
Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity



Abstract
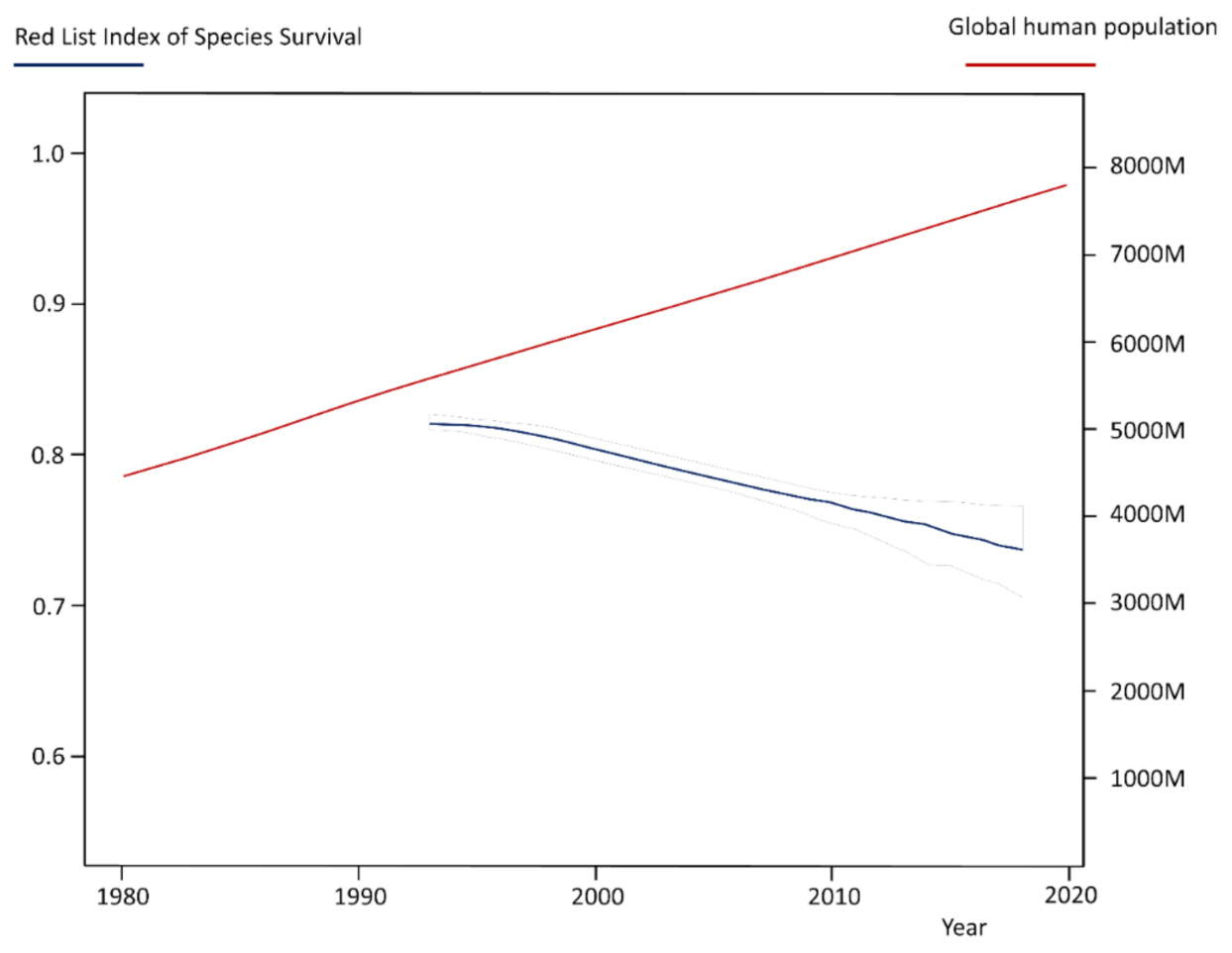
1. Introduction

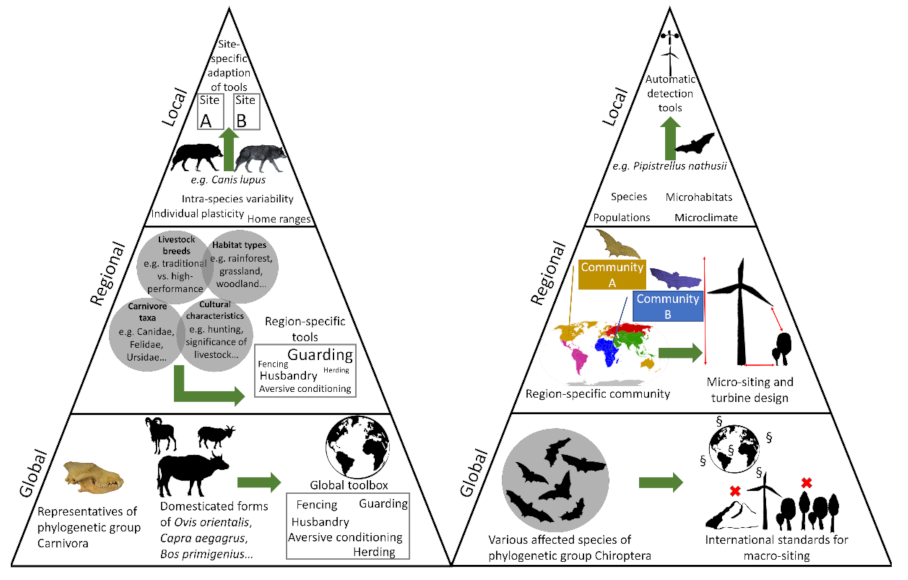
2. Trying to Adopt a Wildlife Species’ Perspective

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Extinct in the Wild | Population Bottleneck | Nucleus of Initial Reintroduction Program (Founders) | Initial Release into Wild | Area of Initial Release | Current In Situ Population [30] |
|---|---|---|---|---|---|---|
| Strigops habroptila | 1997 [31] | 62 animals, 1999 [31] | 61 animals, 1980–1991 [31] | - | Offshore islands of New Zealand [31] | 116 mature individuals |
| Leucopsar rothschildi | - | 6 animals in the wild in 2001 + unknown number in captivity [32] | 426 animals, 1998–2019 [32] | around 1983 [32] | Bali Barat National Park [32] | 1–49 mature individuals |
| Gymnogyps californianus | 1987 [33] | 22 animals, 1981 [33] | 85 animals, 1982–2003 [33] | 1992 [34] | Sespe Condor Sanctuary, Los Padres [34] | 93 mature individuals |
| Canis rufus | 1980 [35] | 14 animals, 1976 [35] | 102 animals, 1987–2002 [35] | 1987 [35] | Alligator River National Wildlife Refuge, Carolina [35] | 20–30 mature individuals |
| Mustela nigripes | 1980s [36] | 18 animals, 1985–87 [36] | 37 animals, 1991 [37] | From 1991 [36] | Shirley Basin, Wyoming [36] | 206 mature individuals |
| Dama mesopotamica | - | 25 animals, 1956 [38] | Iran: 6 animals, 1964–1965 [39] Israel: 7 animals, 1976 [38] | From 1977 [39] 1996 [38] | Different sites in Iran [39] Nahal Kziv Nature Reserve [38,40] | >250 adults |
| Elaphurus davidianus | >1500 years ago [41] | 11 animals, 1900–1910 [41] | 77 animals, 1985–1987 [41] | 1987 [41] | Dafeng Milu Nature Reserve [41] | ~600 individuals |
| Oryx leucoryx | 1972 [42] | 14 animals, 1962–67 (“World Herd”) [43] +few animals from private collections [44] | Oman: 40 animals, 1980–1995 [45] | 1982 [45] | Arabian Oryx Sanctuary [45] | 850 mature individuals |
| Jordan: 14 animals, 1978–84 [46] | 1983 [46] | Shaumari Nature Reserve [46] | ||||
| Israel: 7 animals, 1978 [44] | 1997–1998 [44] | Shahak Spring [44] | ||||
| Saudi Arabia: 35 animals, 1989 [47] | 1990–1993 [48] 1995–2003 [48] | Mahazat as-Sayd [48] Uruq Bani Ma’arid [48] | ||||
| UAE: 90 animals [49] | 2007 [49] | Um El Zumool [49] | ||||
| Bison bonasus | 1927 [50] | 54 animals, 1920s [51] | 18 animals [50] | 1952–1966 [50] | Puszcza Bialowieska [50] | 2518 mature individuals |
| Equus ferus przewalskii | 1960s [52] | 53 animals, 1920s [53] | 16 animals, 1900–1947 [53,54] | 1992–2004 [53] 1992–2000 [53] | Great Gobi B via Takhin Tal [54] Hustai National Park [53] | 178 mature individuals |
| Ceratotherium s. simum | - | 20–50 animals [55] ~200 animals [56], 1900 | South Africa: 330 animals [57] | 1961–67 [57] | Different reserves in South Africa [57] | 10,080 mature individuals |
3. Comparing Ecosystems via Longitudinal Studies—Even beyond the Borders of Biogeographical Regions
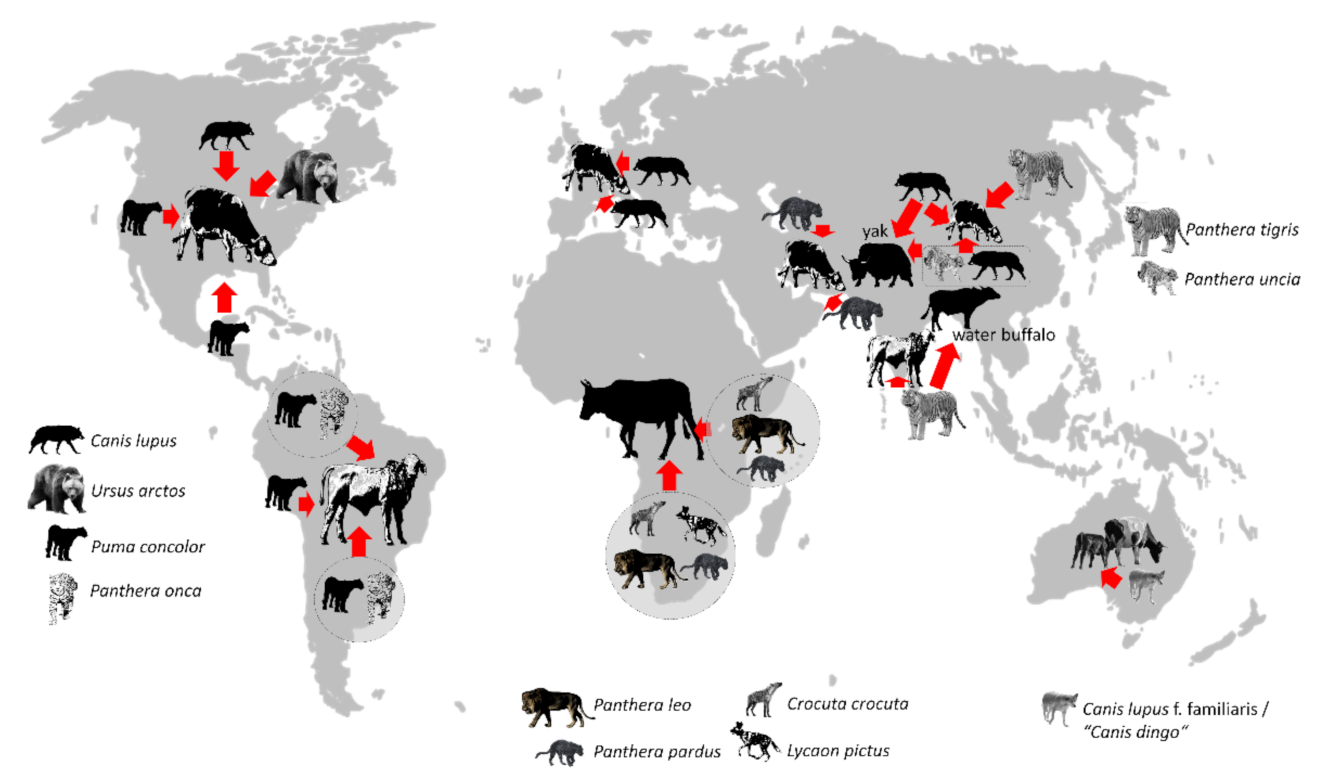
4. Carnivore–Livestock Conflicts—A Classic Example for Transregional Considerations

5. A Novel and Rapidly Emerging, Global Human–Wildlife Conflict Resulting from Wind Energy Development
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; de Vries, W.; de Wit, C.A.; et al. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Kastner, T. Land use biodiversity impacts embodied in international food trade. Glob. Environ. Chang. 2016, 38, 195–204. [Google Scholar] [CrossRef]
- Chaudhary, A.; Pfister, S.; Hellweg, S. Spatially explicit analysis of biodiversity loss due to global agriculture, pasture and forest land use from a producer and consumer perspective. Environ. Sci. Technol. 2016, 50, 3928–3936. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Martins, I.S.; Kastner, T.; Plutzar, C.; Theurl, M.C.; Eisenmenger, N.; Huijbregts, M.A.J.; Wood, R.; Stadler, K.; Bruckner, M.; et al. Increasing impacts of land use on biodiversity and carbon sequestration driven by population and economic growth. Nat. Ecol. Evol. 2019, 3, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Gadenne, D.L.; Kennedy, J.; McKeiver, C. An empirical study of environmental awareness and practices in SMEs. J. Bus. Ethics 2009, 84, 45–63. [Google Scholar] [CrossRef]
- Stoldt, M.; Göttert, T.; Mann, C.; Zeller, U. Transfrontier Conservation Areas and Human-Wildlife Conflict: The Case of the Namibian Component of the Kavango-Zambezi (KAZA) TFCA. Sci. Rep. 2020, 10, 7964. [Google Scholar] [CrossRef]
- Thornton, D.; Branch, L.; Murray, D. Distribution and connectivity of protected areas in the Americas facilitates transboundary conservation. Ecol. Appl. 2020, 30, e02027. [Google Scholar] [CrossRef]
- Distefano, E. Human-Wildlife Conflict Worldwide: Collection of Case Studies, Analysis of Management Strategies and Good Practices; Food and Agricultural Organization of the United Nations (FAO) Sustainable Agriculture; Rural Development Initiative (SARDI): Rome, Italy, 2005. [Google Scholar]
- Madden, F. Creating coexistence between humans and wildlife: Global perspectives on local efforts to address human-wildlife conflict. Hum. Dimens. Wildl. 2004, 9, 247–257. [Google Scholar] [CrossRef]
- Red List Index. Available online: https://www.iucnredlist.org/assessment/red-list-index (accessed on 20 April 2022).
- United Nations—World Population Prospects 2019. Available online: https://population.un.org/wpp/Download/Standard/Population/ (accessed on 20 April 2022).
- Conover, M.R. Resolving Human-Wildlife Conflicts: The Science of Wildlife Damage Management, 1st ed.; Lewis Publishers: Boca Raton, FL, USA, 2001; p. 440. [Google Scholar]
- Woodroffe, R.; Thirgood, S.; Rabinowitz, A. People and Wildlife, Conflict or Co-Existence? Cambridge University Press: New York, NY, USA, 2005. [Google Scholar]
- Nyhus, P.J. Human–wildlife conflict and coexistence. Annu. Rev. Environ. Resour. 2016, 41, 143–171. [Google Scholar] [CrossRef]
- Treves, A.; Karanth, K.U. Human-carnivore conflict and perspectives on carnivore management worldwide. Conserv. Biol. 2003, 17, 1491–1499. [Google Scholar] [CrossRef]
- Peterson, M.N.; Birckhead, J.L.; Leong, K.; Peterson, M.J.; Peterson, T.R. Rearticulating the myth of human–wildlife conflict. Conserv. Lett. 2010, 3, 74–82. [Google Scholar] [CrossRef]
- Fletcher, R.; Toncheva, S. The political economy of human-wildlife conflict and coexistence. Biol. Conserv. 2021, 260, 109216. [Google Scholar] [CrossRef]
- Dickman, A.J. Complexities of conflict: The importance of considering social factors for effectively resolving human–wildlife conflict. Anim. Conserv. 2010, 13, 458–466. [Google Scholar] [CrossRef]
- Redpath, S.M.; Bhatia, S.; Young, J. Tilting at wildlife: Reconsidering human–wildlife conflict. Oryx 2015, 49, 222–225. [Google Scholar] [CrossRef]
- Frank, B.; Glikman, J.A.; Marchini, S. Human–Wildlife Interactions: Turning Conflict into Coexistence; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar] [CrossRef]
- Hodgson, I.D.; Redpath, S.M.; Sandstrom, C.; Biggs, D. The State of Knowledge and Practice on Human-Wildlife Conflicts; Research Report; The Luc Hoffman Institute: Gland, Switzerland, 2020. [Google Scholar]
- Hempel, E.; Bibi, F.; Faith, J.T.; Brink, J.S.; Kalthoff, D.C.; Kamminga, P.; Paijmans, J.L.A.; Westbury, M.V.; Hofreiter, M.; Zachos, F.E. Identifying the true number of specimens of the extinct blue antelope (Hippotragus leucophaeus). Sci. Rep. 2021, 11, 2100. [Google Scholar] [CrossRef]
- Waterhouse, J.; Mitchell, C. Has Anybody Seen a Tasmanian Tiger Lately? Ethical and Ontological Considerations of Thylacine De-Extinction. Green Lett. 2021, 25, 1–14. [Google Scholar] [CrossRef]
- Cuker, B.E. Passenger pigeon and waterfowl: Flights to extinction and not. In Diet for a Sustainable Ecosystem, 1st ed.; Cuker, B.E., Ed.; Springer: Cham, Switzerland, 2020; pp. 155–174. [Google Scholar] [CrossRef]
- Turvey, S.T.; Crees, J.J. Extinction in the Anthropocene. Curr. Biol. 2019, 29, R982–R986. [Google Scholar] [CrossRef]
- Boast, A.P.; Weyrich, L.S.; Wood, J.R.; Metcalf, J.L.; Knight, R.; Cooper, A. Coprolites reveal ecological interactions lost with the extinction of New Zealand birds. Proc. Natl. Acad. Sci. USA 2018, 115, 1546–1551. [Google Scholar] [CrossRef]
- IUCN/SSC. Guidelines for Reintroductions and Other Conservation Translocations; Version 1.0; IUCN Species Survival Commission: Gland, Switzerland, 2013; p. 57. [Google Scholar]
- Kleiman, D.G.; Beck, B.B.; Dietz, J.M.; Dietz, L.A.; Ballou, J.D.; Coimbra-Filho, A.F. Conservation program for the golden lion tamarin: Captive research and management, ecological studies, educational strategies, and reintroduction. In Primates—The Road to Self-Sustaining Populations; Benirschke, K., Ed.; Springer: New York, NY, USA, 1986; pp. 959–979. [Google Scholar]
- Böer, M.; Brain, C.; Cantzler, T.; Hamza, H.; Venzke, K. Reintroduction of a captive born white rhinoceros (Ceratotherium simum simum) to the Etosha National Park. Part II: Behaviour in the boma, procedures before and locomotion during the first ten weeks after release and observations from June until September 1997. Zool. Gart. 1999, 69, 73–81. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 20 April 2022).
- BirdLife International. Strigops habroptila. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2018; p. e.T22685245A129751169. [Google Scholar] [CrossRef]
- BirdLife International. Leucopsar rothschildi. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2021; p. e.T22710912A183006359. [Google Scholar] [CrossRef]
- BirdLife International. Gymnogyps californianus. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2020; p. e.T22697636A181151405. [Google Scholar] [CrossRef]
- Cohn, J.P. Saving the California condor. Bioscience 1999, 49, 864–868. [Google Scholar] [CrossRef]
- Phillips, M. Canis rufus (Errata Version Published in 2020). The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2018; p. e.T3747A163509841. [Google Scholar] [CrossRef]
- Belant, J.; Biggins, D.; Garelle, D.; Griebel, R.G.; Hughes, J.P. Mustela nigripes. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2015; p. e.T14020A45200314. [Google Scholar] [CrossRef]
- Biggins, D.E.; Miller, B.J.; Hanebury, L.R.; Powell, R.A. Mortality of Siberian polecats and black-footed ferrets released onto prairie dog colonies. J. Mammal. 2011, 92, 721–731. [Google Scholar] [CrossRef]
- Bar-David, S.; Saltz, D.; Dayan, T.; Shkedy, Y. Using spatially expanding populations as a tool for evaluating landscape planning: The reintroduced Persian fallow deer as a case study. J. Nat. Conserv. 2008, 16, 164–174. [Google Scholar] [CrossRef]
- Werner, N.Y.; Rabiei, A.; Saltz, D.; Daujat, J.; Baker, K. Dama mesopotamica. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2015; p. e.T6232A97672550. [Google Scholar] [CrossRef]
- Saltz, D. A long-term systematic approach to planning reintroductions: The Persian fallow deer and the Arabian oryx in Israel. Anim. Conserv. Forum 1998, 1, 245–252. [Google Scholar] [CrossRef]
- Jiang, Z.; Harris, R.B. Elaphurus davidianus. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2016; p. e.T7121A22159785. [Google Scholar] [CrossRef]
- Henderson, D.S. Were they the last Arabian oryx? Oryx 1974, 12, 347–350. [Google Scholar] [CrossRef]
- Marshall, T.C.; Spalton, J.A. Simultaneous inbreeding and outbreeding depression in reintroduced Arabian oryx. Anim. Conserv. Forum 2000, 3, 241–248. [Google Scholar] [CrossRef]
- Gilad, O.; Grant, W.E.; Saltz, D. Simulated dynamics of Arabian Oryx (Oryx leucoryx) in the Israeli Negev: Effects of migration corridors and post-reintroduction changes in natality on population viability. Ecol. Model. 2008, 210, 169–178. [Google Scholar] [CrossRef]
- Spalton, J.A.; Lawerence, M.W.; Brend, S.A. Arabian oryx reintroduction in Oman: Successes and setbacks. Oryx 1999, 33, 168–175. [Google Scholar] [CrossRef]
- Harding, L.E.; Abu-Eid, O.F.; Hamidan, N.; al Sha’lan, A. Reintroduction of the Arabian oryx Oryx leucoryx in Jordan: War and redemption. Oryx 2007, 41, 478–487. [Google Scholar] [CrossRef]
- Vassart, M.; Granjon, L.; Greth, A. Genetic variability in the Arabian oryx (Oryx leucoryx). Zoo Biol. 1991, 10, 399–408. [Google Scholar] [CrossRef]
- Mésochina, P.; Bedin, E.; Ostrowski, S. Reintroducing antelopes into arid areas: Lessons learnt from the oryx in Saudi Arabia. Comptes Rendus Biol. 2003, 326, 158–165. [Google Scholar] [CrossRef]
- El Alqamy, H.; Senn, H.; Roberts, M.F.; McEwing, R.; Ogden, R. Genetic assessment of the Arabian oryx founder population in the Emirate of Abu Dhabi, UAE: An example of evaluating unmanaged captive stocks for reintroduction. Conserv. Genet. 2012, 13, 79–88. [Google Scholar] [CrossRef]
- Pucek, Z.; Belousova, I.; Krasińska, M.; Krasiński, Z.; Olech, W. Status survey and conservation action plan. European bison. In IUCN/SSC Bison Specialist Group; Pucek, Z., Ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2004; p. 54. [Google Scholar]
- Olech, W. Bison bonasus. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2008; p. e.T2814A9484719. [Google Scholar] [CrossRef]
- Slotta-Bachmayr, L.; Boegel, R.; Kaczensky, P.; Stauffer, C.; Walzer, C. Use of population viability analysis to identify management priorities and success in reintroducing Przewalski’s horses to southwestern Mongolia. J. Wildl. Manag. 2004, 68, 790–798. [Google Scholar] [CrossRef]
- King, S.R.B.; Boyd, L.; Zimmermann, W.; Kendall, B.E. Equus ferus. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2015; p. e.T41763A97204950. [Google Scholar] [CrossRef]
- Bowling, A.T.; Ryder, O.A. Genetic studies of blood markers in Przewalski’s horses. J. Hered. 1987, 78, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Emslie, R. Ceratotherium simum. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2012; p. e.T4185A16980466. [Google Scholar]
- Rookmaaker, K. The alleged population reduction of the southern white rhinoceros (Ceratotherium simum simum) and the successful recovery. Säugetierkd. Mitt. 2000, 45, 55–70. [Google Scholar]
- Player, I. Translocation of white rhinoceros in South Africa. Oryx 1967, 9, 137–150. [Google Scholar] [CrossRef]
- Wallace, A.R. The Geographical Distribution of Animals with a Study of the Relations of Living and Extinct Faunas as Elucidating the Past Changes of the Earth’s Surface; Macmillan and Co.: London, UK, 1876; Volume II, p. 607. [Google Scholar]
- de Silva, S. Need for longitudinal studies of Asian wildlife in the face of crises. Glob. Ecol. Conserv. 2016, 6, 276–285. [Google Scholar] [CrossRef]
- Zeller, U.; Starik, N.; Göttert, T. Biodiversity, land use and ecosystem services—An organismic and comparative approach to different geographical regions. Glob. Ecol. Conserv. 2017, 10, 114–125. [Google Scholar] [CrossRef]
- Rottstock, T.; Göttert, T.; Zeller, U. Relatively undisturbed African savannas-an important reference for assessing wildlife responses to livestock grazing systems in European rangelands. Glob. Ecol. Conserv. 2020, 23, e01124. [Google Scholar] [CrossRef]
- Zeller, U.; Göttert, T. Humans, megafauna and landscape structure–Rock engravings from Namibia encourage a comparative approach to central Europe and southern Africa. Vertebr. Zool. 2021, 71, 631. [Google Scholar] [CrossRef]
- Meulenkamp, J.E.; Sissingh, W. Tertiary palaeogeography and tectonostratigraphic evolution of the Northern and Southern Peri-Tethys platforms and the intermediate domains of the African–Eurasian convergent plate boundary zone. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 196, 209–228. [Google Scholar] [CrossRef]
- Janis, C.M. Tertiary mammal evolution in the context of changing climates, vegetation, and tectonic events. Annu. Rev. Ecol. Evol. Syst. 1993, 24, 467–500. [Google Scholar] [CrossRef]
- Kahlke, H.D. Biostratigraphical correlations (mammals) of the Quaternary continental deposits of Europe and the Far East. Quartärpaläontologie 1986, 6, 83–86. [Google Scholar]
- Newton, I. The Migration Ecology of Birds, 1st ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Eisenberg, J.F. An introduction to the Carnivora. In Carnivore Behavior, Ecology, and Evolution; Gittleman, J.L., Ed.; Springer: Boston, MA, USA, 1989; pp. 1–9. [Google Scholar] [CrossRef]
- Gervasi, V.; Nilsen, E.B.; Linnell, J.D. Body mass relationships affect the age structure of predation across carnivore–ungulate systems: A review and synthesis. Mammal Rev. 2015, 45, 253–266. [Google Scholar] [CrossRef]
- Prugh, L.R.; Sivy, K.J.; Mahoney, P.J.; Ganz, T.R.; Ditmer, M.A.; van de Kerk, M.; Gilbert, S.L.; Montgomery, R.A. Designing studies of predation risk for improved inference in carnivore-ungulate systems. Biol. Conserv. 2019, 232, 194–207. [Google Scholar] [CrossRef]
- Bohlken, H. Haustiere und zoologische Systematik. Z. Tierzuechtung Zuechtungsbiol. 1961, 76, 107–113. [Google Scholar] [CrossRef]
- Zeller, U.; Göttert, T. The relations between evolution and domestication reconsidered-implications for systematics, ecology, and nature conservation. Glob. Ecol. Conserv. 2019, 20, e00756. [Google Scholar] [CrossRef]
- Ambarlı, H.; Bilgin, C.C. Human–brown bear conflicts in Artvin, northeastern Turkey: Encounters, damage, and attitudes. Ursus 2008, 19, 146–153. [Google Scholar] [CrossRef]
- Zarzo-Arias, A.; Delgado, M.M.; Palazón, S.; Afonso Jordana, I.; Bombieri, G.; González-Bernardo, E.; Bettega, O.C.; García-González, R.; Penteriani, V. Seasonality, local resources and environmental factors influence patterns of brown bear damages: Implications for management. J. Zool. 2021, 313, 1–17. [Google Scholar] [CrossRef]
- Fechter, D.; Storch, I. How many wolves (Canis lupus) fit into Germany? The role of assumptions in predictive rule-based habitat models for habitat generalists. PLoS ONE 2014, 9, e101798. [Google Scholar] [CrossRef]
- Musiani, M.; Muhly, T.; Gates, C.C.; Callaghan, C.; Smith, M.E.; Tosoni, E. Seasonality and reoccurrence of depredation and wolf control in western North America. Wildl. Soc. Bull. 2005, 33, 876–887. [Google Scholar] [CrossRef]
- Wielgus, R.B.; Peebles, K.A. Effects of wolf mortality on livestock depredations. PLoS ONE 2014, 9, e113505. [Google Scholar] [CrossRef] [PubMed]
- Wells, S.L.; McNew, L.B.; Tyers, D.B.; Van Manen, F.T.; Thompson, D.J. Grizzly bear depredation on grazing allotments in the Yellowstone Ecosystem. J. Wildl. Manag. 2019, 83, 556–566. [Google Scholar] [CrossRef]
- Guerisoli, M.D.L.M.; Luengos Vidal, E.; Caruso, N.; Giordano, A.J.; Lucherini, M. Puma–livestock conflicts in the Americas: A review of the evidence. Mammal Rev. 2020, 51, 228–246. [Google Scholar] [CrossRef]
- Zarco-González, M.M.; Monroy-Vilchis, O.; Rodríguez-Soto, C.; Urios, V. Spatial factors and management associated with livestock predations by Puma concolor in Central Mexico. Hum. Ecol. 2012, 40, 631–638. [Google Scholar] [CrossRef]
- Polisar, J.; Maxit, I.; Scognamillo, D.; Farrell, L.; Sunquist, M.E.; Eisenberg, J.F. Jaguars, pumas, their prey base, and cattle ranching: Ecological interpretations of a management problem. Biol. Conserv. 2003, 109, 297–310. [Google Scholar] [CrossRef]
- Michalski, F.; Boulhosa, R.L.P.; Faria, A.; Peres, C.A. Human–wildlife conflicts in a fragmented Amazonian forest landscape: Determinants of large felid depredation on livestock. Anim. Conserv. 2006, 9, 179–188. [Google Scholar] [CrossRef]
- Palmeira, F.B.; Crawshaw Jr, P.G.; Haddad, C.M.; Ferraz, K.M.P.; Verdade, L.M. Cattle depredation by puma (Puma concolor) and jaguar (Panthera onca) in central-western Brazil. Biol. Conserv. 2008, 141, 118–125. [Google Scholar] [CrossRef]
- Kolowski, J.M.; Holekamp, K.E. Spatial, temporal, and physical characteristics of livestock depredations by large carnivores along a Kenyan reserve border. Biol. Conserv. 2006, 128, 529–541. [Google Scholar] [CrossRef]
- Ogada, M.O.; Woodroffe, R.; Oguge, N.O.; Frank, L.G. Limiting depredation by African carnivores: The role of livestock husbandry. Conserv. Biol. 2003, 17, 1521–1530. [Google Scholar] [CrossRef]
- LeFlore, E.G.; Fuller, T.K.; Tomeletso, M.; Stein, A.B. Livestock depredation by large carnivores in northern Botswana. Glob. Ecol. Conserv. 2019, 18, e00592. [Google Scholar] [CrossRef]
- Magrini, C. First data on canids depredation on livestock in an area of recent recolonization by wolf in Central Italy: Considerations on conflict survey and prevention methods. Ekológia 2014, 33, 81. [Google Scholar] [CrossRef][Green Version]
- Gula, R. Wolf depredation on domestic animals in the Polish Carpathian Mountains. J. Wildl. Manag. 2008, 72, 283–289. [Google Scholar] [CrossRef]
- Sidorovich, V.E.; Tikhomirova, L.L.; Jędrzejewska, B. Wolf Canis lupus numbers, diet and damage to livestock in relation to hunting and ungulate abundance in northeastern Belarus during 1990–2000. Wildl. Biol. 2003, 9, 103–111. [Google Scholar] [CrossRef]
- Nakazawa, C.; Tungalagtuya, K.; Maruyama, N.; Suda, K. Food habits of gray wolves in the Bogdkhan mountain strictly protected area, Mongolia. Biosph. Conserv. Nat. Wildl. Hum. 2008, 9, 1–8. [Google Scholar] [CrossRef]
- Kirilyuk, A.; Ke, R. Wolf depredation on livestock in Daursky State Nature Biosphere Reserve, Russia. J. Nat. Conserv. 2020, 58, 125916. [Google Scholar] [CrossRef]
- Khorozyan, I.; Ghoddousi, S.; Soufi, M.; Soofi, M.; Waltert, M. Studded leather collars are very effective in protecting cattle from leopard (Panthera pardus) attacks. Ecol. Solut. Evid. 2020, 1, e12013. [Google Scholar] [CrossRef]
- Akrim, F.; Mahmood, T.; Belant, J.L.; Nadeem, M.S.; Qasim, S.; Asadi, M.A. Livestock depredations by leopards in Pir Lasura National Park, Pakistan: Characteristics, control and costs. Wildl. Biol. 2021, 2021, 00782. [Google Scholar] [CrossRef]
- Chetri, M.; Odden, M.; Devineau, O.; Wegge, P. Patterns of livestock depredation by snow leopards and other large carnivores in the Central Himalayas, Nepal. Glob. Ecol. Conserv. 2019, 17, e00536. [Google Scholar] [CrossRef]
- Suryawanshi, K.R.; Bhatnagar, Y.V.; Redpath, S.; Mishra, C. People, predators and perceptions: Patterns of livestock depredation by snow leopards and wolves. J. Appl. Ecol. 2013, 50, 550–560. [Google Scholar] [CrossRef]
- Kumar, S.; Khan, J.A.; Khan, A.; Musavi, A.; Malik, P.K.; Kushwaha, S.P.S.; Khati, D.S.; Sarin, G.D. An ecological study in the buffer zone of the Corbett Tiger Reserve: Tiger abundance and cattle depredation. Int. J. Ecol. Environ. Sci. 2008, 34, 133–140. [Google Scholar]
- Rahman, H.A.; Barlow, A.C.D.; Greenwood, C.J.; Islam, M.A.; Ahmad, I.U. Livestock Depredation by Tiger on the Edge of the Bangladesh Sundarbans: A Technical Report; Wildlife Trust of Bangladesh: Dhaka, Bangladesh, 2009. [Google Scholar]
- Soh, Y.H.; Carrasco, L.R.; Miquelle, D.G.; Jiang, J.; Yang, J.; Stokes, E.J.; Tang, J.; Kang, A.; Liu, P.; Rao, M. Spatial correlates of livestock depredation by Amur tigers in Hunchun, China: Relevance of prey density and implications for protected area management. Biol. Conserv. 2014, 169, 117–127. [Google Scholar] [CrossRef]
- Bing, L.I.; Endi, Z.H.A.N.G.; Zhongbao, L.I.U. Livestock depredation by Amur tigers in Hunchun Nature Reserve, Jilin, China. Acta Theriol. Sin. 2009, 29, 231. [Google Scholar]
- Campbell, G.; Coffey, A.; Miller, H.; Read, J.L.; Brook, A.; Fleming, P.J.S.; Bird, P.; Eldridge, S.; Allen, B.L. Dingo baiting did not reduce fetal/calf loss in beef cattle in northern South Australia. Anim. Prod. Sci. 2019, 59, 319–330. [Google Scholar] [CrossRef]
- Salafsky, N.; Salzer, D.; Stattersfield, A.J.; Hilton-Taylor, C.; Neugarten, R.; Butchart, S.H.; Collen, B.; Cox, N.; Master, L.L.; O’Connor, S.; et al. A standard lexicon for biodiversity conservation: Unified classifications of threats and actions. Conserv. Biol. 2008, 22, 897–911. [Google Scholar] [CrossRef]
- Saidur, R.; Rahim, N.A.; Islam, M.R.; Solangi, K.H. Environmental impact of wind energy. Renew. Sustain. Energy Rev. 2011, 15, 2423–2430. [Google Scholar] [CrossRef]
- Jaber, S. Environmental impacts of wind energy. J. Clean Energy Technol. 2013, 1, 251–254. [Google Scholar] [CrossRef]
- Schuster, E.; Bulling, L.; Köppel, J. Consolidating the state of knowledge: A synoptical review of wind energy’s wildlife effects. Environ. Manag. 2015, 56, 300–331. [Google Scholar] [CrossRef]
- Allison, T.D.; Diffendorfer, J.E.; Baerwald, E.F.; Beston, J.A.; Drake, D.; Hale, A.M.; Hein, C.D.; Huso, M.M.; Loss, S.R.; Lovich, J.E.; et al. Impacts to wildlife of wind energy siting and operation in the United States. Issues Ecol. 2019, 21, 2–18. [Google Scholar]
- Hall, L.S.; Richards, G.C. Notes on Tadarida australis (Chiroptera: Molossidae). Aust. Mammal. 1972, 1, 2. [Google Scholar]
- Johnson, G.D. A review of bat mortality at wind-energy developments in the United States. Bat Res. News 2005, 46, 45–49. [Google Scholar]
- Arnett, E.B.; Brown, W.K.; Erickson, W.P.; Fiedler, J.K.; Hamilton, B.L.; Henry, T.H.; Jain, A.; Johnson, G.D.; Kerns, J.; Koford, R.R.; et al. Patterns of bat fatalities at wind energy facilities in North America. J. Wildl. Manag. 2008, 72, 61–78. [Google Scholar] [CrossRef]
- Rydell, J.; Engström, H.; Hedenström, A.; Kyed Larsen, J.; Pettersson, J.; Green, M. The Effect of Wind Power on Birds and Bats: A Synthesis; Naturvårdsverket: Stockholm, Sweden, 2012. [Google Scholar]
- Voigt, C.C.; Lehnert, L.S.; Petersons, G.; Adorf, F.; Bach, L. Wildlife and renewable energy: German politics cross migratory bats. Eur. J. Wildl. Res. 2015, 61, 213–219. [Google Scholar] [CrossRef]
- Thaker, M.; Zambre, A.; Bhosale, H. Wind farms have cascading impacts on ecosystems across trophic levels. Nat. Ecol. Evol. 2018, 2, 1854–1858. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.; Rahmel, U. Summary of wind turbine impacts on bats—Assessment of a conflict. Bremer Beiträge Nat. Nat. 2004, 7, 245–252. [Google Scholar]
- Arnett, E.B.; Baerwald, E.F.; Mathews, F.; Rodrigues, L.; Rodríguez-Durán, A.; Rydell, J.; Villegas-Patraca, R.; Voigt, C.C. Impacts of wind energy development on bats: A global perspective. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Cham, Switzerland, 2016; pp. 295–323. [Google Scholar]
- O’Shea, T.J.; Cryan, P.M.; Hayman, D.T.; Plowright, R.K.; Streicker, D.G. Multiple mortality events in bats: A global review. Mammal Rev. 2016, 46, 175–190. [Google Scholar] [CrossRef]
- Voigt, C.C.; Straka, T.M.; Fritze, M. Producing wind energy at the cost of biodiversity: A stakeholder view on a green-green dilemma. J. Renew. Sustain. Energy 2019, 11, 063303. [Google Scholar] [CrossRef]
- Straka, T.M.; Fritze, M.; Voigt, C.C. The human dimensions of a green–green-dilemma: Lessons learned from the wind energy—Wildlife conflict in Germany. Energy Rep. 2020, 6, 1768–1777. [Google Scholar] [CrossRef]
- MacEwan, K.L.; Morgan, T.W.; Lötter, C.A.; Tredennick, A.T. Bat activity across South Africa: Implications for wind energy development. Afr. J. Wildl. Res. 2000, 50. [Google Scholar] [CrossRef]
- Dürr, T. Fledermäuse als Opfer von Windkraftanlagen in Deutschland. Nyctalus 2002, 8, 115–118. [Google Scholar]
- Dürr, T.; Bach, L. Bat deaths and wind turbines—A review of current knowledge, and of the information available in the database for Germany. Bremer Beiträge Nat. Nat. 2004, 7, 253–264. [Google Scholar]
- Rydell, J.; Bach, L.; Dubourg-Savage, M.J.; Green, M.; Rodrigues, L.; Hedenström, A. Mortality of bats at wind turbines links to nocturnal insect migration? Eur. J. Wildl. Res. 2010, 56, 823–827. [Google Scholar] [CrossRef]
- Rydell, J.; Bach, L.; Dubourg-Savage, M.J.; Green, M.; Rodrigues, L.; Hedenström, A. Bat mortality at wind turbines in northwestern Europe. Acta Chiropterologica 2010, 12, 261–274. [Google Scholar] [CrossRef]
- Grodsky, S.M.; Behr, M.J.; Gendler, A.; Drake, D.; Dieterle, B.D.; Rudd, R.J.; Walrath, N.L. Investigating the causes of death for wind turbine-associated bat fatalities. J. Mammal. 2011, 92, 917–925. [Google Scholar] [CrossRef]
- Baerwald, E.F.; D’Amours, G.H.; Klug, B.J.; Barclay, R.M. Barotrauma is a significant cause of bat fatalities at wind turbines. Curr. Biol. 2008, 18, R695–R696. [Google Scholar] [CrossRef]
- Ingersoll, T.E.; Sewall, B.J.; Amelon, S.K. Improved analysis of long-term monitoring data demonstrates marked regional declines of bat populations in the eastern United States. PLoS ONE 2013, 8, e65907. [Google Scholar] [CrossRef]
- Zahn, A.; Lustig, A.; Hammer, M. Potenzielle Auswirkungen von Windenergieanlagen auf Fledermauspopulationen. Anliegen Nat. 2014, 36, 21–35. [Google Scholar]
- Frick, W.F.; Baerwald, E.F.; Pollock, J.F.; Barclay, R.M.; Szymanski, J.A.; Weller, T.J.; Russell, A.L.; Loeb, S.C.; Medellin, R.A.; McGuire, L.P. Fatalities at wind turbines may threaten population viability of a migratory bat. Biol. Conserv. 2017, 209, 172–177. [Google Scholar] [CrossRef]
- Brinkmann, R.; Schauer-Weisshahn, H.; Bontadina, F. Survey of Possible Operational Impacts on Bats by Wind Facilities in Southern Germany; Report for Administrative District of Freiburg–Department; ECON Ecological Consultancy: Gundelfingen, Germany, 2006; p. 56. [Google Scholar]
- Frey-Ehrenbold, A.; Bontadina, F.; Arlettaz, R.; Obrist, M.K. Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. J. Appl. Ecol. 2013, 50, 252–261. [Google Scholar] [CrossRef]
- Claireau, F.; Bas, Y.; Pauwels, J.; Barré, K.; Machon, N.; Allegrini, B.; Puechmaille, S.J.; Kerbiriou, C. Major roads have important negative effects on insectivorous bat activity. Biol. Conserv. 2019, 235, 53–62. [Google Scholar] [CrossRef]
- Rodrigues, L.; Bach, L.; Dubourg-Savage, M.-J.; Karapandza, B.; Kovac, D.; Kervyn, T.; Dekker, J.; Kepel, A.; Bach, P.; Collins, J.; et al. Guidelines for Consideration of Bats in Wind Farm Projects—Revision 2014; EUROBATS Publication Serie; UNEP/EUROBATS: Bonn, Germany, 2015; p. 133. [Google Scholar]
- Barré, K.; Le Viol, I.; Bas, Y.; Julliard, R.; Kerbiriou, C. Estimating Habitat Loss Due to Wind Turbine Avoidance by Bats: Implications for European Siting Guidance. Biol. Conserv. 2018, 226, 205–214. [Google Scholar] [CrossRef]
- Ferri, V.; Battisti, C.; Soccini, C. Bats in a mediterranean mountainous landscape: Does wind farm repowering induce changes at assemblage and species level? Environ. Manag. 2016, 57, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Barclay, R.M.; Baerwald, E.F.; Gruver, J.C. Variation in bat and bird fatalities at wind energy facilities: Assessing the effects of rotor size and tower height. Can. J. Zool. 2007, 85, 381–387. [Google Scholar] [CrossRef]
- Cryan, P.M.; Barclay, R.M. Causes of bat fatalities at wind turbines: Hypotheses and predictions. J. Mammal. 2009, 90, 1330–1340. [Google Scholar] [CrossRef]
- Guest, E.E.; Stamps, B.F.; Durish, N.D.; Hale, A.M.; Hein, C.D.; Morton, B.P.; Weaver, S.P.; Fritts, S.R. An updated review of hypotheses regarding bat attraction to wind turbines. Animals 2022, 12, 343. [Google Scholar] [CrossRef] [PubMed]
- Kunz, T.H.; Arnett, E.B.; Erickson, W.P.; Hoar, A.R.; Johnson, G.D.; Larkin, R.P.; Strickland, M.D.; Thresher, R.W.; Tuttle, M.D. Ecological impacts of wind energy development on bats: Questions, research needs, and hypotheses. Front. Ecol. Environ. 2007, 5, 315–324. [Google Scholar] [CrossRef]
- Russ, J.M.; Montgomery, W.I. Habitat associations of bats in Northern Ireland: Implications for conservation. Biol. Conserv. 2002, 108, 49–58. [Google Scholar] [CrossRef]
- Kusch, J.; Schotte, F. Effects of fine-scale foraging habitat selection on bat community structure and diversity in a temperate low mountain range forest. Folia Zool. 2007, 56, 263. [Google Scholar]
- Buchholz, S.; Kelm, V.; Ghanem, S.J. Mono-specific forest plantations are valuable bat habitats: Implications for wind energy development. Eur. J. Wildl. Res. 2021, 67, 1–12. [Google Scholar] [CrossRef]
- Starik, N.; Göttert, T.; Heitlinger, E.; Zeller, U. Bat community responses to structural habitat complexity resulting from management practices within different land use types—A case study from north-eastern Germany. Acta Chiropterologica 2018, 20, 387–405. [Google Scholar] [CrossRef]
- Lehnert, L.S.; Kramer-Schadt, S.; Schönborn, S.; Lindecke, O.; Niermann, I.; Voigt, C.C. Wind farm facilities in Germany kill noctule bats from near and far. PLoS ONE 2014, 9, e103106. [Google Scholar] [CrossRef]
- Kuvlesky, W.P., Jr.; Brennan, L.A.; Morrison, M.L.; Boydston, K.K.; Ballard, B.M.; Bryant, F.C. Wind energy development and wildlife conservation: Challenges and opportunities. J. Wildl. Manag. 2007, 71, 2487–2498. [Google Scholar] [CrossRef]
- Rodrigues, L.; Bach, L.; Dubourg-Savage, M.-J.; Goodwin, J.; Harbusch, C. Guidelines for Consideration of Bats in Wind Farm Projects; EUROBATS Publication Series No. 3 (English Version); UNEP/EUROBATS Secretariat: Bonn, Germany, 2008; p. 51. [Google Scholar]
- Baerwald, E.F.; Edworthy, J.; Holder, M.; Barclay, R.M. A large-scale mitigation experiment to reduce bat fatalities at wind energy facilities. J. Wildl. Manag. 2009, 73, 1077–1081. [Google Scholar] [CrossRef]
- Arnett, E.B.; Huso, M.M.; Schirmacher, M.R.; Hayes, J.P. Altering turbine speed reduces bat mortality at wind-energy facilities. Front. Ecol. Environ. 2011, 9, 209–214. [Google Scholar] [CrossRef]
- Köppel, J.; Dahmen, M.; Helfrich, J.; Schuster, E.; Bulling, L. Cautious but committed: Moving toward adaptive planning and operation strategies for renewable energy‘s wildlife implications. Environ. Manag. 2014, 54, 744–755. [Google Scholar] [CrossRef]
- Richardson, S.M.; Lintott, P.R.; Hosken, D.J.; Economou, T.; Mathews, F. Peaks in bat activity at turbines and the implications for mitigating the impact of wind energy developments on bats. Sci. Rep. 2021, 11, 3636. [Google Scholar] [CrossRef]
- Voigt, C.C.; Russo, D.; Runkel, V.; Goerlitz, H.R. Limitations of acoustic monitoring at wind turbines to evaluate fatality risk of bats. Mammal Rev. 2021, 51, 559–570. [Google Scholar] [CrossRef]
- Romano, W.B.; Skalski, J.R.; Townsend, R.L.; Kinzie, K.W.; Coppinger, K.D.; Miller, M.F. Evaluation of an acoustic deterrent to reduce bat mortalities at an Illinois wind farm. Wildl. Soc. Bull. 2019, 43, 608–618. [Google Scholar] [CrossRef]
- Arnett, E.B.; Hein, C.D.; Schirmacher, M.R.; Huso, M.M.; Szewczak, J.M. Evaluating the effectiveness of an ultrasonic acoustic deterrent for reducing bat fatalities at wind turbines. PLoS ONE 2013, 8, e65794. [Google Scholar] [CrossRef]
- Fritze, M.; Lehnert, L.S.; Heim, O.; Lindecke, O.; Roeleke, M.; Voigt, C.C. Fledermäuse im Schatten der Windenergie: Deutschlands Expert*innen vermissen Transparenz und bundesweite Standards in den Genehmigungsverfahren. Nat. Landsch. 2019, 51, 20–27. [Google Scholar]
- Manfredo, M.J.; Bruskotter, J.T.; Teel, T.L.; Fulton, D.; Schwartz, S.H.; Arlinghaus, R.; Oishi, S.; Uskul, A.K.; Redford, K.; Kitayama, S.; et al. Why social values cannot be changed for the sake of conservation. Conserv. Biol. 2017, 31, 772–780. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Göttert, T.; Starik, N. Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity. Diversity 2022, 14, 380. https://doi.org/10.3390/d14050380
Göttert T, Starik N. Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity. Diversity. 2022; 14(5):380. https://doi.org/10.3390/d14050380
Chicago/Turabian StyleGöttert, Thomas, and Nicole Starik. 2022. "Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity" Diversity 14, no. 5: 380. https://doi.org/10.3390/d14050380
APA StyleGöttert, T., & Starik, N. (2022). Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity. Diversity, 14(5), 380. https://doi.org/10.3390/d14050380