
Breeding Ecology of Adélie Penguins in Mid Victoria Land, Ross Sea Antarctica

 , , and
, , and
Abstract
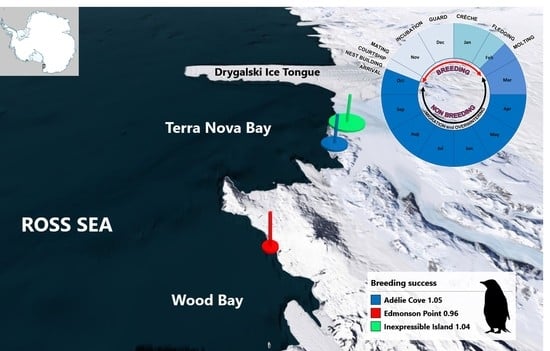
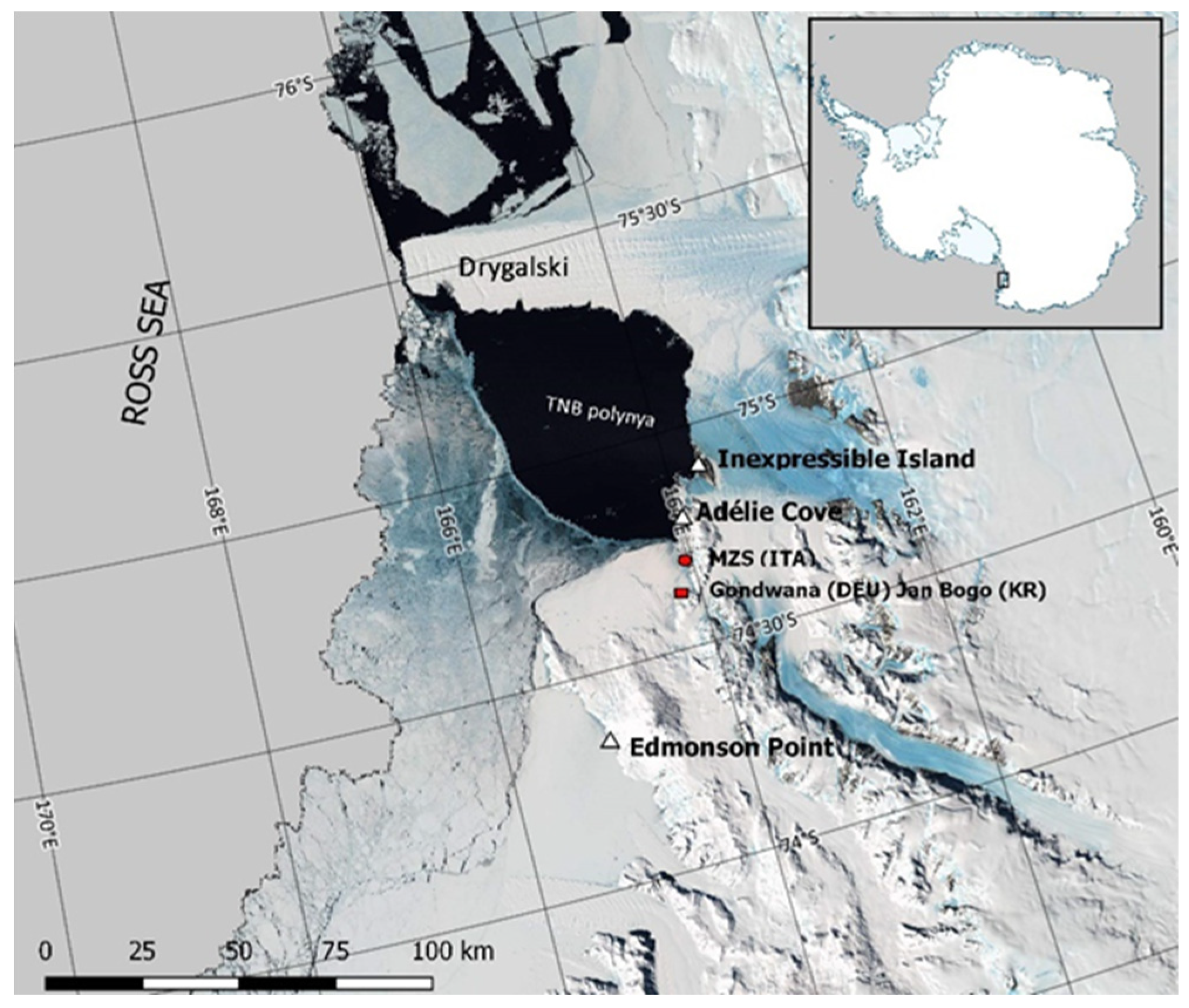
:
1. Introduction
2. Materials and Methods
2.1. Study Areas and Ecological Background
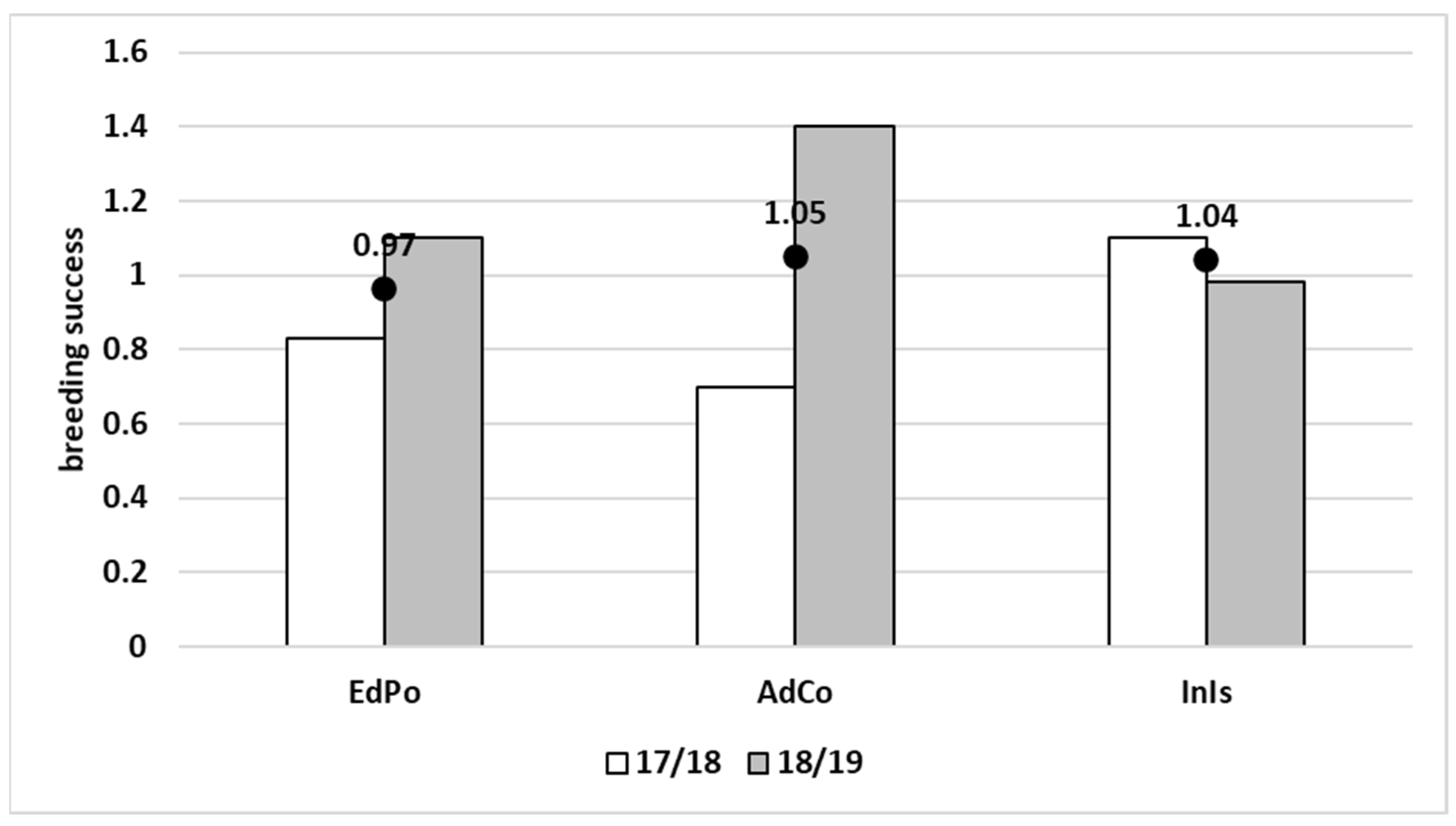
2.2. Naïve Estimation of Breeding Success (Three Colonies-Comparison)
2.3. Hatching and Breeding Success (Two Colonies-Comparison)
2.4. Chick Growth (Two Colonies-Comparison)
2.5. Predation Risk (Two Colonies-Comparison)
2.6. At Nest-Weather (Two Colonies-Comparison)
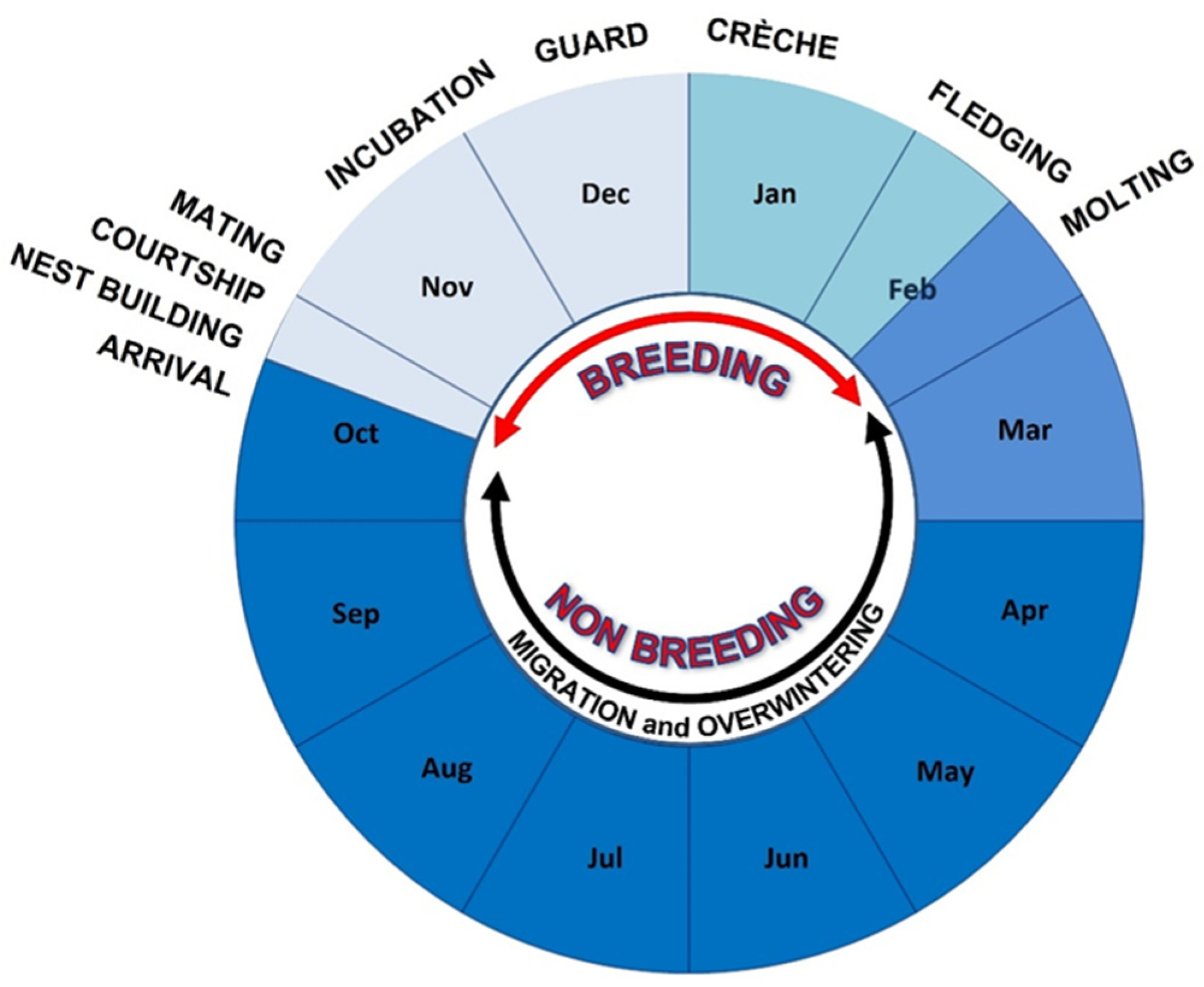
2.7. Breeding Chronology (Two Colonies-Comparison)
2.8. Statistical Analyses (Two Colonies-Comparison)
3. Results
3.1. Breeding Chronology in the Two Colonies
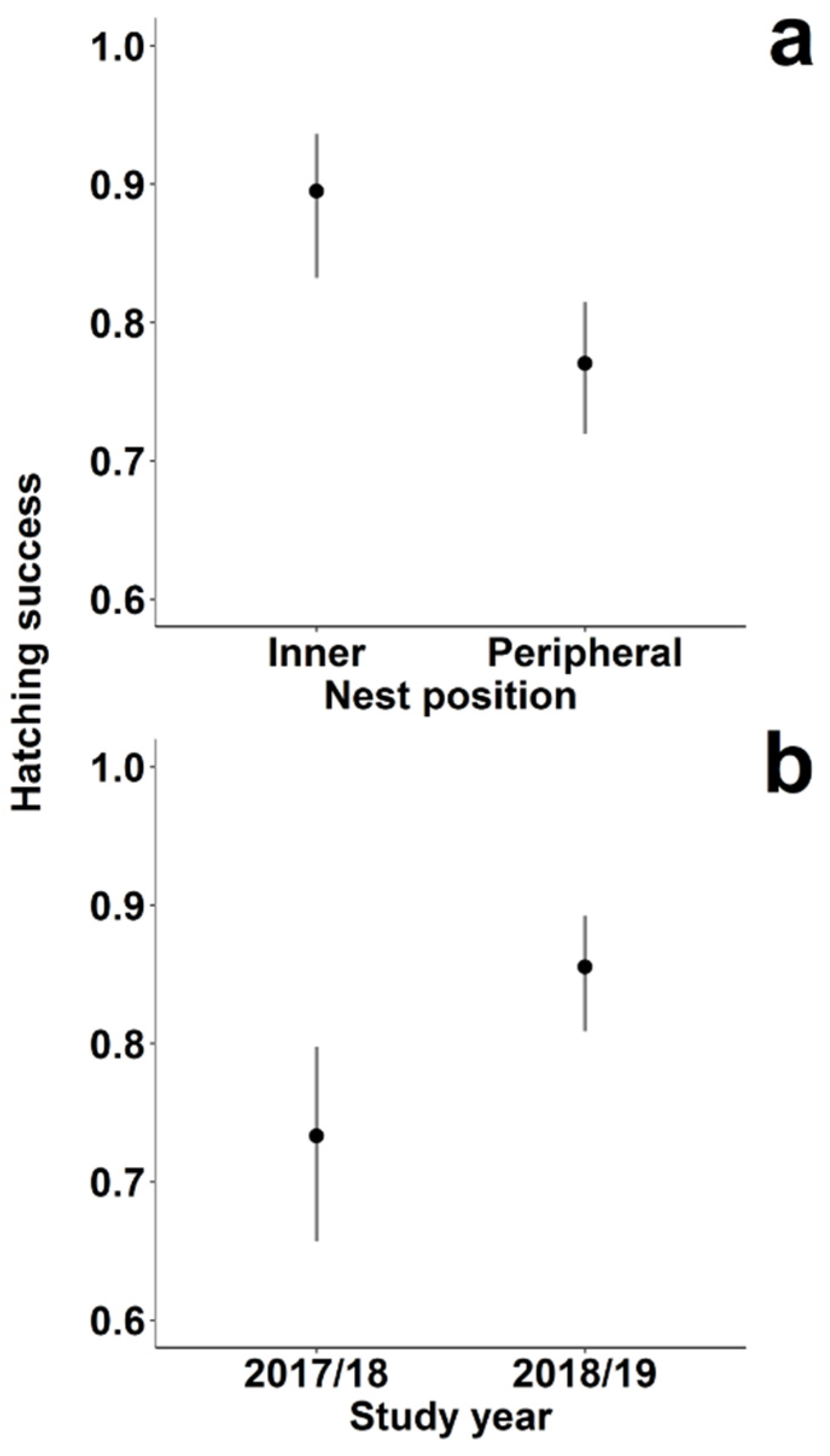
3.2. Factors Influencing Hatching and Breeding Success in the Two Colonies
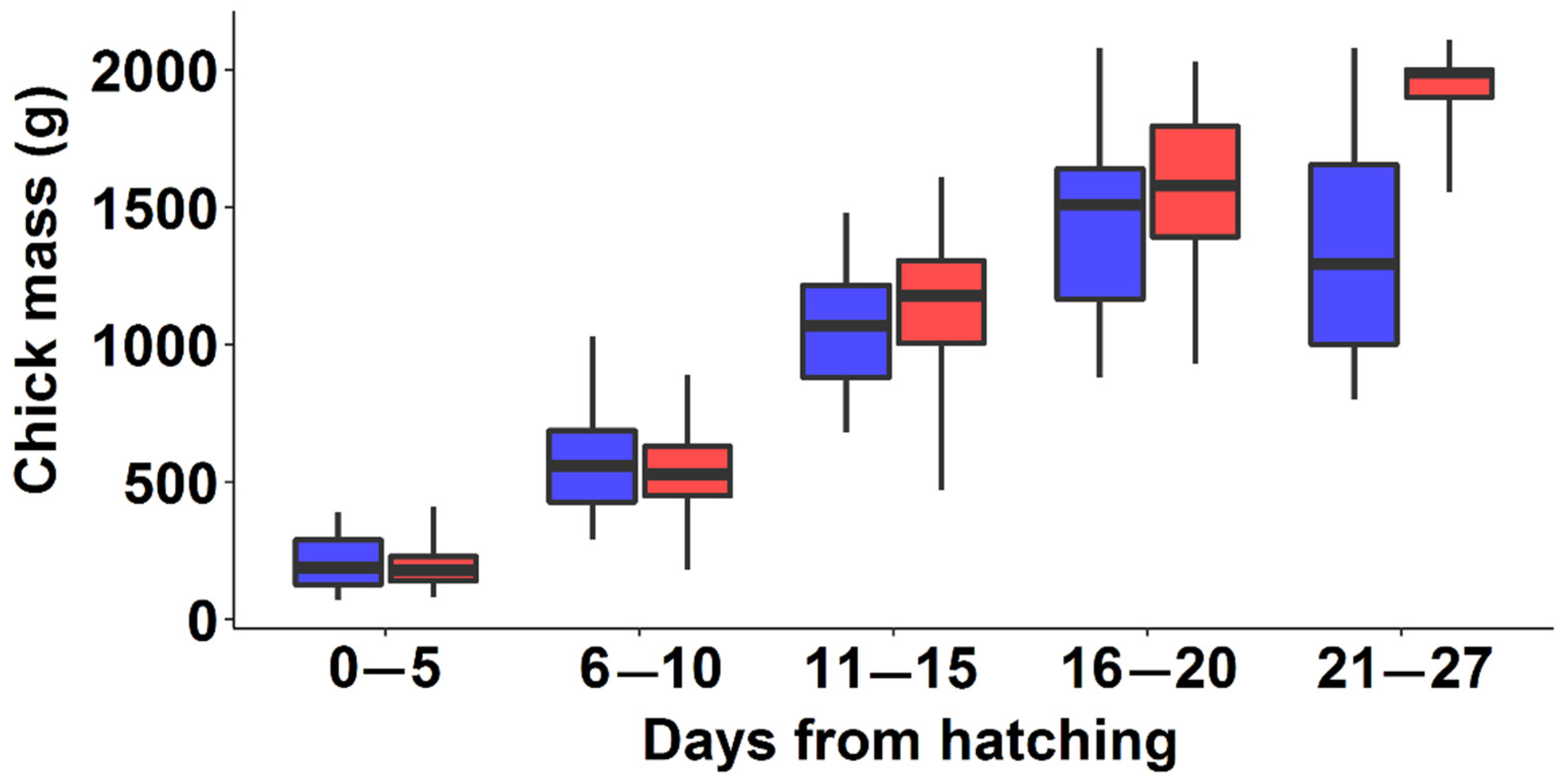
3.3. Differences in Chick Growth between the Two Colonies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montevecchi, W.A. Birds as Indicators of Change in Marine Prey Stocks. In Birds as Monitors of Environmental Change; Furness, R.W., Greenwood, J.J.D., Eds.; Chapman & Hall: London, UK, 1993; pp. 217–266. [Google Scholar]
- Frederiksen, M.; Edwards, M.; Richardson, A.J.; Halliday, N.C.; Wanless, S. From Plankton to Top Predators: Bottom-up Control of a Marine Food Web across Four Trophic Levels. J. Anim. Ecol. 2006, 75, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.; Mitchell, I.; Butler, A.; Ratcliffe, N.; Frederiksen, M.; Foster, S.; Reid, J.B. Seabirds as Indicators of the Marine Environment. ICES J. Mar. Sci. 2008, 65, 1520–1526. [Google Scholar] [CrossRef] [Green Version]
- Forcada, J.; Trathan, P.N. Penguin Responses to Climate Change in the Southern Ocean. Glob. Chang. Biol. 2009, 15, 1618–1630. [Google Scholar] [CrossRef]
- Oro, D. Seabirds and Climate: Knowledge, Pitfalls, and Opportunities. Front. Ecol. Evol. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Croxall, J.P.; McCann, T.S.; Prince, P.A.; Rothery, P. Reproductive Performance of Seabirds and Seals at South Georgia and Signy Island, South Orkney Islands, 1976–1987: Implications for Southern Ocean Monitoring Studies. In Antarctic Ocean and Resources Variability; Sahrhage, D., Ed.; Springer: Berlin, Germany, 1988; pp. 261–285. [Google Scholar]
- Schreiber, E.A.; Burger, J. Biology of Marine Birds, 1st ed.; CRC Press: Boca Raton, FL, USA, 2001; ISBN 978-0-429-12786-1. [Google Scholar]
- Votier, S.C.; Hatchwell, B.J.; Mears, M.; Birkhead, T.R. Changes in the Timing of Egg-Laying of a Colonial Seabird in Relation to Population Size and Environmental Conditions. Mar. Ecol. Prog. Ser. 2009, 393, 225–233. [Google Scholar] [CrossRef]
- Polito, M.J.; Trivelpiece, W.Z.; Patterson, W.P.; Karnovsky, N.J.; Reiss, C.S.; Emslie, S.D. Contrasting Specialist and Generalist Patterns Facilitate Foraging Niche Partitioning in Sympatric Populations of Pygoscelis Penguins. Mar. Ecol. Prog. Ser. 2015, 519, 221–237. [Google Scholar] [CrossRef]
- Trathan, P.N.; Wienecke, B.; Barbraud, C.; Jenouvrier, S.; Kooyman, G.; Le Bohec, C.; Ainley, D.G.; Ancel, A.; Zitterbart, D.P.; Chown, S.L.; et al. The Emperor Penguin—Vulnerable to Projected Rates of Warming and Sea Ice Loss. Biol. Conserv. 2020, 241, 108216. [Google Scholar] [CrossRef]
- Ronconi, R.A.; Lascelles, B.G.; Langham, G.M.; Reid, J.B.; Oro, D. The Role of Seabirds in Marine Protected Area Identification, Delineation, and Monitoring: Introduction and Synthesis. Seab. Mar. Prot. Areas Plan. 2012, 156, 1–4. [Google Scholar] [CrossRef]
- Hazen, E.L.; Abrahms, B.; Brodie, S.; Carroll, G.; Jacox, M.G.; Savoca, M.S.; Scales, K.L.; Sydeman, W.J.; Bograd, S.J. Marine Top Predators as Climate and Ecosystem Sentinels. Front. Ecol. Environ. 2019, 17, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Hindell, M.A.; Reisinger, R.R.; Ropert-Coudert, Y.; Hückstädt, L.A.; Trathan, P.N.; Bornemann, H.; Charrassin, J.-B.; Chown, S.L.; Costa, D.P.; Danis, B.; et al. Tracking of Marine Predators to Protect Southern Ocean Ecosystems. Nature 2020, 580, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Brooks, C.M.; Crowder, L.B.; Österblom, H.; Strong, A.L. Reaching Consensus for Conserving the Global Commons: The Case of the Ross Sea, Antarctica. Conserv. Lett. 2020, 13, e12676. [Google Scholar] [CrossRef]
- Brooks, C.M.; Bloom, E.; Kavanagh, A.; Nocito, E.S.; Watters, G.M.; Weller, J. The Ross Sea, Antarctica: A Highly Protected MPA in International Waters. Mar. Policy 2021, 134, 104795. [Google Scholar] [CrossRef]
- Dunn, A.; Vacchi, M.; Watters, G. The Ross Sea Region Marine Protected Area Research and Monitoring Plan; WG-EMM-17/43; CCAMLR: Hobart, Australia, 2017. [Google Scholar]
- CCAMLR Conservation Measure 91-05. Ross Sea Region Marine Protected Area 2016. Available online: https://cm.ccamlr.org/en/measure-91-05-2016 (accessed on 20 March 2022).
- Olmastroni, S.; Fattorini, N.; Ferretti, F.; Mori, E.; Burrini, L.; Simonetti, S.; Pezzo, F.; Ademollo, N.; Corsi, I. PenguinERA: Ecology, Reproduction and Adaptation for a Climate Change Sentinel. Italian PNRA Project for Monitoring Mid Victoria Land, Ross Sea, Adélie Penguin Population. In Proceedings of the Scar Open Science Conference, Online. 3–7 August 2020; Available online: https://www.scar.org/library/conferences/scar-open-science-conferences/abstracts/5534-scar-osc-2020-abstracts/ (accessed on 20 March 2022).
- CCAMLR. MPA Information Repository. Available online: https://cmir.ccamlr.org/node/67 (accessed on 15 March 2022).
- Youngflesh, C.; Jenouvrier, S.; Li, Y.; Ji, R.; Ainley, D.G.; Ballard, G.; Barbraud, C.; Delord, K.; Dugger, K.M.; Emmerson, L.M.; et al. Circumpolar Analysis of the Adélie Penguin Reveals the Importance of Environmental Variability in Phenological Mismatch. Ecology 2017, 98, 940–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimino, M.A.; Fraser, W.R.; Patterson-Fraser, D.L.; Saba, V.S.; Oliver, M.J. Large-Scale Climate and Local Weather Drive Interannual Variability in Adélie Penguin Chick Fledging Mass. Mar. Ecol. Prog. Ser. 2014, 513, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Emmerson, L.; Southwell, C. Sea Ice Cover and Its Influence on Adélie Penguin Reproductive Performance. Ecology 2008, 89, 2096–2102. [Google Scholar] [CrossRef]
- Dugger, K.M.; Ballard, G.; Ainley, D.G.; Lyver, P.O.; Schine, C. Adélie Penguins Coping with Environmental Change: Results from a Natural Experiment at the Edge of Their Breeding Range. Front. Ecol. Evol. 2014, 2, 68. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.-J.; Turney, C.S.M.; Fogwill, C.J.; Blair, E. The Impact of the Giant Iceberg B09B on Population Size and Breeding Success of Adélie Penguins in Commonwealth Bay, Antarctica. Antarct. Sci. 2016, 28, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Kentaro, I.; Nobuo, K.; Akinori, T. Foraging Behavior Links Sea Ice to Breeding Success in Antarctic Penguins. Sci. Adv. 2020, 6, eaba4828. [Google Scholar] [CrossRef]
- Wilson, P.R.; Ainley, D.G.; Nur, N.; Jacobs, S.S.; Barton, K.J.; Ballard, G.; Comiso, J.C. Adélie Penguin Population Change in the Pacific Sector of Antarctica: Relation to Sea-Ice Extent and the Antarctic Circumpolar Current. Mar. Ecol. Prog. Ser. 2001, 213, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Ballerini, T.; Tavecchia, G.; Olmastroni, S.; Pezzo, F.; Focardi, S. Nonlinear Effects of Winter Sea Ice on the Survival Probabilities of Adélie Penguins. Oecologia 2009, 161, 253–265. [Google Scholar] [CrossRef]
- Cimino, M.A.; Lynch, H.J.; Saba, V.S.; Oliver, M.J. Projected asymmetric response of Adélie penguins to Antarctic climate change. Sci. Rep. 2016, 6, 28785. [Google Scholar] [CrossRef] [PubMed]
- Lynch, H.J.; Naveen, R.; Trathan, P.N.; Fagan, W.F. Spatially integrated assessment reveals widespread changes in penguin populations on the Antarctic Peninsula. Ecology 2012, 93, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Barbraud, C.; Weimerskirch, H. Antarctic Birds Breed Later in Response to Climate Change. Proc. Natl. Acad. Sci. USA 2006, 103, 6248–6251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenouvrier, S.; Desprez, M.; Fay, R.; Barbraud, C.; Weimerskirch, H.; Delord, K.; Caswell, H. Climate Change and Functional Traits Affect Population Dynamics of a Long-Lived Seabird. J. Anim. Ecol. 2018, 87, 906–920. [Google Scholar] [CrossRef] [Green Version]
- Smiley, K.M.; Emmerson, L.M. A Matter of Timing: Adélie Penguin Reproductive Success in a Seasonally Varying Environment. Mar. Ecol. Prog. Ser. 2016, 542, 235–249. [Google Scholar] [CrossRef]
- Morandini, V.; Dugger, K.M.; Lescroël, A.; Schmidt, A.E.; Ballard, G. Maintenance of Nest Quality in Adélie Penguins Pygoscelis Adeliae: An Additional Benefit to Life in the Center. Polar Biol. 2021, 44, 1553–1562. [Google Scholar] [CrossRef]
- Schmidt, A.E.; Ballard, G.; Lescroël, A.; Dugger, K.M.; Jongsomjit, D.; Elrod, M.L.; Ainley, D.G. The Influence of Subcolony-Scale Nesting Habitat on the Reproductive Success of Adélie Penguins. Sci. Rep. 2021, 11, 15380. [Google Scholar] [CrossRef]
- Young, E. Skua and Penguin: Predator and Prey; Studies in Polar Research; Cambridge University Press: Cambridge, UK, 1994; ISBN 978-0-521-01813-5. [Google Scholar]
- Thierry, A.-M.; Ropert-Coudert, Y.; Raclot, T. Elevated Corticosterone Levels Decrease Reproductive Output of Chick-Rearing Adélie Penguins but Do Not Affect Chick Mass at Fledging. Conserv. Physiol. 2013, 1, cot007. [Google Scholar] [CrossRef]
- Whitehead, A.L.; Lyver, P.; Ballard, G.; Barton, K.; Karl, B.J.; Dugger, K.M.; Jennings, S.; Lescroel, A.; Wilson, P.R.; Ainley, D.G. Factors Driving Adélie Penguin Chick Size, Mass and Condition at Colonies of Different Sizes in the Southern Ross Sea. Mar. Ecol. Prog. Ser. 2015, 523, 199–213. [Google Scholar] [CrossRef]
- Wilson, D.J.; Lyver, P.O.; Greene, T.C.; Whitehead, A.L.; Dugger, K.M.; Karl, B.J.; Barringer, J.R.F.; McGarry, R.; Pollard, A.M.; Ainley, D.G. South Polar Skua Breeding Populations in the Ross Sea Assessed from Demonstrated Relationship with Adélie Penguin Numbers. Polar Biol. 2016, 40, 577–592. [Google Scholar] [CrossRef]
- Olmastroni, S.; Fattorini, N.; Pezzo, F.; Focardi, S. Gone Fishing: Adélie Penguin Site-Specific Foraging Tactics and Breeding Performance. Antarct. Sci. 2020, 32, 199–209. [Google Scholar] [CrossRef]
- Mori, E.; Brunetti, C.; Carapelli, A.; Burrini, L.; Fattorini, N.; Ferretti, F.; Olmastroni, S. Genetic Diversity in Clustered Colonies of an Antarctic Marine Mesopredator: A Role for Habitat Quality? Antarct. Sci. 2021, 33, 233–242. [Google Scholar] [CrossRef]
- Park, S.; Thiebot, J.-B.; Kim, J.-H.; Kim, K.W.; Chung, H.; Lee, W.Y. Mare Incognita: Adélie Penguins Foraging in Newly Exposed Habitat after Calving of the Nansen Ice Shelf. Environ. Res. 2021, 201, 111561. [Google Scholar] [CrossRef]
- NASA EOSDIS Worldview. Available online: https://worldview.earthdata.nasa.gov (accessed on 15 March 2022).
- Oceanites Inc. Explore MAPPPD. Available online: https://www.penguinmap.com/ (accessed on 15 March 2022).
- Ainley, D.G. The Adélie Penguin: Bellwether of Climate Change; Columbia University Press: New York, NY, USA, 2002. [Google Scholar]
- CCAMLR. CCAMLR Standard Methods; CCAMLR: Hobart, Australia, 2004. [Google Scholar]
- Pezzo, F.; Olmastroni, S.; Volpi, V.; Focardi, S. Annual Variation in Reproductive Parameters of Adélie Penguins at Edmonson Point, Victoria Land, Antarctica. Polar Biol. 2007, 31, 39–45. [Google Scholar] [CrossRef]
- Taylor, R.H.; Wilson, P.R.; Thomas, B.W. Status and Trends of Adélie Penguin Populations in the Ross Sea Region. Polar Rec. 1990, 26, 293–304. [Google Scholar] [CrossRef]
- Tenaza, R. Behaviour and Nesting Success Relative to Nest Location in Adélie Penguins (Pygoscelis Adeliae). Condor 1971, 73, 81–92. [Google Scholar] [CrossRef]
- Müller-Schwarz, D.; Müller-Schwarze, C. Differential Predation by South Polar Skuas in an Adélie Penguin Rookery. Condor 1973, 75, 127–131. [Google Scholar] [CrossRef]
- Trillmich, F. Feeding Territories and Breeding Success of South Polar Skuas. Auk 1978, 95, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Pezzo, F.; Olmastroni, S.; Corsolini, S.; Focardi, S. Factors Affecting the Breeding Success of the South Polar Skua Catharacta Maccormicki at Edmonson Point, Victoria Land, Antarctica. Polar Biol. 2001, 24, 389–393. [Google Scholar] [CrossRef]
- Southwell, C.; Emmerson, L. Remotely-Operating Camera Network Expands Antarctic Seabird Observations of Key Breeding Parameters for Ecosystem Monitoring and Management. J. Nat. Conserv. 2015, 23, 1–8. [Google Scholar] [CrossRef]
- Southwell, C.; Barbosa, A.; Emmerson, L.; Hart, T.; Hinke, J.; Juàres, M.; Korczak-Abshire, M.; Milinevsky, G.; Newbery, K.; Olmastroni, S.; et al. Remotely Operating Camera Network Provides Spatially Extensive, Long-Term Observations of Breeding Pygoscelis Penguins around Antarctica. In Proceedings of the Marine Ecosystem Assessment for the Southern Ocean, Hobart, Australia, 9 April 2018. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R, 1st ed.; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87458-6. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Mac Nally, R.; Duncan, R.P.; Thomson, J.R.; Yen, J.D.L. Model Selection Using Information Criteria, but Is the “Best” Model Any Good? J. Appl. Ecol. 2018, 55, 1441–1444. [Google Scholar] [CrossRef]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.D.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A Brief Introduction to Mixed Effects Modelling and Multi-Model Inference in Ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartòn, K. MuMIn: Multi-Model Inference. R Package Version 1.15.6. Available online: https://cran.r-project.org/web/packages/MuMIn (accessed on 15 December 2021).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R. J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Carlini, A.; Coria, N.; Santos, M.; Libertelli, M.; Donini, G. Breeding Success and Population Trends in Adélie Penguins in Areas with Low and High Levels of Human Disturbance. Polar Biol. 2007, 30, 917–924. [Google Scholar] [CrossRef]
- Irvine, L.; Clarke, J.; Kerry, K. Low Breeding Success of the Adélie Penguin at Béchervaise Island in the 1998/99 Season. CCAMLR Sci. 2000, 7, 151–167. [Google Scholar]
- Ainley, D.G.; Ribic, C.A.; Ballard, G.; Heath, S.; Gaffney, I.; Karl, B.J.; Barton, K.R.; Wilson, P.R. Geographic Structure of Adélie Penguin Populations: Size Overlap and Use of Adjacent Colony-Specific Foraging Areas. Ecol. Monogr. 2004, 74, 159–178. [Google Scholar] [CrossRef]
- Williams, T.D.; Croxall, J.P. Is Chick Fledging Weight a Good Index of Food Availability in Seabird Populations? Oikos 1990, 59, 414–416. [Google Scholar] [CrossRef]
- Jafari, V.; Maccapan, D.; Careddu, G.; Sporta Caputi, S.; Calizza, E.; Rossi, L.; Costantini, M.L. Spatial and Temporal Diet Variability of Adélie (Pygoscelis Adeliae) and Emperor (Aptenodytes Forsteri) Penguin: A Multi Tissue Stable Isotope Analysis. Polar Biol. 2021, 44, 1869–1881. [Google Scholar] [CrossRef]
- Spurr, E.B. Individual Differences in Aggressiveness of Adélie Penguins. Anim. Behav. 1974, 22, 611–616. [Google Scholar] [CrossRef]
- Davis, L.S.; McCaffrey, F.T. Survival Analysis of Eggs and Chicks of Adélie Penguins (Pygoscelis Adeliae). Auk 1986, 103, 379–388. [Google Scholar] [CrossRef]
- Aebischer, N.J.; Coulson, J.C. Survival of the Kittiwake in Relation to Sex, Year, Breeding Experience and Position in the Colony. J. Anim. Ecol. 1990, 59, 1063–1071. [Google Scholar] [CrossRef]
- Minias, P. Evolution of Within-Colony Distribution Patterns of Birds in Response to Habitat Structure. Behav. Ecol. Sociobiol. 2014, 68, 851–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gochfeld, M.C.F. Timing of Breeding and Chick Mortality in Central and Peripheral Nests of Magellanic Penguins. Auk 1980, 97, 191–193. [Google Scholar] [CrossRef]
- Marks, E.J.; Rodrigo, A.G.; Brunton, D.H. Using Logistic Regression Models to Predict Breeding Success in Male Adélie Penguins (Pygoscelis Adeliae). Polar Biol. 2010, 33, 1083–1094. [Google Scholar] [CrossRef]
- Olmastroni, S.; Pezzo, F.; Volpi, V.; Focardi, S. Effects of Weather and Sea-Ice on the Reproductive Performance of the Adélie Penguins at Edmonson Point, Ross Sea. CCAMLR Sci. 2004, 11, 99–109. [Google Scholar]
- Ropert-Coudert, Y.; Kato, A.; Shiomi, K.; Barbraud, C.; Angelier, F.; Delord, K.; Poupart, T.; Koubbi, P.; Raclot, T. Two Recent Massive Breeding Failures in an Adélie Penguin Colony Call for the Creation of a Marine Protected Area in D’Urville Sea/Mertz. Front. Mar. Sci. 2018, 5, 264. [Google Scholar] [CrossRef] [Green Version]
- Fraser, W.R.; Patterson-Fraser, D.L.; Ribic, C.A.; Schofield, O.; Ducklow, H. A Nonmarine Source of Variability in Adélie Penguin Demography. Oceanography 2013, 26, 207–209. [Google Scholar] [CrossRef] [Green Version]
- McDowall, P.S.; Lynch, H.J. When the “selfish herd” becomes the “frozen herd”: Spatial dynamics and population persistence in a colonial seabird. Ecology 2019, 100, e02823. [Google Scholar] [CrossRef]
- Morrison, K.W.; Armstrong, D.P.; Battley, P.F.; Jamieson, S.E.; Thompson, D.R. Predation by New Zealand Sea Lions and Brown Skuas Is Causing the Continued Decline of an Eastern Rockhopper Penguin Colony on Campbell Island. Polar Biol. 2017, 40, 735–751. [Google Scholar] [CrossRef]
- Ancel, A.; Beaulieu, M.; Gilbert, C. The Different Breeding Strategies of Penguins: A Review. C. R. Biol. 2013, 336, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southwell, C.; Emmerson, L.; Takahashi, A.; Barbraud, C.; Delord, K.; Weimerskirch, H. Large-Scale Population Assessment Informs Conservation Management for Seabirds in Antarctica and the Southern Ocean: A Case Study of Adélie Penguins. Glob. Ecol. Conserv. 2017, 9, 104–115. [Google Scholar] [CrossRef]
- Tin, T.; Fleming, Z.L.; Hughes, K.A.; Ainley, D.G.; Convey, P.; Moreno, C.A.; Pfeiffer, S.; Scott, J.; Snape, I. Impacts of Local Human Activities on the Antarctic Environment. Antarct. Sci. 2009, 21, 3–33. [Google Scholar] [CrossRef] [Green Version]
- Woehler, E.J.; Ainley, D.; Jabour, J. Human Impacts to Antarctic Wildlife: Predictions and Speculations for 2060. In Antarctic Futures Human Engagement with the Antarctic Environment; Tina Tin, D.L., Ed.; Springer Science & Business Media: Dordrecht, The Netherlands, 2014; ISBN 978-94-007-6582-5. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. of Nests | Colony Area (km2) | Skua Nests c | Linear Fast Ice Extension (max–min, km) | Linear Distance to Polynya (km) | |
|---|---|---|---|---|---|
| EdPo | 2704 a | 0.03 | 105; 78 | 36–13; 32–13 | 44 |
| AdCo | 11,438 b | 0.06 | 25; 30 | 5–0; 2–0 | 0 |
| InIs | 29,899 b | 0.25 | 29; 35 | 3.5–0; 0.5–0 | 0 |
| 2017/2018 | 2018/2019 | |||||
|---|---|---|---|---|---|---|
| AdCo | EdPo | Test Statistic | AdCo | EdPo | Test Statistic | |
| Wind speed (knots) | 9.36 ± 0.75 | 4.12 ± 0.20 | r = 58 (p < 0.001) * | 6.01 ± 0.41 | 6.36 ± 0.54 | r = 39 (p = 0.055) |
| Temperature (°C) | −4.98 ± 0.22 | −3.11 ± 0.25 | r = 55 (p < 0.001) * | −4.21 ± 0.24 | −1.96 ± 0.27 | r = 58 (p < 0.001) * |
| Windchill (°C) | −12.35 ± 0.37 | −8.57 ± 0.24 | r = 59 (p < 0.001) * | −12.42 ± 0.41 | −8.17 ± 0.23 | r = 59 (p < 0.001) * |
| Relative humidity (%) | 62.37 ± 1.57 | 56.83 ± 1.72 | r = 40 (p = 0.030) * | 62.17 ± 1.74 | 61.18 ± 2.10 | r = 37 (p = 0.162) |
| Breeding Output Index | Predictor | β Coefficient | 95% CI |
|---|---|---|---|
| Hatching success | Intercept | 1.659 | 1.102; 2.216 * |
| Nest position (peripheral) | −0.931 | −1.533; −0.328 * | |
| Year (2018/2019) | 0.766 | 0.274; 1.259 * | |
| Breeding success | Intercept | 0.776 | 0.381; 1.171 * |
| Skuas’ nests in 50 m | −0.523 | −0.767; −0.278 * | |
| Nest position (peripheral) | −0.494 | −0.961; −0.027 * | |
| Days elapsed from hatching date | −0.230 | −0.444; −0.016 * | |
| Mean daily windchill (°C) from hatching date | 0.181 | −0.063; 0.426 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olmastroni, S.; Ferretti, F.; Burrini, L.; Ademollo, N.; Fattorini, N. Breeding Ecology of Adélie Penguins in Mid Victoria Land, Ross Sea Antarctica. Diversity 2022, 14, 429. https://doi.org/10.3390/d14060429
Olmastroni S, Ferretti F, Burrini L, Ademollo N, Fattorini N. Breeding Ecology of Adélie Penguins in Mid Victoria Land, Ross Sea Antarctica. Diversity. 2022; 14(6):429. https://doi.org/10.3390/d14060429
Chicago/Turabian StyleOlmastroni, Silvia, Francesco Ferretti, Lucia Burrini, Nicoletta Ademollo, and Niccolò Fattorini. 2022. "Breeding Ecology of Adélie Penguins in Mid Victoria Land, Ross Sea Antarctica" Diversity 14, no. 6: 429. https://doi.org/10.3390/d14060429