
Molecular Characterization of Ctenotaenia marmotae (Frölich, 1802) Railliet, 1893 (Cyclophyllidea: Anoplocephalidae) Parasitizing Rodents of the Genus Marmota and Spermophilus from Eurasia


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beveridge, I. Family Anoplocephalidae Cholodkovsky, 1902. In Keys to the Cestode Parasites of Vertebrates; Khalil, L.F., Jones, A., Bray, R.A., Eds.; CAB International: Wallingford, UK, 1994; pp. 315–366. [Google Scholar]
- Wilson, D.E.; Reederm, D.M. (Eds.) Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2005. [Google Scholar]
- Preleuthner, M.; Calderola, S.; Lanfranchi, P.; Prosl, H. Parasiten des Alpenmurmeltieres (Marmota marmota): Systematik, Entwicklung, Verbreitung. Stapfia 1999, 63, 77–92. [Google Scholar]
- Ryzhikov, K.M.; Gvozdev, E.V.; Tokobaev, M.M.; Shaldybin, L.S.; Macaberidze, G.V.; Merkusheva, I.V.; Nadtochij, E.V.; Hohlova, I.G.; Sharpilo, L.D. Keys to the Helminths of The Rodent Fauna of the USSR. Cestodes and Trematodes; Nauka: Moscow, Russia, 1978. (In Russian) [Google Scholar]
- Beveridge, I. A taxonomic revision of the genera Cittotaenia Riehm, 1881, Ctenotaenia, Railliet, 1893, Mosgovoyia Spasskii, 1951 and Pseudocittotaenia Tenora, 1976. (Cestoda: Anoplocephalidae). Mem. Mus. Natl. D’hist. Nat. Ser. A Zool. 1978, 107, 1–64. [Google Scholar]
- Haukisalmi, V.; Hardman, L.M.; Hoberg, E.P.; Henttonen, H. Phylogenetic relationships and taxonomic revision of Paranoplocephala Lühe, 1910 sensu lato (Cestoda, Cyclophyllidea, Anoplocephalidae). Zootaxa 2014, 3873, 371–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haukisalmi, V.; Wickström, L.M.; Henttonen, H.; Hantula, J.; Gubányi, A. Molecular and morphological evidence for multiple species within Paranoplocephala omphalodes (Cestoda, Anoplocephalidae) in Microtus voles (Arvicolinae). Zool. Scr. 2004, 33, 277–290. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Henttonen, H.; Hardman, L.M. Taxonomy and diversity of Paranoplocephala spp. (Cestoda: Anoplocephalidae) in voles and lemmings of Beringia, with a description of three new species. Biol. J. Linn. Soc. 2006, 89, 277–299. [Google Scholar] [CrossRef] [Green Version]
- Vlasenko, P.; Abramov, S.; Bugmyrin, S.; Dupal, T.; Fomenko, N.; Gromov, A.; Zakharov, E.; Ilyashenko, V.; Kabdolov, Z.; Tikunov, A.; et al. Geographical distribution and hosts of the cestode Paranoplocephala omphalodes (Hermann, 1783) Lühe, 1910 in Russia and adjacent territories. Parasitol. Res. 2019, 118, 3543–3548. [Google Scholar] [CrossRef] [PubMed]
- Kachanovsky, I.M.; Nikiforov, M.E.; Parfenov, V.I. (Eds.) Red Data Book of the Republic of Belarus: Rare and Threatened Species of Wild Animals; Petrus Brovka Belarusian Encyclopedia: Minsk, Belarus, 2015. [Google Scholar]
- Haukisalmi, V.; Hardman, L.M.; Foronda, P.; Feliu, C.; Henttonen, H. Systematic relationships of Mosgovoyia Spasskii, 1951 (Cestoda: Anoplocephalidae) and related genera inferred from mitochondrial and nuclear sequence data. Syst. Parasitol. 2010, 77, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Multiple Sequence Alignment Methods. Methods in Molecular Biology; Russell, D.J., Ed.; Humana Press: Totowa, MJ, USA, 2014; Volume 1079, pp. 105–116. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Krivopalov, A.; Vlasenko, P.; Abramov, S.; Akimova, L.; Barkhatova, A.; Dokuchaev, N.; Gromov, A.; Konyaev, S.; Lopatina, N.; Vlasov, E.; et al. Distribution and Molecular Diversity of Paranoplocephala kalelai (Tenora, Haukisalmi & Henttonen, 1985) Tenora, Murai & Vaucher, 1986 in Voles (Rodentia: Myodes) in Eurasia. Diversity 2022, 14, 472. [Google Scholar] [CrossRef]
- Gromov, I.M.; Bibikov, D.I.; Kalabuchov, N.I.; Mejer, M.H. Fauna SSSR. Mlekopitajuscie [Mammalia]. Nazemnye belich’i (Marmotinae). Tom 3, Vyp. 2; Nauka: Moskva, Russia; Leningrad, Russia, 1995; 464p. (In Russian) [Google Scholar]
- Schmid-Hempel, P. Sociality and parasite transmission. Behav. Ecol. Sociobiol. 2021, 75, 156. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Locality (Number) | Coordinates | GenBank Acc. No. (Cox1/Nad1) | Final Host |
|---|---|---|---|
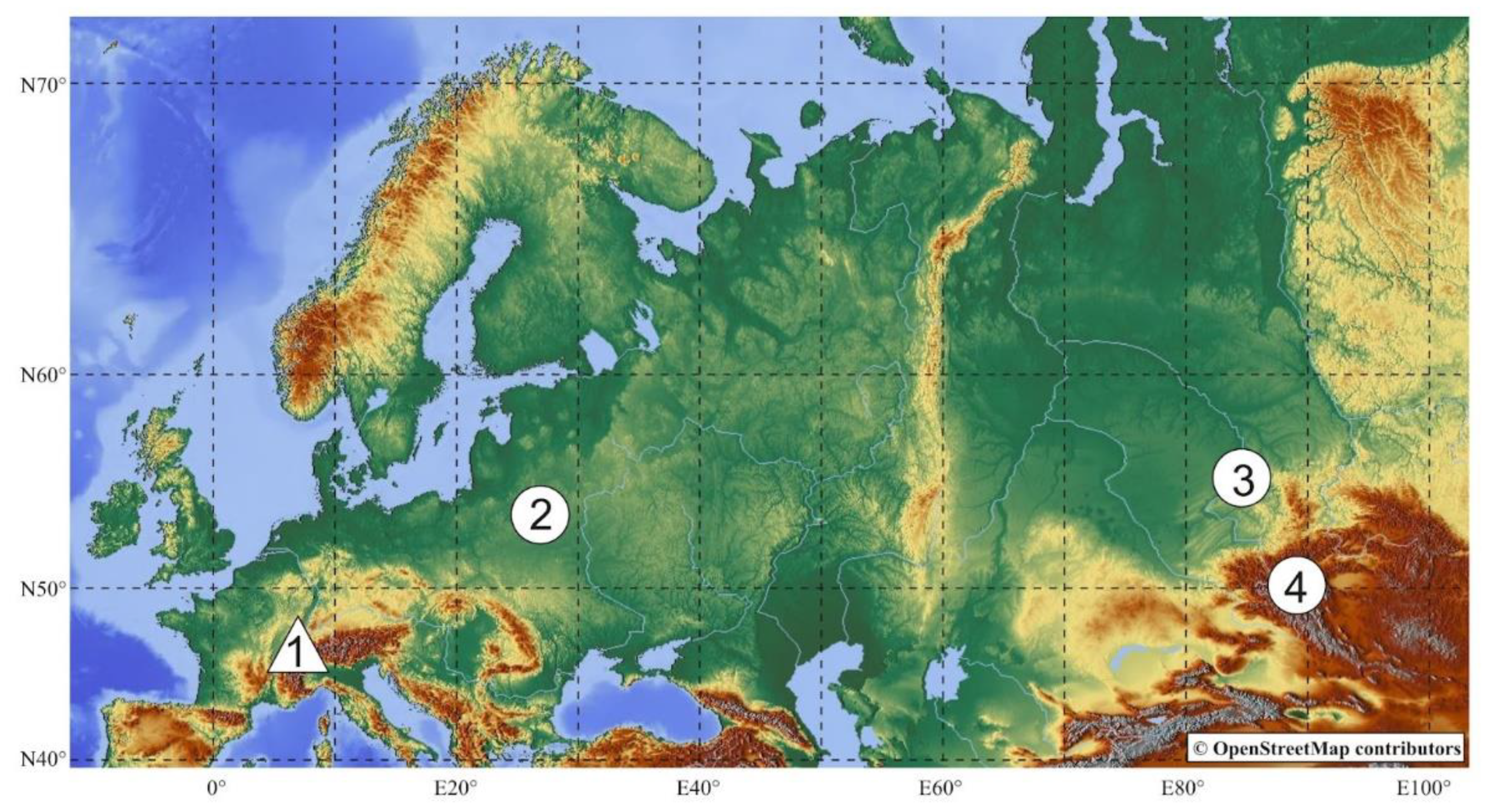
| France (1) | -- | AY568187/HM134271 | Marmota marmota (Linnaeus, 1758) |
| Nesvizh, Minsk Oblast, Belarus (2) | 53.24530438 N 26.76108783 E | ON459626/ON505824 | Spermophilus suslicus (Guldenstaedt, 1770) |
| Doronino, Novosibirsk Oblast, Russia (4) | 54.98292101 N 84.58460474 E | ON459627/ON505825 | Spermophilus erythrogenys Brandt, 1841 |
| Ulandryk, Altai Republic, Russia (3) | 49.60679776 N 89.07580720 E | ON459628/ON505826 | Marmota baibacina Kastschenko, 1899 |
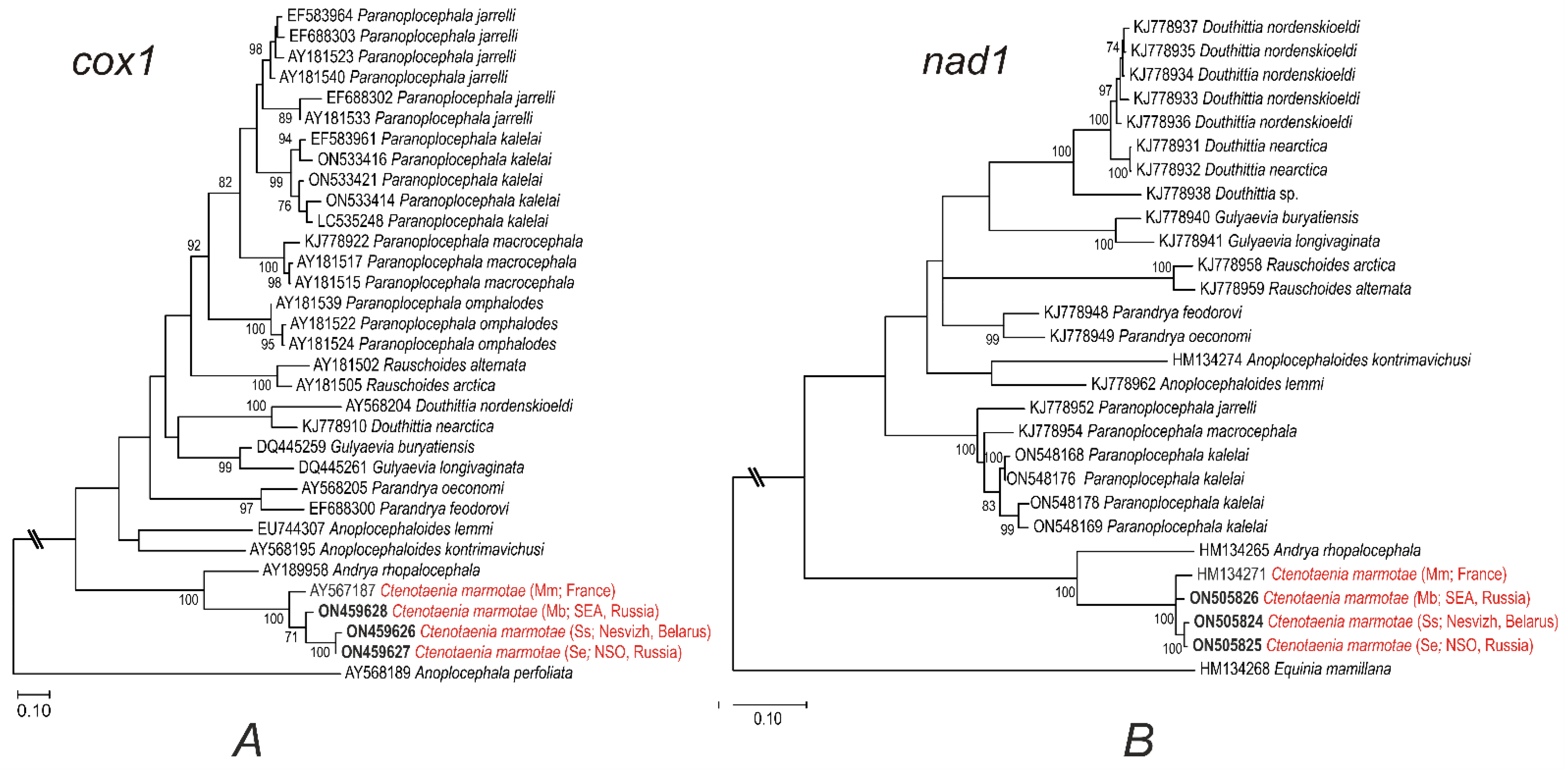
| Cestode Species | Cox1 | Nad1 | Final Host |
|---|---|---|---|
| Paranoplocephala jarrelli | AY181523, AY181540, AY181533, EF688302, EF688303, EF583964 | KJ778952 | Microtus oeconomus |
| Paranoplocephala kalelai | EF583961, ON533421, ON533414, ON533416, LC535248 | ON548168, OM548169, ON548176, ON548178 | Myodes rufocanus |
| Paranoplocephala macrocephala | KJ778922, AY1811515, | KJ778954 | Microtus longicaudus |
| AY181517 | Microtus pennsylvanicus | ||
| Paranoplocephala omphalodes | AY18122, AY18124, | Microtus agrestis | |
| AY18139 | Myodes glareolus | ||
| Rauschoides alternata | AY181502 | KJ778959 | Dicrostonyx groenlandicus |
| Rauschoides arctica | AY181505 | KJ778958 | Dicrostonyx groenlandicus |
| Gulyaevia buryatiensis | DQ445259 | KJ778940 | Myodes rufocanus |
| Gulyaevia longivaginata | DQ445261 | KJ778941 | Myodes rutilus |
| Douthittia nordenskioeldi | AY568204 | KJ778933-KJ778937 | Microtus miurus |
| Douthittia nearctica | KJ778910 | KJ778931, KJ778932 | Myodes rutilus |
| Douthitta sp. | KJ778938 | Myodes rutilus | |
| Anoplocephaloides lemmi | EU744307 | KJ778962 | Lemmus sibiricus |
| Anoplocephaloides kontrimavichusi | AY568195 | HM134274 | Synaptomys borealis |
| Parandrya oeconomi | AY568205 | KJ778949 | Microtus oeconomus |
| Parandrya feodorovi | EF688300 | KJ778948 | Microtus sp. |
| Andrya rhopalocephala | AY189958 | HM134265 | Lepus europaeus |
| Equinia mamilliana | --- | HM134268 | Equus caballus |
| Anoplocephala perfoliata | AY568189 | --- | Equus caballus |
| Host M. baibacina (Altai, Russia) | Host M. marmota (Alps, France) | Host S. erythrogenus (West Siberia, Russia) | Host S. suslicus (Belarus) | |
|---|---|---|---|---|
| Host M. baibacina (Altai, Russia) | 96.6 | 97.2 | 96.6 | |
| Host M. marmota (Alps, France) | 97.7 | 95.2 | 94.8 | |
| Host S. erythrogenus (West Siberia, Russia) | 98.0 | 97.2 | 99.5 | |
| Host S. suslicus (Belarus) | 97.7 | 96.9 | 99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krivopalov, A.; Abramov, S.; Akimova, L.; Barkhatova, A.; Gromov, A.; Konyaev, S.; Lopatina, N.; Sidorovich, A.; Vlasov, E.; Vlasenko, P.; et al. Molecular Characterization of Ctenotaenia marmotae (Frölich, 1802) Railliet, 1893 (Cyclophyllidea: Anoplocephalidae) Parasitizing Rodents of the Genus Marmota and Spermophilus from Eurasia. Diversity 2022, 14, 531. https://doi.org/10.3390/d14070531
Krivopalov A, Abramov S, Akimova L, Barkhatova A, Gromov A, Konyaev S, Lopatina N, Sidorovich A, Vlasov E, Vlasenko P, et al. Molecular Characterization of Ctenotaenia marmotae (Frölich, 1802) Railliet, 1893 (Cyclophyllidea: Anoplocephalidae) Parasitizing Rodents of the Genus Marmota and Spermophilus from Eurasia. Diversity. 2022; 14(7):531. https://doi.org/10.3390/d14070531
Chicago/Turabian StyleKrivopalov, Anton, Sergey Abramov, Lyudmila Akimova, Alina Barkhatova, Anton Gromov, Sergey Konyaev, Natalia Lopatina, Anna Sidorovich, Egor Vlasov, Pavel Vlasenko, and et al. 2022. "Molecular Characterization of Ctenotaenia marmotae (Frölich, 1802) Railliet, 1893 (Cyclophyllidea: Anoplocephalidae) Parasitizing Rodents of the Genus Marmota and Spermophilus from Eurasia" Diversity 14, no. 7: 531. https://doi.org/10.3390/d14070531
APA StyleKrivopalov, A., Abramov, S., Akimova, L., Barkhatova, A., Gromov, A., Konyaev, S., Lopatina, N., Sidorovich, A., Vlasov, E., Vlasenko, P., & Zinchenko, V. (2022). Molecular Characterization of Ctenotaenia marmotae (Frölich, 1802) Railliet, 1893 (Cyclophyllidea: Anoplocephalidae) Parasitizing Rodents of the Genus Marmota and Spermophilus from Eurasia. Diversity, 14(7), 531. https://doi.org/10.3390/d14070531