
Re-Examination of the Phylogenetic Relationship among Merulinidae Subclades in Non-Reefal Coral Communities of Northeastern Taiwan

{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Uncovering Taxonomic Progress
1.2. Revision of Merulinidae (Clade XVII)
1.3. Taiwan Taxonomy and Species Diversity
1.4. Purpose of This Research
2. Materials and Methods
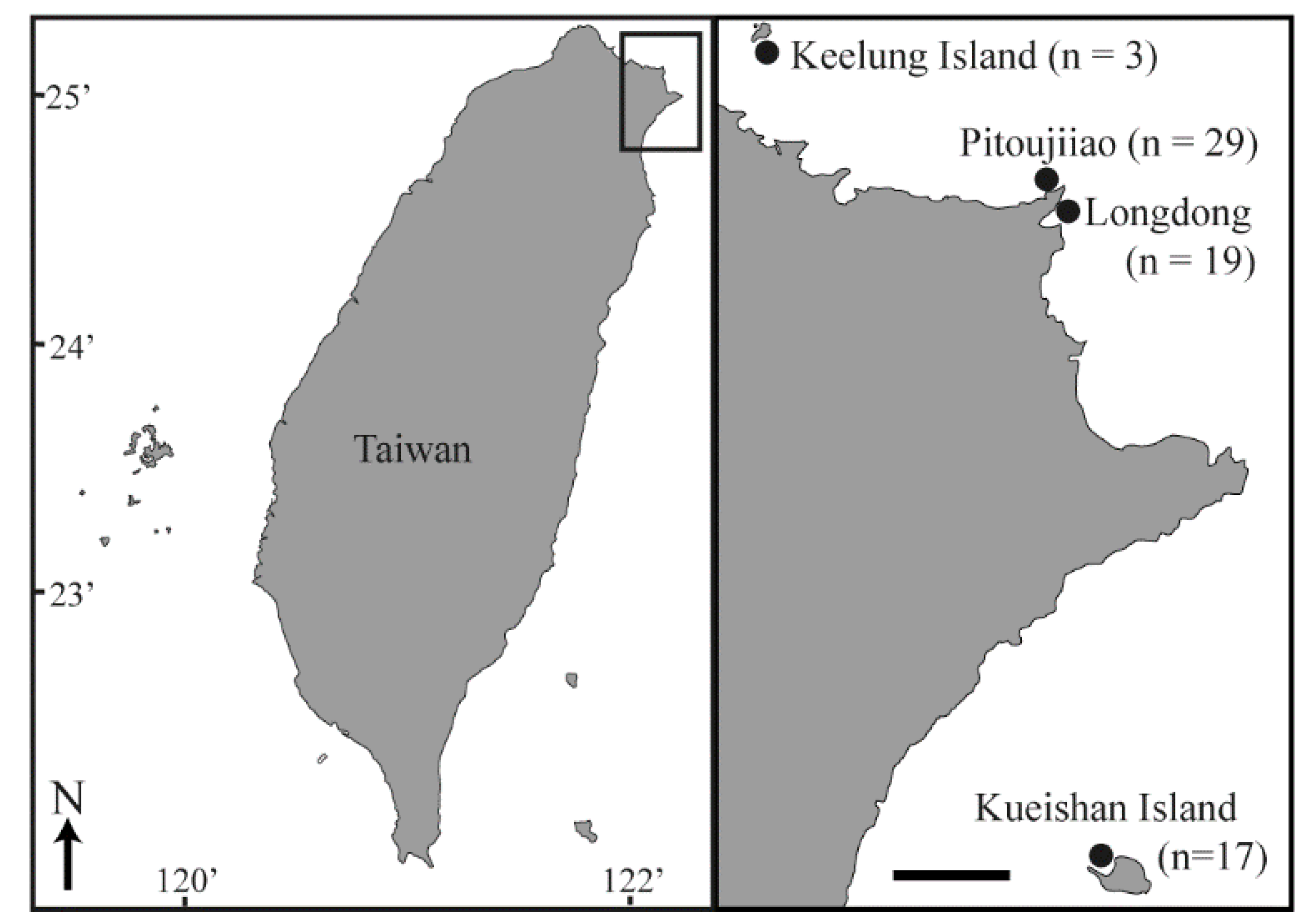
2.1. Sample Collection
2.2. Species Identification
2.3. Sequence Management, Alignment, and Matrix
2.4. Molecular Datasets
2.5. Molecular Phylogenetic Analysis
3. Results
3.1. Characteristics of the Gene Data
3.2. Results of the Analysis Matrix
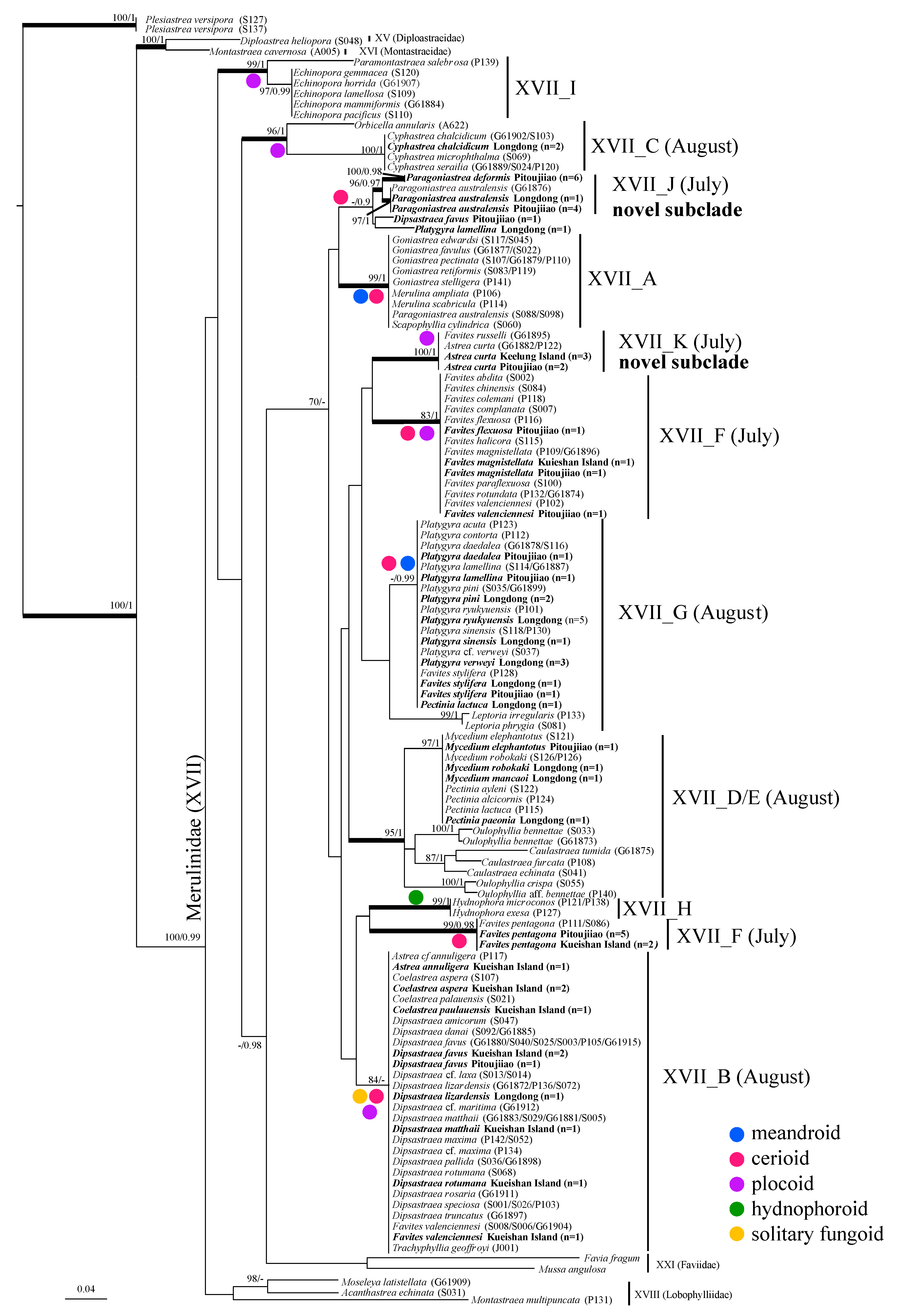
3.3. Phylogenetic Relationship
3.4. The Phylogenetic Tree
4. Discussion
4.1. Phylogentic Relationship of Merulinid Subclades
4.2. Application of Molecular Phylogentic Approaches
4.3. Sexual Reproduction in Merulinidae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veron, J.E.N.; Pichon, M. Scleractinia of Eastern Australia; Australian Govt. Pub. Service: Townsville, Australia, 1976; Volume 1. [Google Scholar]
- Veron, J.E.N. Corals of the World; Australian Institute of Marine Science: Townsville, Australia, 2000; Volume 3. [Google Scholar]
- Todd, P.A. Morphological plasticity in scleractinian corals. Biol. Rev. Camb. Philos. Soc. 2008, 83, 315–337. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.W. Scleractinia. In Treatise on Invertebrate Paleontology, Coelenterata; Teichert, C., Ed.; Geological Society of America and University of Kansas Press: Lawrence, KS, USA, 1956; pp. 328–444. [Google Scholar]
- Veron, J.E. Corals in Space and Time: The Biogeography & Evolution of the Scleractinia; Cornell University Press: Ithaca, NY, USA, 1995. [Google Scholar]
- Romano, S.L.; Palumbi, S.R. Evolution of scleractinian corals inferred from molecular systematics. Science 1996, 271, 640–642. [Google Scholar] [CrossRef]
- Romano, S.L.; Palumbi, S.R. Molecular evolution of a portion of the mitochondrial 16S ribosomal gene region in scleractinian corals. J. Mol. Evol. 1997, 45, 397–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitahara, M.V.; Cairns, S.D.; Stolarski, J.; Blair, D.; Miller, D.J. A comprehensive phylogenetic analysis of the scleractinia (Cnidaria, Anthozoa) based on mitochondrial CO1 sequence data. PLoS ONE 2010, 5, 1490. [Google Scholar] [CrossRef]
- Fukami, H.; Chen, C.A.; Budd, A.F.; Collins, A.; Wallace, C.; Chuang, Y.Y.; Chen, C.; Dai, C.F.; Iwao, K.; Sheppard, C.; et al. Mitochondrial and nuclear genes suggest that stony corals are monophyletic but most families of stony corals are not (Order Scleractinia, Class Anthozoa, Phylum Cnidaria). PLoS ONE 2008, 3, e3222. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Benzoni, F.; Fukami, H.; Knowlton, N.; Smith, N.D.; Budd, A.F. Taxonomic classification of the reef coral families Merulinidae, Montastraeidae, and Diploastraeidae (Cnidaria: Anthozoa: Scleractinia). Zool. J. Linnean Soc. 2014, 171, 277–355. [Google Scholar] [CrossRef]
- Kuzon, K.S.; Lin, M.F.; Lagman, M.C.A.A.; Licuanan, W.R.Y.; Chen, C.A. Resurrecting a subgenus to genus: Molecular phylogeny of Euphyllia and Fimbriaphyllia (order Scleractinia; family Euphyllidae; clade V). PeerJ 2017, 5, e4074. [Google Scholar]
- Benzoni, F.; Arrigoni, R.; Waheed, Z.; Stefani, F.; Hoeksema, B.W. Phylogenetic relationships and revision of the genus Blastomussa (Cnidaria: Anthozoa: Scleractinia) with description of a new species. Baffles Bull. Zool. 2014, 62, 358–378. [Google Scholar]
- Budd, A.F. Systematics and evolution of scleractinian corals. In Encyclopedia of Life Synthesis Meeting Report; National Museum of Natural History: Washington, DC, USA, 2009. [Google Scholar]
- Budd, A.F.; Stolarski, J. Corallite wall and septal microstructure in scleractinian reef corals: Comparison of molecular clades within the family Faviidae. J. Morphol. 2011, 272, 66–88. [Google Scholar] [CrossRef]
- Huang, D.; Benzoni, F.; Arrigoni, R.; Baird, A.H.; Berumen, M.L.; Bouwmeester, J.; Chou, L.M.; Fukami, H.; Licuanan, W.Y.; Lovell, E.R.; et al. Towards a phylogenetic classification of reef corals: The Indo-Pacific genera Merulina, Goniastrea and Scapophyllia (Scleractinia, Merulinidae). Zool. Scr. 2014, 43, 531–548. [Google Scholar] [CrossRef]
- Huang, D.; Licuanan, W.Y.; Baird, A.H.; Fukami, H. Cleaning up the ‘Bigmessidae’: Molecular phylogeny of scleractinian corals from Faviidae, Merulinidae, Pectiniidae and Trachyphylliidae. BMC Evol. Biol. 2011, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Meier, R.; Tood, P.A.; Chou, L.M. More evidence for pervasive paraphyly in scleractinian corals: Systematic study of Southeast Asian Faviidae (Cnidaria; Scleractinia) based on molecular and morphological data. Mol. Phylogenet. Evol. 2009, 50, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Fukami, H.; Budd, A.F.; Paulay, G.; SolIé-Cava, A.; Chen, C.A.; Iwao, K.; Knowlton, N. Conventional taxonomy obsecures deep divergence between Pacific and Atlantic corals. Nature 2004, 427, 832–834. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.A.; Budd, A.F.; Carlon, D.B. Molecules and fossils reveal punctuated diversification in Caribbean “faviid” corals. BMC Evol. Biol. 2012, 12, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World List of Scleractinia. Available online: http://www.marinespecies.org/scleractinia (accessed on 2 August 2021).
- Khalih, H.M.; Fathy, M.S.; Sawy, S.M.A. Quaternary corals (Scleractinia: Merulinidae) from the Egyptian and Saudi Arabian Red Sea Coast. Geol. J. 2021, 56, 4150–4188. [Google Scholar] [CrossRef]
- Veron, J.E.N.; De Vantier, L.M.; Turak, E.; Green, A.L.; Kininmonth, S.; Stafford-Smith, M.; Peterson, N. The Coral Triangle. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 47–55. [Google Scholar]
- Dai, C.F.; Soong, K.; Chen, C.A.; Fan, T.Y.; Hsieh, H.J.; Jeng, M.S.; Chen, C.H.; Horng, S. Status of Coral Reefs of Taiwan; Institute of Oceanography, National Taiwan University: Taipei, Taiwan, 2009. [Google Scholar]
- Ribas-Deulofeu, L.; Denis, V.; Palmass, S.; Kuo, C.Y.; Hsieh, H.; Chen, C. Structure of benthic communities along the Taiwan latitudinal gradient. PLoS ONE 2016, 11, e0160601. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.F.; Cheng, Y.L. Corals of Taiwan; Institute of Oceanography: Taipei, Taiwan, 2020; Volume 1. [Google Scholar]
- Chen, C.A.; Wallace, C.C.; Yu, J.K.; Wei, N.V. Strategies for amplification by polymerase chain reaction of the complete sequence of the gene encoding nuclear large subunit ribosomal RNA in corals. Mar. Biotechnol. 2000, 2, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.F.; Kitahara, M.V.; Tachikawa, H.; Keshavmurthy, S.; Chen, C.A. A new shallow-water species, Polycyathus chaishanensis sp. nov. (Scleractinia: Caryophylliidae), from Chaishan, Kaohsiung, Taiwan. Zool. Stud. 2012, 51, 213–221. [Google Scholar]
- De Palmas, S.; Soto, D.; Denis, V.; Ho, M.J.; Chen, C.A. Molecular assessment of Pocillopora verrucosa (Scleractinia; Pocilloporidae) distribution along a depth gradient in Ludao, Taiwan. PeerJ 2018, 6, e5797. [Google Scholar] [CrossRef] [Green Version]
- Soto, D.; De Palmas, S.; Ho, M.J.; Denis, V.; Chen, C.A. Spatial variation in the morphological traits of Pocillopora verrucosa along a depth gradient in Taiwan. PLoS ONE 2018, 13, e0202586. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; Arrigoni, R. DNA barcoding of a stowaway reef coral in the international aquarium trade results in a new distribution record. Mar. Biodivers. 2020, 50, 41. [Google Scholar] [CrossRef]
- Shikina, S.; Chang, C.F. Sexual Reproduction in Stony Corals and Insight into the Evolution of Oogenesis in Cnidaria. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer: Cham, Switzerland, 2016; pp. 249–268. [Google Scholar]
- Chen, C.J.; Chen, W.J.; Chang, C.F. Multispecies spawning of scleractinian corals in non-reefal coral communities of northern Taiwan in the northwestern Pacific Ocean. Bull. Mar. Sci. 2021, 97, 351–371. [Google Scholar] [CrossRef]
- Lin, Y.V.; Denis, V. Acknowledging differences: Number, characteristics, and distribution of marine benthic communities along Taiwan coast. Ecosphere 2019, 10, e02803. [Google Scholar] [CrossRef]
- Madden, T. The NCBI Handbook; National Center for Biotechnology: Bethesda, MD, USA, 2002. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. Se-Al Sequence Alignment Editor, Version 2.0a11. Available online: http://tree.bio.ed.ac.uk/software/seal/ (accessed on 8 August 2002).
- Sworfford, D.L. PAUP: Phylogenetic Analysis Using Parsimony, Version 3.1; Illinois Natural History Survey: Champaign, IL, USA, 1991. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In 2010 Gateway Computing Environments Workshop (GCE); IEEE: Piscataway, NJ, USA, 2010. [Google Scholar]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhana, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Calcott, B.; Ho, S.Y.; Guindon, S. Partitionfinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.6. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 30 November 2009).
- Rambaut, A. FigTree v1.4.2. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 March 2019).
- Harrison, P.L. Sexual Reproduction of Scleractinian Corals. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 59–85. [Google Scholar]
- Kerr, A.M.; Baird, A.H.; Hughes, T.P. Correlated evolution of sex and reproductive mode in corals (Anthozoa: Scleractinia). Proc. R. Soc. B 2010, 278, 75–81. [Google Scholar] [CrossRef]
- Baird, A.H.; Guest, J.R.; Willis, B.L. Systematic and biogeographical patterns in the reproductive biology of scleractinian coral. Annu. Rev. Ecol. Evol. 2009, 40, 551–571. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.F.; Soong, K.; Fan, T.Y. Sexual reproduction of corals in northern and southern Taiwan. In Proceedings of the International Coral Reef Symposium (ICRS), Guam, Micronesia, 22–27 June 1992; Volume 1, pp. 448–455. [Google Scholar]
- Babcock, R. Reproduction and distribution of two species of Goniastrea (Scleractinia) from the Great Barrier Reef Province. Coral Reefs 1984, 2, 187–195. [Google Scholar]
- Babcock, R.C.; Bull, G.D.; Harrison, P.L.; Heyward, A.J.; Oliver, J.K.; Wallace, C.C.; Willis, B.L. Synchronous spawnings of 105 scleractinian coral species on the Great Barrier Reef. Mar. Biol. 1986, 90, 379–394. [Google Scholar] [CrossRef]
- Harrison, P.L.; Babcock, R.C.; Bull, G.D.; Oliver, J.K.; Wallace, C.C.; Willis, B.L. Mass spawning in tropical reef corals. Science 1984, 223, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Willis, B.L.; Babcock, R.C.; Harrison, P.L.; Oliver, J.K.; Wallace, C.C. Patterns in the mass spawning of corals on the Great Barrier Reef from 1981 to 1984. In Proceedings of the Fifth International Coral Reef Congress, Tahiti, French Polynesia, 27 May–1 June 1985; pp. 343–348. [Google Scholar]
- Abe, N. Postlarval development of the coral Fungia actiniformis var. palawensis Doderlein. Palao Trop. Biol. Stat. Stud. 1937, 1, 73–93. [Google Scholar]
- Motoda, S. Observation of period of extrusion of planula of Goniastrea aspera (Verrill.). Kagaku Nanyo 1939, 1, 5–7. [Google Scholar]
- Sakai, K. Gametogenesis, spawning, and planula brooding by the reef coral Goniastrea aspera (Scleractinia) in Okinawa, Japan. Mar. Ecol. Prog. Ser. 1997, 151, 67–72. [Google Scholar] [CrossRef]
- Heyward, A.K. Sexual reproduction of corals in Okinawa. Galaxea 1987, 6, 331–343. [Google Scholar]
- Hayashibara, T.; Shimoike, K.; Kimura, T.; Hosaka, S.; Heyward, A.; Harrison, P.L.; Kudo, K.; Omori, M. Patterns of coral spawning at Akajima Island, Okinawa, Japan. Mar. Ecol. Prog. Ser. 1993, 101, 253–262. [Google Scholar] [CrossRef]
- Nozawa, Y.; Harrison, P.L. Temporal settlement patterns of larvae of the broadcast spawning reef coral Favites chinensis and the broadcast spawning and brooding reef coral Goniastrea aspera from Okinawa, Japan. Coral Reefs 2005, 24, 274–282. [Google Scholar] [CrossRef]
- Castrillón, A.; Muñoz, C.; Zapata, F. Reproductive patterns of the coral Pocillopora damicornis at Gorgona Island, Colombian Pacific Ocean. Mar. Biol. Res. 2015, 15, 1065–1075. [Google Scholar] [CrossRef]
- Richmond, R.H.; Jokiel, P.L. Lunar periodicity in larva release in the reef coral Pocillopora damicornis at Enewetak and Hawaii. Bull. Mar. Sci. 1984, 34, 280–287. [Google Scholar]
- Stoddart, J.A.; Black, R. Cycles of gametogenesis and planulation in the coral Pocillopora damicornis. Mar. Ecol. Prog. Ser. 1985, 23, 153–164. [Google Scholar] [CrossRef]
- Ward, S. Evidence for broadcast spawning as well as brooding in the scleractinian coral Pocillopora damicornis. Mar. Biol. 1992, 112, 641–646. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-J.; Ho, Y.-Y.; Chang, C.-F. Re-Examination of the Phylogenetic Relationship among Merulinidae Subclades in Non-Reefal Coral Communities of Northeastern Taiwan. Diversity 2022, 14, 144. https://doi.org/10.3390/d14020144
Chen C-J, Ho Y-Y, Chang C-F. Re-Examination of the Phylogenetic Relationship among Merulinidae Subclades in Non-Reefal Coral Communities of Northeastern Taiwan. Diversity. 2022; 14(2):144. https://doi.org/10.3390/d14020144
Chicago/Turabian StyleChen, Chieh-Jhen, Yu-Ying Ho, and Ching-Fong Chang. 2022. "Re-Examination of the Phylogenetic Relationship among Merulinidae Subclades in Non-Reefal Coral Communities of Northeastern Taiwan" Diversity 14, no. 2: 144. https://doi.org/10.3390/d14020144
APA StyleChen, C.-J., Ho, Y.-Y., & Chang, C.-F. (2022). Re-Examination of the Phylogenetic Relationship among Merulinidae Subclades in Non-Reefal Coral Communities of Northeastern Taiwan. Diversity, 14(2), 144. https://doi.org/10.3390/d14020144