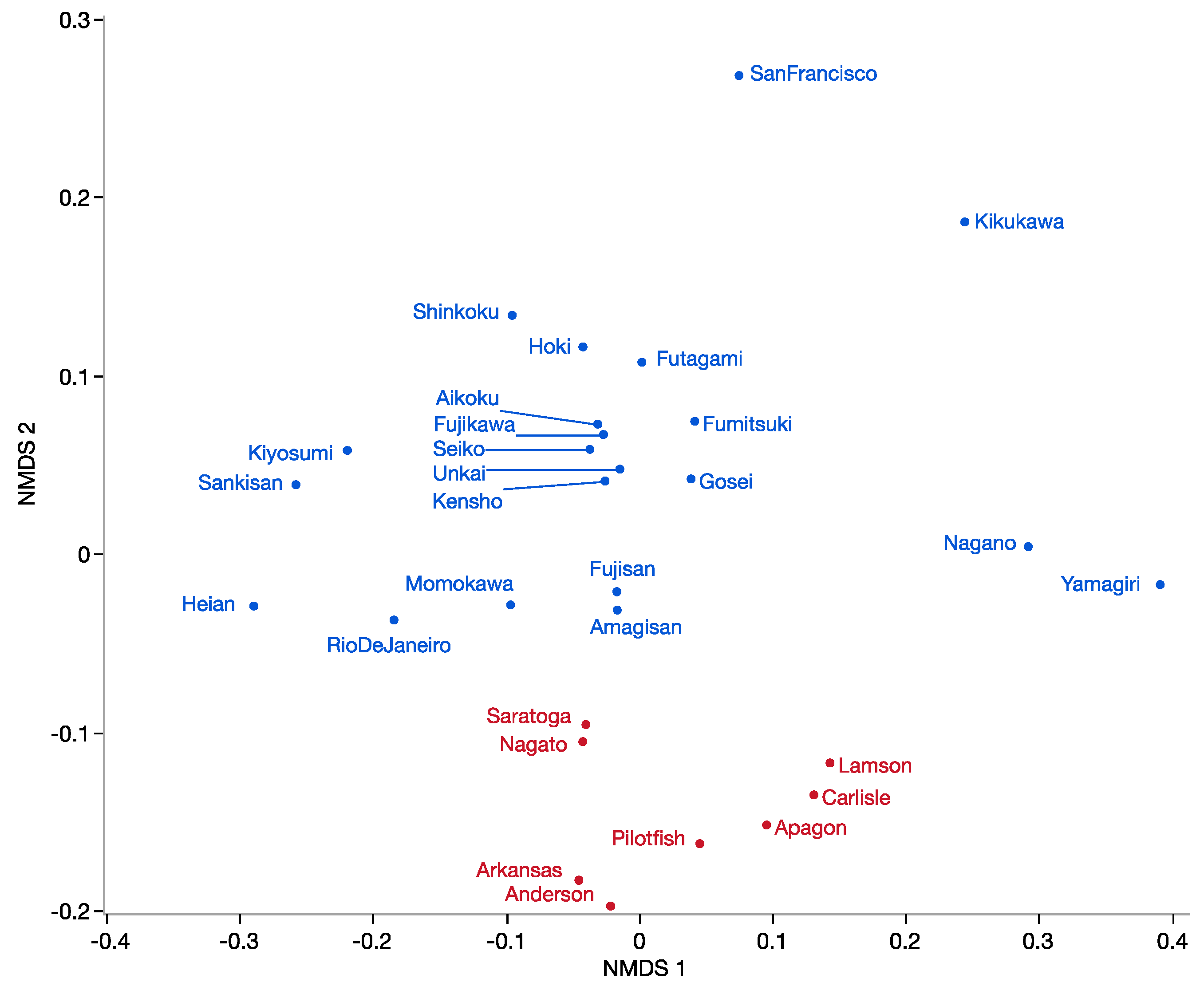
3.2. Ship-Coral Relationships
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SHIP NAME | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| GENUS | Anderson | Apogon | Arkansas | Carlisle | Lamson | Nagato | Pilotfish | Saratoga | Total Abundance by Genus |
| Acanthastrea | 1 | 1 | 3 | 1 | 1 | 8 | 0 | 2 | 17 |
| Acropora | 5 | 3 | 1 | 9 | 7 | 3 | 1 | 18 | 47 |
| Astreopora | 0 | 1 | 8 | 1 | 2 | 17 | 1 | 30 | 60 |
| Cynaria | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cyphastrea | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| Echinophyllia | 1 | 4 | 3 | 6 | 2 | 1 | 1 | 1 | 19 |
| Euphyllia | 1 | 2 | 1 | 1 | 1 | 0 | 1 | 1 | 8 |
| Favia | 84 | 37 | 132 | 22 | 15 | 53 | 70 | 29 | 442 |
| Favites | 41 | 5 | 36 | 6 | 4 | 24 | 27 | 18 | 161 |
| Fungia | 0 | 11 | 2 | 1 | 1 | 1 | 3 | 2 | 21 |
| Galaxea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Goniastrea | 9 | 17 | 19 | 18 | 5 | 9 | 3 | 7 | 87 |
| Goniopora | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 7 |
| Heliofungia | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Herpolitha | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 1 | 15 |
| Hydnophora | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 9 |
| Leptastrea | 29 | 5 | 43 | 6 | 2 | 26 | 1 | 14 | 126 |
| Leptoseris | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 3 |
| Lithophyllon | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 |
| Lobophyllia | 3 | 1 | 7 | 1 | 1 | 4 | 0 | 4 | 21 |
| Merulina | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 2 |
| Montastrea | 63 | 11 | 74 | 3 | 19 | 90 | 11 | 83 | 354 |
| Montipora | 1 | 1 | 1 | 1 | 1 | 9 | 0 | 6 | 20 |
| Oulophyllia | 0 | 0 | 1 | 0 | 0 | 3 | 1 | 2 | 7 |
| Pavona | 0 | 1 | 3 | 2 | 1 | 0 | 0 | 1 | 8 |
| Plerogyra | 1 | 1 | 1 | 2 | 12 | 3 | 1 | 4 | 25 |
| Pocillopora | 1 | 1 | 1 | 1 | 1 | 6 | 1 | 27 | 39 |
| Polyphyllia | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Porites | 1 | 1 | 13 | 1 | 4 | 54 | 3 | 60 | 137 |
| Psammocora | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 7 |
| Scolymia | 14 | 0 | 12 | 0 | 1 | 2 | 1 | 2 | 32 |
| Stylocoeniella | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 2 |
| Symphyllia | 7 | 2 | 1 | 0 | 1 | 13 | 0 | 3 | 27 |
| Tubastrea | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 4 | 7 |
| Turbinaria | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 4 | 6 |
| UND | 8 | 3 | 8 | 3 | 5 | 5 | 8 | 7 | 47 |
| Total Individuals by Ship | 275 | 116 | 377 | 92 | 93 | 338 | 147 | 336 | 1774 |
Appendix B
| SHIP NAME | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GENUS | Rio de Janeiro | Heian | Fumitsuki | momokawa | Seiko | Hoki | Kensho | Fujikawa Combined | Sankisan | Shinkoku | Yamagiri | San Francisco Combined | Kiyosumi Combined | Aikoku | Amagisan | Gosei | Fujisan | Unkai | Nagano | Futagami | Kikukawa | Total Abundance By Genus |
| Acropora | 6 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 9 | 7 | 1 | 0 | 1 | 0 | 3 | 3 | 1 | 4 | 0 | 1 | 0 | 40 |
| Astreopora | 20 | 0 | 0 | 4 | 2 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 31 |
| Cycloseris | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Cynarina | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 7 |
| Diploastrea | 5 | 4 | 1 | 11 | 11 | 11 | 4 | 6 | 15 | 18 | 101 | 0 | 49 | 6 | 37 | 2 | 14 | 2 | 0 | 0 | 1 | 298 |
| Distichopora | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 5 |
| Duncanopsammia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 305 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 16 | 8 | 0 | 0 | 0 | 329 |
| Echinophyllia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 11 |
| Echinopora | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 5 | 0 | 6 | 0 | 0 | 14 |
| Euphyllia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Favia | 15 | 56 | 11 | 92 | 13 | 3 | 22 | 3 | 28 | 0 | 2 | 4 | 9 | 4 | 32 | 4 | 51 | 14 | 12 | 0 | 1 | 376 |
| Favites | 29 | 131 | 14 | 78 | 9 | 7 | 14 | 15 | 35 | 14 | 12 | 0 | 24 | 15 | 37 | 28 | 24 | 9 | 4 | 5 | 2 | 506 |
| Fungia | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 5 | 5 | 23 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 39 |
| Galaxea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 18 | 0 | 1 | 0 | 0 | 0 | 1 | 8 | 0 | 0 | 0 | 0 | 0 | 28 |
| Gardineroseris | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 13 |
| Goniastrea | 32 | 33 | 5 | 32 | 17 | 10 | 3 | 11 | 24 | 1 | 1 | 1 | 8 | 7 | 37 | 40 | 15 | 14 | 3 | 8 | 5 | 307 |
| Goniopora | 0 | 1 | 12 | 0 | 8 | 6 | 8 | 1 | 19 | 36 | 2 | 27 | 8 | 4 | 7 | 1 | 43 | 4 | 14 | 0 | 0 | 201 |
| Heliofungia | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| Herpolitha | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 15 | 4 | 22 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 54 |
| Hydnophora | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| Leptastrea | 169 | 3 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 187 |
| Leptoria | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Leptoseris | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 7 | 1 | 0 | 0 | 13 |
| Lithophyllon | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 |
| Lobophyllia | 19 | 38 | 13 | 34 | 28 | 9 | 27 | 23 | 32 | 59 | 0 | 0 | 17 | 2 | 2 | 0 | 5 | 9 | 2 | 0 | 0 | 319 |
| Merulina | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 |
| Montastrea | 17 | 18 | 4 | 31 | 8 | 1 | 11 | 4 | 8 | 0 | 0 | 2 | 7 | 6 | 27 | 4 | 28 | 9 | 1 | 8 | 1 | 195 |
| Montipora | 6 | 3 | 0 | 1 | 0 | 0 | 1 | 1 | 37 | 1 | 2 | 0 | 3 | 0 | 1 | 1 | 0 | 0 | 4 | 0 | 0 | 61 |
| Moseleya | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 |
| Mycedium | 0 | 0 | 0 | 0 | 4 | 5 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 11 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 26 |
| Oulophyllia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 4 |
| Oxypora | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 3 |
| Pachyseris | 0 | 3 | 0 | 0 | 4 | 10 | 2 | 7 | 15 | 5 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 49 |
| Pavona | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| Pectinia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| Physogyra | 0 | 0 | 3 | 0 | 8 | 5 | 3 | 0 | 42 | 4 | 1 | 3 | 0 | 8 | 4 | 1 | 15 | 6 | 0 | 1 | 0 | 104 |
| Platygyra | 2 | 56 | 0 | 9 | 6 | 0 | 2 | 0 | 5 | 1 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 88 |
| Plerogyra | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 0 | 0 | 4 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 14 |
| Pocillopora | 57 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 2 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 68 |
| Polyphyllia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Porites | 235 | 961 | 41 | 120 | 90 | 102 | 71 | 77 | 363 | 102 | 0 | 21 | 498 | 94 | 45 | 43 | 43 | 87 | 0 | 82 | 6 | 3081 |
| Psammocora | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 118 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 114 | 0 | 0 | 246 |
| Scolymia | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 4 | 0 | 2 | 0 | 0 | 0 | 0 | 12 |
| Seriotopora | 2 | 14 | 2 | 19 | 10 | 1 | 19 | 9 | 100 | 20 | 0 | 0 | 4 | 14 | 25 | 7 | 14 | 7 | 5 | 0 | 0 | 272 |
| Stylaster | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Stylophora | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 11 |
| Symphyllia | 10 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 |
| Trachyphyllia | 2 | 3 | 0 | 1 | 1 | 0 | 0 | 1 | 8 | 8 | 6 | 0 | 0 | 2 | 28 | 5 | 16 | 8 | 3 | 0 | 0 | 92 |
| Tubastrea | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 0 | 3 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 |
| Turbinaria | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 9 |
| UND | 14 | 15 | 1 | 2 | 8 | 1 | 4 | 11 | 6 | 3 | 3 | 0 | 13 | 4 | 9 | 0 | 1 | 10 | 12 | 10 | 9 | 136 |
| Zoopilus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 26 |
| Total Abundance By Ship | 664 | 1343 | 111 | 438 | 228 | 174 | 195 | 193 | 1109 | 356 | 273 | 63 | 657 | 221 | 316 | 150 | 302 | 207 | 193 | 117 | 25 | 7335 |
References
- Hoegh-Guldberg, O. Coral reefs in the Anthropocene: Persistence or the end of the line? Geol. Soc. Lond. Spec. Publ. 2014, 395, 167–183. [Google Scholar] [CrossRef]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.C.; Kleypas, J.; van de Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, N. The future of coral reefs. Proc. Natl. Acad. Sci. USA 2001, 98, 5419–5425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandolfi, J.M.; Kiessling, W. Gaining insights from past reefs to inform understanding of coral reef response to global climate change. Curr. Opin. Environ. Sustain. 2014, 7, 52–58. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. The adaptation of coral reefs to climate change: Is the Red Queen being outpaced? Sci. Mar. 2012, 76, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Matz, M.V.; Treml, E.A.; Aglyamova, G.V.; Bay, L.K. Potential and limits for rapid genetic adaptation to warming in a Great Barrier Reef coral. PLoS Genet. 2018, 14, e1007220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claudet, J.; Pelletier, D. Marine protected areas and artificial reefs: A review of the interactions between management and scientific studies. Aquat. Living Resour. 2004, 17, 129–138. [Google Scholar] [CrossRef]
- Ilieva, I.; Jouvet, L.; Seidelin, L.; Best, B.D.; Aldabet, S.; da Silva, R.; Conde, D.A. A global database of intentionally deployed wrecks to serve as artificial reefs. Data Brief 2019, 23, 103584. [Google Scholar] [CrossRef] [PubMed]
- Hynes, M.V.; Peters, J.E.; Rushworth, D. Artificial Reefs: A Disposal Option for Navy and MARAD Ships; DB-391-NAVY.; National Defense Research Institute: Arlington, VA, USA, 2004; p. 57. [Google Scholar]
- Pinheiro, H.T.; Bernardi, G.; Simon, T.; Joyeux, J.C.; Macieira, R.M.; Gasparini, J.L.; Rocha, C.; Rocha, L.A. Island biogeography of marine organisms. Nature 2017, 549, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Burt, J.; Bartholomew, A.; Usseglio, P.; Bauman, A.; Sale, P.F. Are artificial reefs surrogates of natural habitats for corals and fish in Dubai, United Arab Emirates? Coral Reefs 2009, 28, 663–675. [Google Scholar] [CrossRef]
- Perkol-Finkel, S.; Shashar, N.; Benayahu, Y. Can artificial reefs mimic natural reef communities? The roles of structural features and age. Mar. Environ. Res. 2006, 61, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, J.A.; Sutherland, D.L. Artificial reef research: A review with recommendations for future priorities. Bull. Mar. Sci. 1985, 37, 11–39. [Google Scholar]
- Preston, F.W. Time and space and the variation of species. Ecology 1960, 41, 611–627. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Smith, T.B.; Glynn, P.W.; Maté, J.L.; Toth, L.T.; Gyory, J. A depth refugium from catastrophic coral bleaching prevents regional extinction. Ecology 2014, 95, 1663–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baird, A.H.; Madin, J.S.; Álvarez-Noriega, M.; Fontoura, L.; Kerry, J.T.; Kuo, C.-Y.; Precoda, K.; Torres-Pulliza, D.; Woods, R.M.; Zawada, K.J.A. A decline in bleaching suggests that depth can provide a refuge from global warming in most coral taxa. Mar. Ecol. Prog. Ser. 2018, 603, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Venegas, R.M.; Oliver, T.; Liu, G.; Heron, S.F.; Clark, S.J.; Pomeroy, N.; Young, C.; Eakin, C.M.; Brainard, R.E. The rarity of depth refugia from coral bleaching heat stress in the western and central Pacific Islands. Sci. Rep. 2019, 9, 19710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainger, R. Science at the crossroads: The Navy, Bikini Atoll, and American oceanography in the 1940s. Hist. Stud. Phys. Biol. Sci. 2000, 30, 349–371. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.W. Recent Corals of the Marshall Islands. Bikini and Nearby Atolls, Part 2, Oceanography; US Government Printing Office: Washington, DC, USA, 1954.
- Richards, Z.T.; Beger, M.; Pinca, S.; Wallace, C.C. Bikini Atoll coral biodiversity resilience five decades after nuclear testing. Mar. Pollut. Bull. 2008, 56, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Veron, J.E.N. Corals in Space and Time: The Biogeography and Evolution of the Scleractinia; Cornell University Press: Ithaca, NY, USA, 1995. [Google Scholar]
- Veron, J.E.N. Corals of the World [Three Volumes]; Australian Institute of Marine Science: Townsville, QLD, Australia, 2000.
- Oliver, E.C.J.; Donat, M.G.; Burrows, M.T.; Moore, P.J.; Smale, D.A.; Alexander, L.V.; Benthuysen, J.A.; Feng, M.; Sen Gupta, A.; Hobday, A.J.; et al. Longer and more frequent marine heatwaves over the past century. Nat. Commun. 2018, 9, 1324. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Ospina, A.; Kendrick, G.A.; Hovey, R.K. Depth moderates loss of marine foundation species after an extreme marine heatwave: Could deep temperate reefs act as a refuge? Proc. R. Soc. B 2020, 287, 20200709. [Google Scholar] [CrossRef] [PubMed]



| Ship Name | Length (m) | Survey Depth (m) |
|---|---|---|
| Bikini Atoll | ||
| Anderson | 106 | 49 |
| Apogon | 95 | 48 |
| Arkansas | 171 | 44 |
| Carlisle | 130 | 48 |
| Lamson | 104 | 52 |
| Nagato | 221 | 42 |
| Pilotfish | 95 | 52 |
| Saratoga | 268 | 50 |
| Chuuk Atoll | ||
| Aikoku | 161 | 56 |
| Amagisan | 137 | 45 |
| Fujikawa | 133 | 29 |
| Fujisan | 156 | 56 |
| Fumitsuki | 101 | 29 |
| Futagami | 40 | 29 |
| Gosei | 82 | 29 |
| Heian | 155 | 29 |
| Hoki | 137 | 45 |
| Kensho | 116 | 29 |
| Kikukawa | 108 | 29 |
| Kiyosumi | 137 | 29 |
| Momokawa | 107 | 45 |
| Nagano | 105 | 66 |
| Rio de Janeiro | 137 | 29 |
| San Francisco | 117 | 56 |
| Sankisan | 113 | 29 |
| Seiko | 120 | 56 |
| Shinkoku | 152 | 29 |
| Unkai | 93 | 29 |
| Yamagiri | 134 | 29 |
| Ship Name | Richness | Rel. Richness (%) | Total Abundance | Rel. Abundance (%) |
|---|---|---|---|---|
| Bikini Atoll | ||||
| Anderson | 22 | 64.7 | 275 | 15.5 |
| Apogon | 27 | 79.4 | 116 | 6.5 |
| Arkansas | 28 | 82.4 | 377 | 21.3 |
| Carlisle | 25 | 73.5 | 92 | 5.2 |
| Lamson | 27 | 79.4 | 93 | 5.2 |
| Nagato | 26 | 76.5 | 338 | 19.1 |
| Pilotfish | 22 | 64.7 | 147 | 8.3 |
| Saratoga | 29 | 85.3 | 336 | 18.9 |
| Chuuk Atoll | ||||
| Aikoku | 28 | 54.9 | 221 | 3.0 |
| Amagisan | 23 | 45.1 | 316 | 4.3 |
| Fujikawa | 19 | 37.3 | 193 | 2.6 |
| Fujisan | 23 | 45.1 | 302 | 4.1 |
| Fumitsuki | 14 | 27.5 | 111 | 1.5 |
| Futagami | 9 | 17.6 | 117 | 1.6 |
| Gosei | 16 | 31.4 | 150 | 2.0 |
| Heian | 19 | 37.3 | 1343 | 18.3 |
| Hoki | 15 | 29.4 | 174 | 2.4 |
| Kensho | 17 | 33.3 | 195 | 2.7 |
| Kikukawa | 7 | 13.7 | 25 | 0.3 |
| Kiyosumi | 19 | 37.3 | 657 | 9.0 |
| Momokawa | 16 | 31.4 | 438 | 6.0 |
| Nagano | 17 | 33.3 | 193 | 2.6 |
| Rio de Janeiro | 22 | 43.1 | 664 | 9.1 |
| San Francisco | 8 | 15.7 | 63 | 0.9 |
| Sankisan | 29 | 56.9 | 1109 | 15.1 |
| Seiko | 17 | 33.3 | 228 | 3.1 |
| Shinkoku | 21 | 41.2 | 356 | 4.9 |
| Unkai | 19 | 37.3 | 207 | 2.8 |
| Yamagiri | 17 | 33.3 | 273 | 3.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asner, G.P.; Giardina, S.F.; Balzotti, C.; Drury, C.; Hopson, S.; Martin, R.E. Are Sunken Warships Biodiversity Havens for Corals? Diversity 2022, 14, 139. https://doi.org/10.3390/d14020139
Asner GP, Giardina SF, Balzotti C, Drury C, Hopson S, Martin RE. Are Sunken Warships Biodiversity Havens for Corals? Diversity. 2022; 14(2):139. https://doi.org/10.3390/d14020139
Chicago/Turabian StyleAsner, Gregory P., Sonja F. Giardina, Christopher Balzotti, Crawford Drury, Sean Hopson, and Roberta E. Martin. 2022. "Are Sunken Warships Biodiversity Havens for Corals?" Diversity 14, no. 2: 139. https://doi.org/10.3390/d14020139
APA StyleAsner, G. P., Giardina, S. F., Balzotti, C., Drury, C., Hopson, S., & Martin, R. E. (2022). Are Sunken Warships Biodiversity Havens for Corals? Diversity, 14(2), 139. https://doi.org/10.3390/d14020139