1. Introduction
On decadal time scales, the Ross Sea has been experiencing changes in both its physical and biological conditions. In the last 60 years, the Ross Sea shelf waters have undergone a continuing near-linear temperature increase and salinity decrease [
1], although a rebound in salinity was observed from 2016 to 2019 [
2,
3]. Similar interannual variability has been observed in sea ice concentration that has increased from 1979 to 2014 [
4] and decreased during the summer after 2015 [
3].
Thermohaline changes are most evident in Terra Nova Bay (TNB) which is located in the western sector of the Ross Sea off Victoria Land and is bounded by the Drygalski Ice Tongue to the south and the Campbell Ice Tongue to the north. Here, the salinity of the Dense Shelf Water (DSW) decreased from 1995 to 2014 by −0.045 ± 0.016 dec
−1, with a change in neutral density from 28.796 to 28.718 kg m
−3 [
2]. After 2014, a sharp increase in salinity has been observed with a rebound to values last observed in the mid-late 1990s [
2]. Superimposed on this trend, Castagno et al. [
2] also observed an interannual variability with 5–10 years fluctuations. Relative minimum values were registered in 2000, 2005, and 2014, whereas relative maximum values were measured in 1995, 2002, 2008, and 2018 [
2,
3]. These salinity anomalies extend throughout the water column [
2], with the surface water showing the same variability as the DSW, (see Figure 3a in Castagno et al. [
2]). DSW temperatures have increased in proportion to the rise in sea surface freezing point that would accompany the observed salinity decrease [
5].
The predominant feature of TNB is a coastal latent heat polynya formed and maintained by katabatic winds [
6]. TNB polynya is bounded by the Drygalski Ice Tongue to the south that acts as a barrier for the pack ice advection from the south [
7], keeping the polynya open during winter. The mean polynya extent during winter ranges from 1000 to 1300 km
2 estimated from thermal infrared data [
6,
7] to about 4200 km
2 using passive microwave data [
8]. However, the polynya extent shows a large interannual variability [
9]. TNB polynya influences the entire water column, the persistent sea-ice production, and export by the katabatic winds during winter results in brine rejection that densifies the water column. TNB water column could be divided into an upper layer of about 150 m characterized by high thermohaline variability and a layer beneath where the water column is nearly isothermal, and the vertical stability is preserved by the increasing salinity [
9]. During winter, from May to October, surface water temperature is close to the freezing point, while salinity increases due to brine rejection in response to sea ice formation [
9]. Starting from November, temperature increases, and salinity decrease due to sea ice melting and the heat gained from the solar radiation [
10]. Due to a large amount of sea ice melting associated with increased springtime temperatures the TNB water column becomes highly stratified in late spring and summer [
11]. Furthermore, in early austral spring, TNB polynya greatly increases in size [
12], and hosts large seasonal phytoplankton blooms, typically dominated by the colonial haptophyte
Phaeocystis antarctica in spring through early summer, with an increase in abundance of diatoms in mid to late summer [
11,
12,
13,
14,
15,
16]. The summer diatom blooms are documented by a European Long Term Ecological Research (LTER) data, initiated in 2006 near the Italian station Mario Zucchelli in TNB (
http://www.lteritalia.it/?q=macrositi/it17-stazioni-di-ricerca-antartide accessed on 21 July 2022) as well as by previous expedition to TNB (from 1987 to 1995) [
17].
Recent studies suggest that future phytoplankton assemblage and productivity will likely increase due to changes in summer sea ice concentrations and hypothesize that shallower mixed layer depths will cause diatoms to dominate the future phytoplankton assemblage relative to
P. antarctica [
18,
19,
20]. The considerable increases in phytoplankton biomass and large size structure suggest that the Ross Sea could now be extremely productive in summer [
20] and these changes in phytoplankton composition, productivity, and export would have implications for the rest of the Ross Sea food web as provide increased energy to grazers and thus increase secondary production.
Microzooplankton organisms (20 µm to 200 µm size range) are pivotal species in the microbial community and they play a fundamental role in the Antarctic food webs [
21,
22]. Microzooplankton herbivory constitutes a major source of mortality for phytoplankton in the ocean [
23,
24,
25] and may thus exert significant top-down control on phytoplankton blooms in the Southern Ocean [
21,
22].
Tintinnid ciliates are part of microzooplankton, which in the Southern Ocean sometimes represent up to 50% of microzooplankton abundance and biomass [
26]. They are characterized by a species-specific shell (lorica), shaped like a bowl or vase, or tube [
27] on which their taxonomy is based. Some tintinnids species are known to display considerable plasticity in lorica morphology, (e.g., [
28,
29,
30]), which has led to a different classification of some species over the years.
In the Southern Ocean, tintinnids appear to be a widely important and exploited food resource. They are feeding primarily on small phytoplankton sizes and, when phytoplankton is not dominated by large diatoms or
Phaeocystis, they are recognized to be the major consumers of primary production. Moreover, a wide variety of animals are known to consume tintinnids. They have been found in the guts of crustacean zooplankters such as copepods [
31], krill and mysid shrimp [
32], salps [
33], chaetognaths [
34], and larval Antarctic silverfish [
35].
The Antarctic waters’ studies on tintinnids date back to the earliest scientific expedition and they were almost exclusively devoted to taxonomic description [
36,
37]. The most recent reports provide quali-quantitative data and they are available for different Antarctic environments [
38,
39,
40,
41,
42,
43]. Despite the relatively abundant data on tintinnids distribution and composition, to the best of our knowledge, there are no long time series on tintinnids in the Antarctic area.
In this paper, we report the first plurennial series on tintinnids collected on eleven occasions in the polynya of TNB over the period 1988–2017. The aim of this study was to describe the tintinnids community in TNB over the past thirty years and analyze if tintinnids were affected by the significant changes in the Ross Sea environment. We hypothesize that the rising ocean temperature, as expected from climate change predictions, might have an effect on tintinnids’ community structure, with ecological consequences in its quantity and diversity, including modification of keystone species.
2. Material and Methods
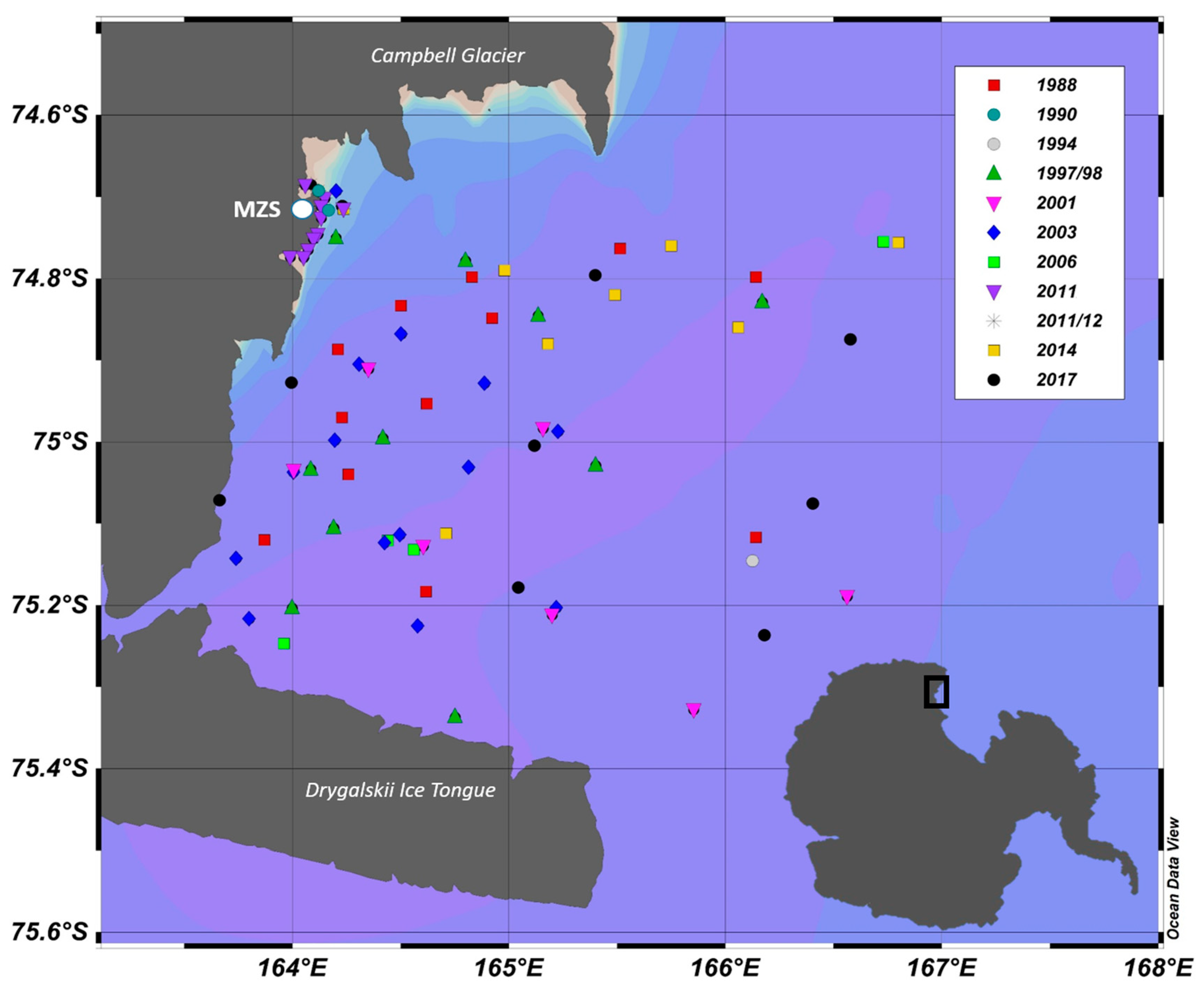
Tintinnids’ community was studied during eleven expeditions in the Ross Sea within the frame of the Italian Project in Antarctica (PNRA) in the Terra Nova Bay (TNB) polynya (74°60–75°40 S and 163°50–167° E), between the Campbell Glacier Tongue and Drygalskii Ice Tongue (
Figure 1). Sampling was performed during the austral summer (December–February) of each expedition.
Table S1 reports the sampling site and number of stations for each year. Samples were collected using 12 L Niskin bottles (Sea-Bird Electronics 32) mounted on a multisampler equipped with a CTD sensor (Meerestechnik Mod. KMS multiparametric; 9/11 Plus, Sea-Bird Electronics, Bellevue, WA, USA). Sea water samples were collected at 3 to 5 depths, relative to the surface, DCM (deep chlorophyll maximum), and intermediate depths depending on the bottom of the sampled site (max sampled depth 1128 m). In total, 321 seawater samples were analyzed. Temperature and salinity values used in the following analyses correspond to the depths where Niskin bottle samples were taken for tintinnids samples.
During the expedition of January–February 1990, we sampled repeatedly at two coastal stations: MER (Mergellina) with a bottom depth of 25 m and SMN (Santa Maria Novella) with 500 m depth. In 1994, only one station was considered in this paper, as the long transect sampled was outside of the Terra Nova polynya. In 2011/12 we sampled repeatedly at six stations: SMN, PTF (Portofino), FAR (Faraglioni), T10, and TER (Tergeste), the last two closers to Mario Zucchelli Station (MZS) (
Table S1).
At each depth, from 2 to 5 L of seawater was reverse-filtered through a 10 µm mesh, in order to reduce the volume to 250 mL and immediately fixed with buffered formaldehyde (1.6% final concentration). Subsamples (50 mL) were then examined in a settling chamber using an inverted microscope (magnification 200×) (Leitz Labovert, Leica DMI 300B, Wetzlar, Germany), according to the Utermöhl method [
44]. The entire surface of the chamber was examined.
Tintinnids encountered were assigned species names based only on lorica morphology. Many species in the Antarctic are considered morphological variants [
29,
30] and molecular analyses could be useful to distinguish between different species. In this study, the samples were fixed with formalin and the tintinnids species identifications were made on the basis of description by Brand [
45,
46], Laackmann [
36,
37], Kofoid and Campbell [
47,
48], Alder [
49], and Petz [
50].
Empty loricae were not differentiated from filled ones because tintinnid protoplasms are attached to the lorica by a fragile strand, which detaches with ease during collection and fixation of the samples.
Salpingella genus showed considerable variability in lorica length that made the identification extremely difficult and uncertain. For this reason, we did not distinguish the different Salpingella species and for the calculation of the biomass we divided this genus into different classes according to the different lengths (from 70 to 200 µm). The oral diameter varied from 15 to 20 µm.
For each species, the biomass was estimated by measuring the linear dimensions of each organism using an eyepiece scale and relating the individual shapes to standard geometric figures. Cell volumes were converted to carbon values using the formula: pgC cell
−1 = µm
3 × 0.053 + 444.5 [
51].
Correlation between biotic and abiotic factors was analyzed using STATISTICA11, StatSoft, TIBCO software, Paloalto, CA, USA and the Spearman correlation index was used to identify the relations between temperature, salinity, and abundance. To test the community composition and the similarity pattern presented by all tintinnids genera, the samples were analyzed on the base of the different depths, years, and coastal distances. The factor “zone” was also considered to divide samples collected in the photic zone (<200 m) from the ones collected in the aphotic one (≥200 m). Multivariate analyses were based on Bray–Curtis similarities index [
52], as calculated from the Log (× + 1). PERMANOVA analysis was carried out to test the effect of the considered variable on the community structure. PCO ordination, nMDS, cluster analysis, SIMPER, and ANOSIM analysis were useful to detect which genera were more involved in the communities’ similarity pattern. The analyses were conducted using the PRIMER v7 software, Plumouth, UK [
53], and the significance level for all statistical tests was set at 5%.
3. Results
3.1. Tintinnid Abundance and Biomass
The results of multivariate analyses, performed on the entire dataset, point out a significant (PERMANOVA
p < 0.05) effect of the factor season (early: December, late: January and February) and the factor layer (photic: <200 m; apothic: ≥200 m), in shaping the community composition. Concerning seasonality, pair-wise test and ANOSIM outputs detected significant (
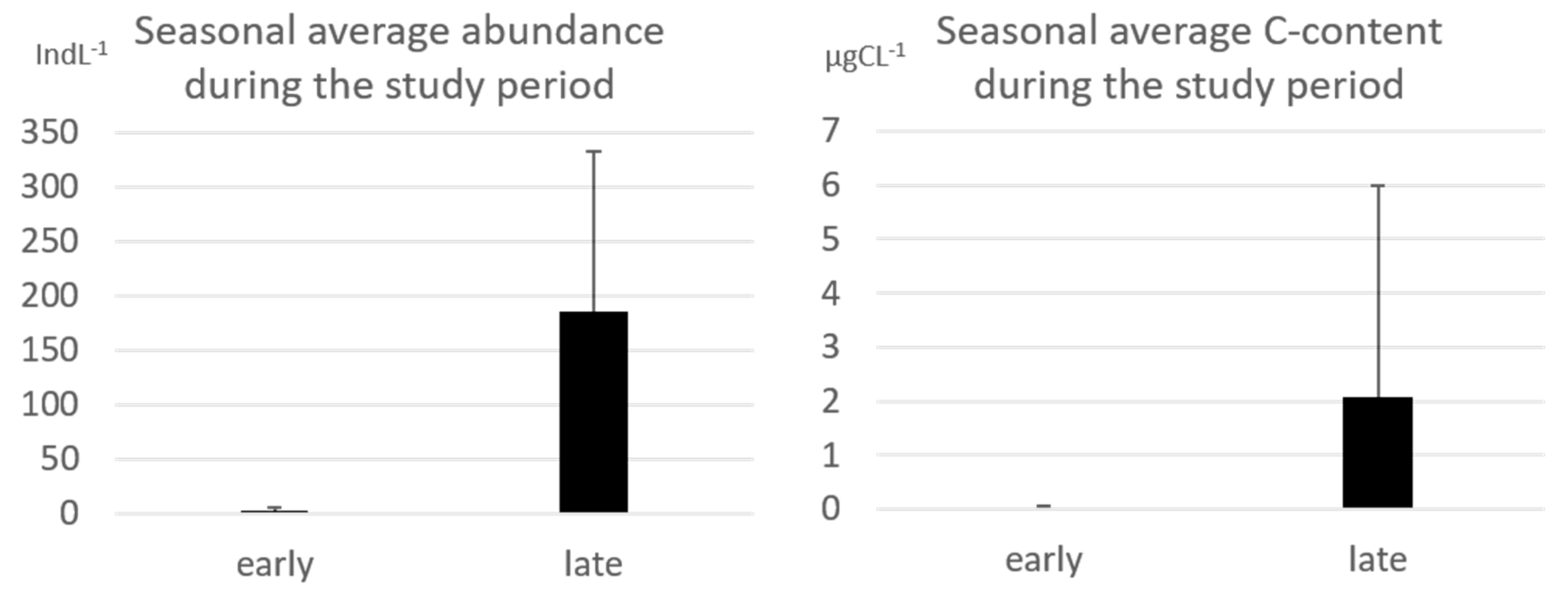
p < 0.05) differences between the communities of the two groups: diversity was mainly due to the higher average abundance and carbon content of the considered taxa. During early summer, the average abundance and the carbon content of all taxa were two orders less than those measured during the late summer (
Figure 2).
The maximum abundance detected during the early summer (December 2011) was 14 indL
−1 at St. Tergeste (TER) while during late summer (January 2014) the maximum value was 4980 indL
−1 at St. 39 (
Table S2). As well the carbon content presented higher values in the late summer, with a maximum value of 36 µgCL
−1 (St. 39 January 2014).
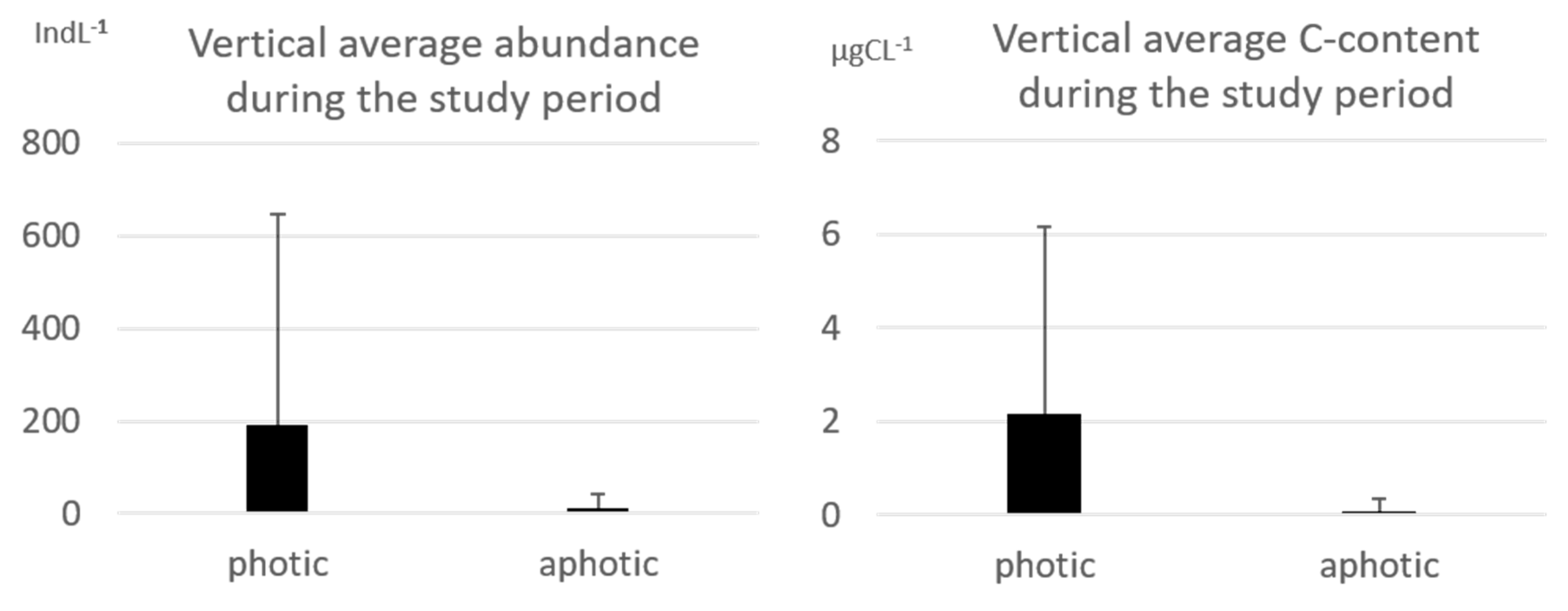
Considering all the periods and the entire study area, the abundances were higher in the photic zone (<200 m), showing the average abundance of 192 ± 454 indL
−1 while in the aphotic zone (≥200 m) they showed a lower value of 10 ± 30 indL
−1 (
Figure 3). The same trend was detected for the carbon content attesting at the average value of 2.1 ± 3.3 µgCL
−1 in the photic zone.
All the genera considered in this work showed the minimum average abundance in the aphotic zone. Pair-wise test and ANOSIM outputs detected a significant (p < 0.05) difference between the communities detected in the two zones highlighting how the aphotic layers (d ≥ 200 m) showed the lowest value in similarity because of the scarce abundances. In order to reduce the variability in the study and to detect the long-term changes in the tintinnids community, only the samples collected in the photic layer (d < 200 m) were considered in the analysis of the genera composition.
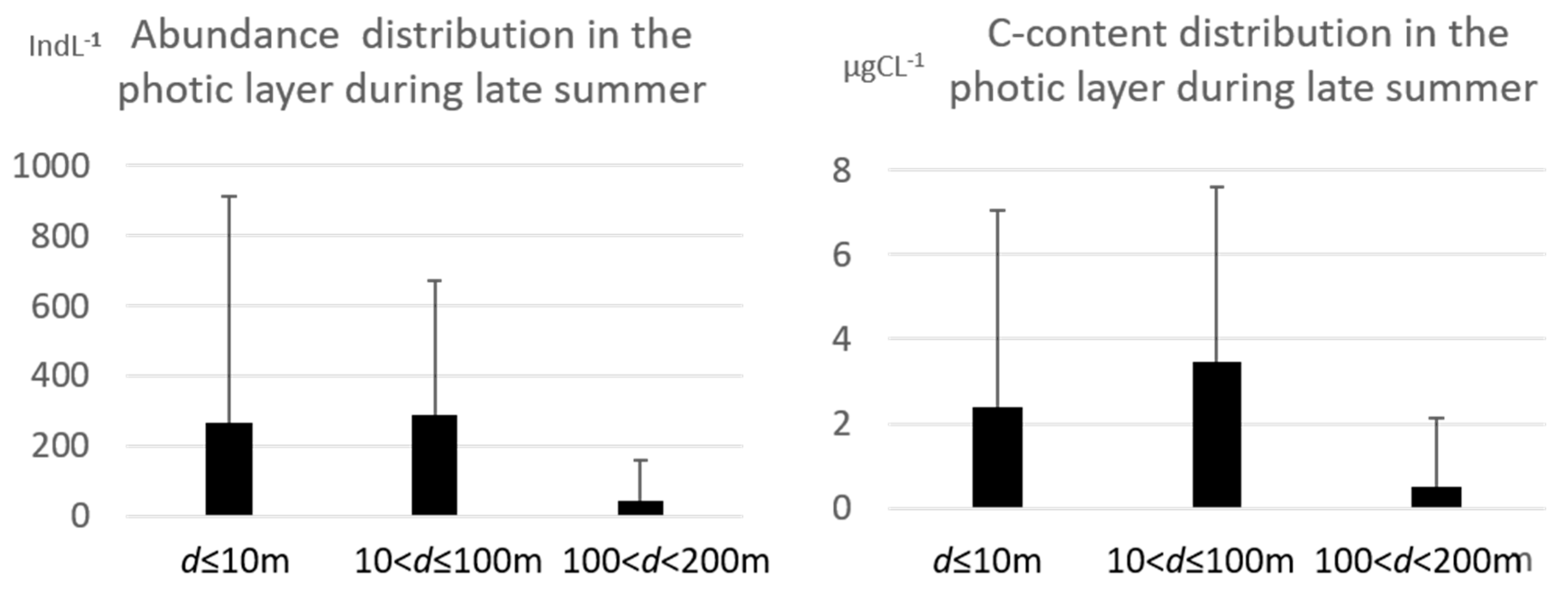
The abundance distribution in the upper 200 m of the water column showed a similar average abundance between the surface layer and the intermediate one (10 <
d ≤ 100 m) with values of 267 ± 642 indL
−1 and 287 ± 384 indL
−1, respectively (
Figure 4).
The maximum abundance peak was recorded at surface with 4980 indL−1. The maximum abundance at the intermediate depth was 1566 indL−1, while at the 100 < d < 200 m layer the average abundances sharply decreased (MAX = 854 indL−1) at 46 ± 115 indL−1.
The carbon content showed the highest average value at the 10 < d ≤ 100 m layer (3.5 ± 4.1 µgCL−1) thanks to the presence of the genera Cymatocylis and Laackmnniella.
3.2. Tintinnids Composition
In total, 13 taxa, corresponding to 9 species from 5 genera, were identified (
Table 1).
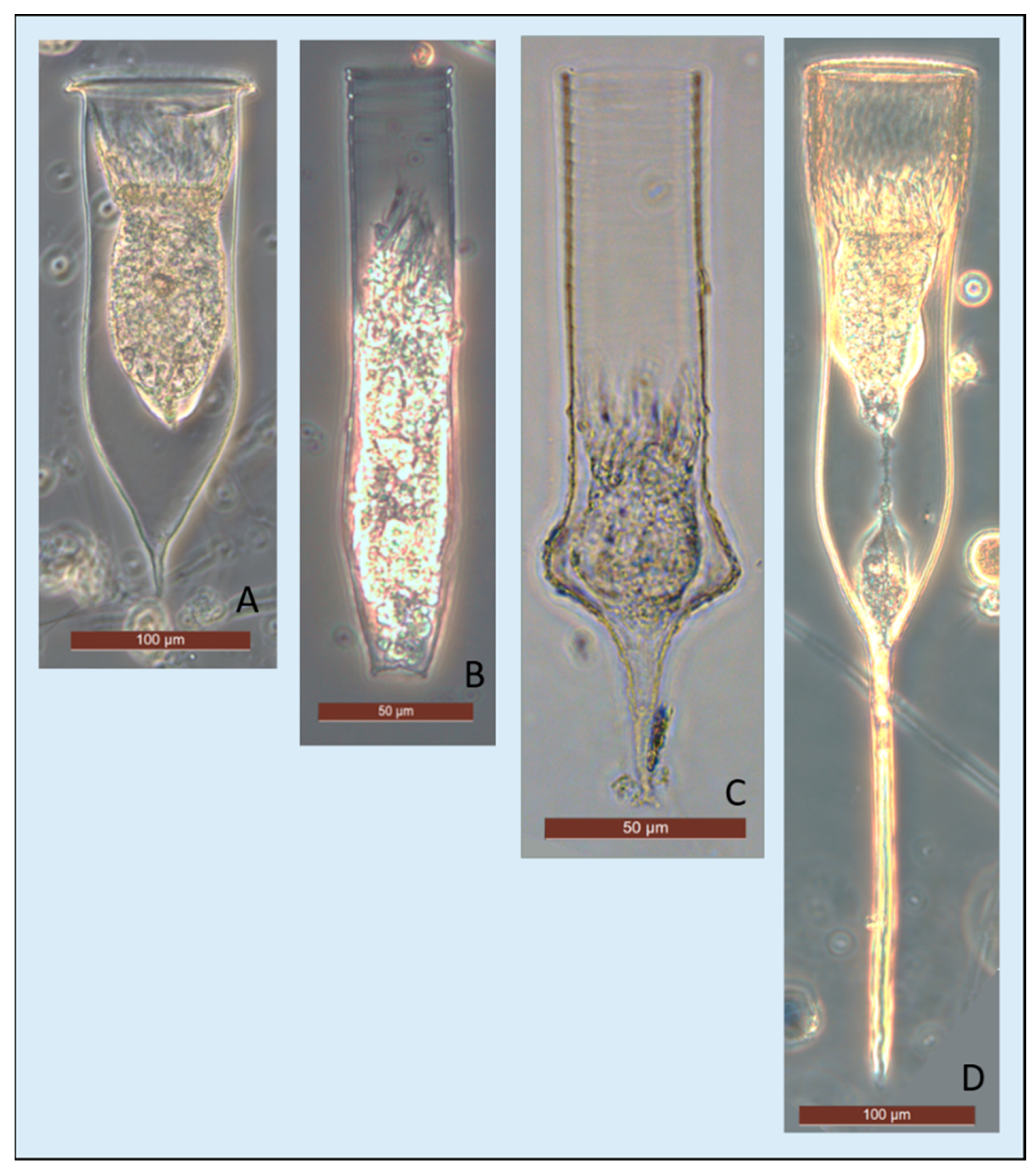
The most representative species were
C. drygalskii (
Figure 5A),
L. naviculaefera (
Figure 5B), and
C. gaussi (
Figure 5C). These three species constituted more than 56% of the tintinnids and they were present in all the years, except in 1994 (sampled only one station) and, for
C. gaussi, also in 2012 (
Table 2). In the last two periods (2014 and 2017),
C. drygalskii,
L. naviculaefera,
C. gaussi and
Salpingella genus increased and reached high abundances. In particular, in 2014
Salpingella contributed together with
C. drygalskii and
C. gaussi, to the highest abundance (4980 indL
−1) of all the investigated periods. In this contest,
Salpingella showed the value of 3250 indL
−1, the highest ever recorded in this time series.
Among the
Cymatocylis genus,
C. drygalskii was the most common species. It reached a maximum of 1100 indL
−1 in 2014 (St. 39, 0 m).
Cymatocylis drygalskii presented a wall hyaline cylindrical shape with different lengths of the lorica (180–340 µm), mainly due to the antapical horn (5–100 µm). The collar rim was serrated and bent downwards, and the external diameter of the bowl was quite constant (around 90 µm) and poorly correlated with lorica length. Among the
Cymatocylis genus,
C. vanhöffeni (
Figure 5D) was the second well-represented species, detected with a maximum abundance of 154 indL
−1 in 1988 (St. 22, 100 m), especially below 50 m. The bowl elongated hyaline lorica shows an apical region of lorica striated. The size varied from around 400 µm long and 90 µm wide, with a long antapical horn (94–230 µm). The rim was serrated without an outer collar and external opening diameter.
Cymatocylis convallaria,
C.
cristallina, and
C. nobilis were very rare, normally detected in the upper 50 m and with maxima abundance < 15 indL
−1.
Among the genus Codenollepsis, C. gaussi was the most abundant species with a maximum of 1003 indL−1 in 1988 (St. 32, 0 m). This species presented a short lorica around 130 µm long and 40 µm wide. Codonellopsis glacialis, was not present in all the years and reached the maximum abundance (81 indL−1) in 1997 (St. 214, 0 m). This species presented a size range of around 95 µm in length and 25 µm in width, slightly agglutinated lorica with a bullet-shaped and hyaline annulated short collar that corresponds to the width.
Laackmanniella naviculaefera, with a maximum abundance of 535 indL−1 in 1988 (St. 26, 50 m), presented a cylindrical annulated hyaline lorica sizing around 200 µm long and a very constant opening diameter (around 45 µm).
Amphorides laackmanni was detected only during the last two campaigns (2014 and 2017) with maxima values of 133 indL−1 (St. 15b, 25 m). Amphorides laackmanni presented a hyaline wall with few longitudinal fins, very short body sizing around 60 µm long and 20 µm opening diameter. This species, like the genus Salpingella, is not as peculiar to Antarctic waters.
3.3. Tintinnid Community Structure
Analyzing the pattern regulating the genera composition and the genera covariance, PERMANOVA analyses evidence of the influence of the factors “depth layer” in shaping the composition of tintinnids assemblage, in this contest significant (p < 0.05) differences were highlighted among the three layers in the photic zone.
The vertical distribution in the study area, resumed in
Table 3, is characterized by the genera
Cymatocylis and
Laackmaniella that show the highest occurrence in the three layers. The average abundances and the frequencies of these two genera followed the pattern of total abundance increasing from the surface to the intermediate layer and decreasing in the 100 <
d < 200 m layer. The genus
Amphorides was absent at 0–10 m while
Salpingella and
Codonellopsis showed higher abundances and frequencies on the surface following a decreasing trend with the depth.
In regard to the horizontal distribution (
Table 4) the “on/off-shore” factors result is significant in the PERMANOVA analysis (
p < 0.05), evidencing different community compositions in the samples collected near the coast (on-shore) from those collected at the intermediate distance (int) or in the open waters (off-shore). In general, the off-shore samples showed higher abundance for the most frequent genera while
Amphorides showed higher values in abundance and occurrence near the coast.
Cymatocylis and
Laackmaniella were more frequent near the coast while
Salpingella and
Codonellopsis followed the same trend increasing with the coastal distance.
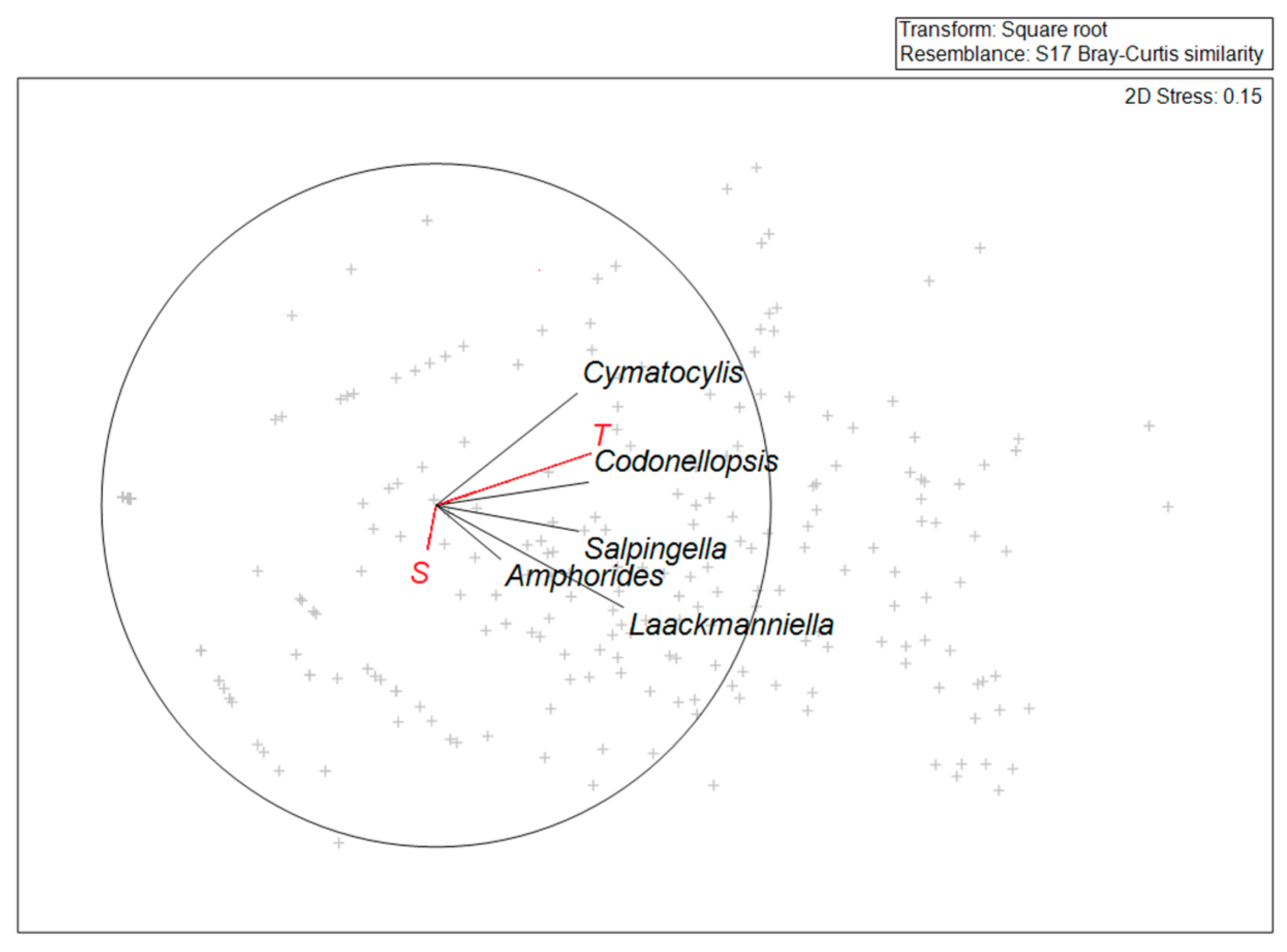
Analyzing the relation between the considered genera and the abiotic factors (temperature and salinity), the vectors presented in
Figure 6 showed the biotic variables positively related to temperature, while there is no relationship to salinity.
Spearman correlation index values indicated the significance (
p < 0.05) of the correlation between temperature and all the genera, except
Amphorides (
Table 5). The negative correlation of
Amphorides was probably due to the lower frequency in the observation of this genus. Furthermore, the total abundance of tintinnids showed a higher value in the correlation index with temperature (
Table 5).
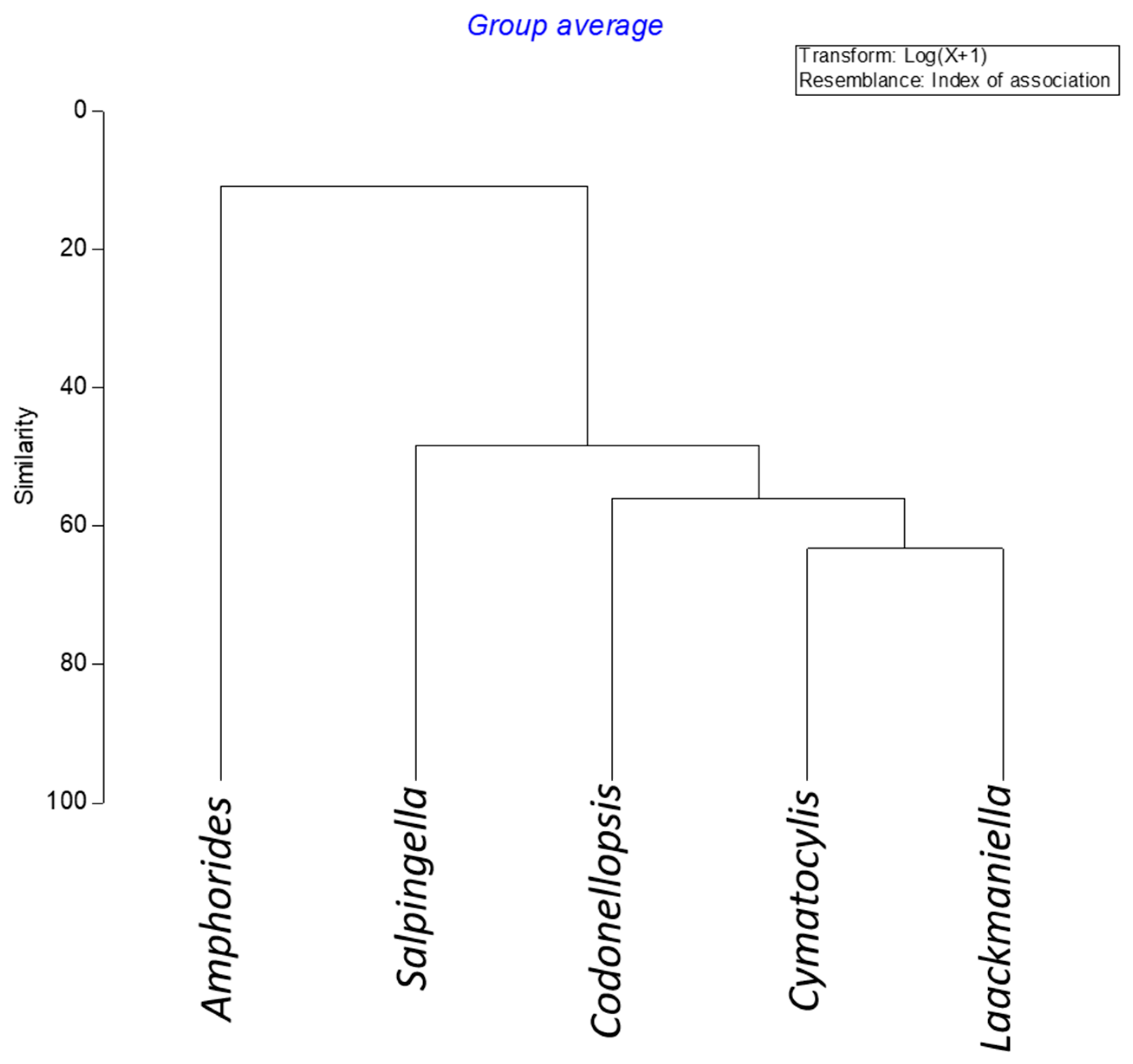
The cluster analysis, performed on the index of the association matrix among genera (
Figure 7), reveals the association between
Cymatocylis and
Laackmanniella. These genera followed the same trend both in the vertical and horizontal distribution, highlighting similarity in the abundance covariance.
Amphorides genus appeared less frequent and rare than the other genera considered in this study and was never detected at the surface layer or in the offshore stations.
3.4. Plurennial Pattern of the Tintinnids Community
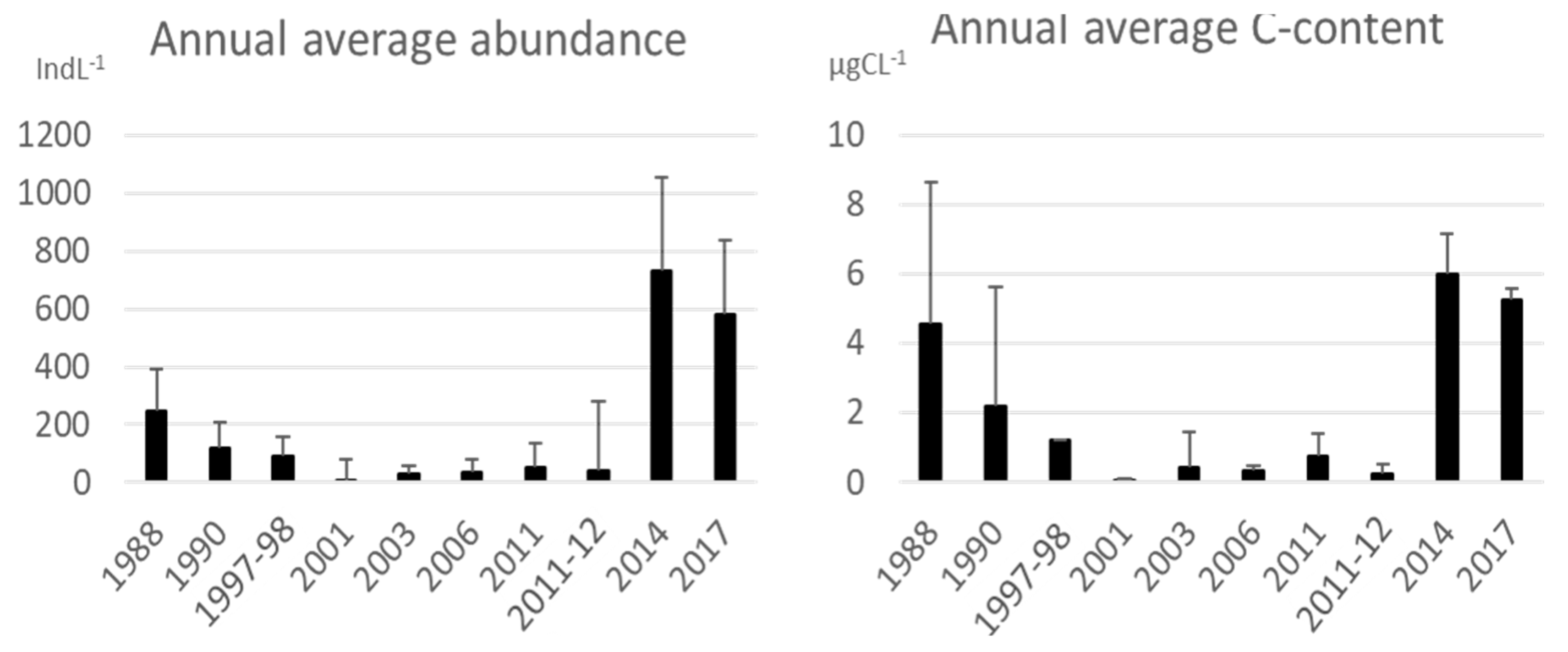
Average abundances recorded during years are summarized in
Figure 8: in 1988 the abundances were 249 ± 140 indL
−1 and then they decreased to 118 ± 90 indL
−1 in 1990, these two years with moderate abundances were followed by 5 years with very low abundances (never > 50 ± 87 indL
−1) which, however, returned to growth reaching the maximum peaks in 2014 (728 ± 325 indL
−1) and 2017 (582 ± 254 indL
−1). In general, the carbon content follows the same pattern as the abundance with small discrepancies related to the abundance of genera with larger or smaller dimensions in the tintinnids community.
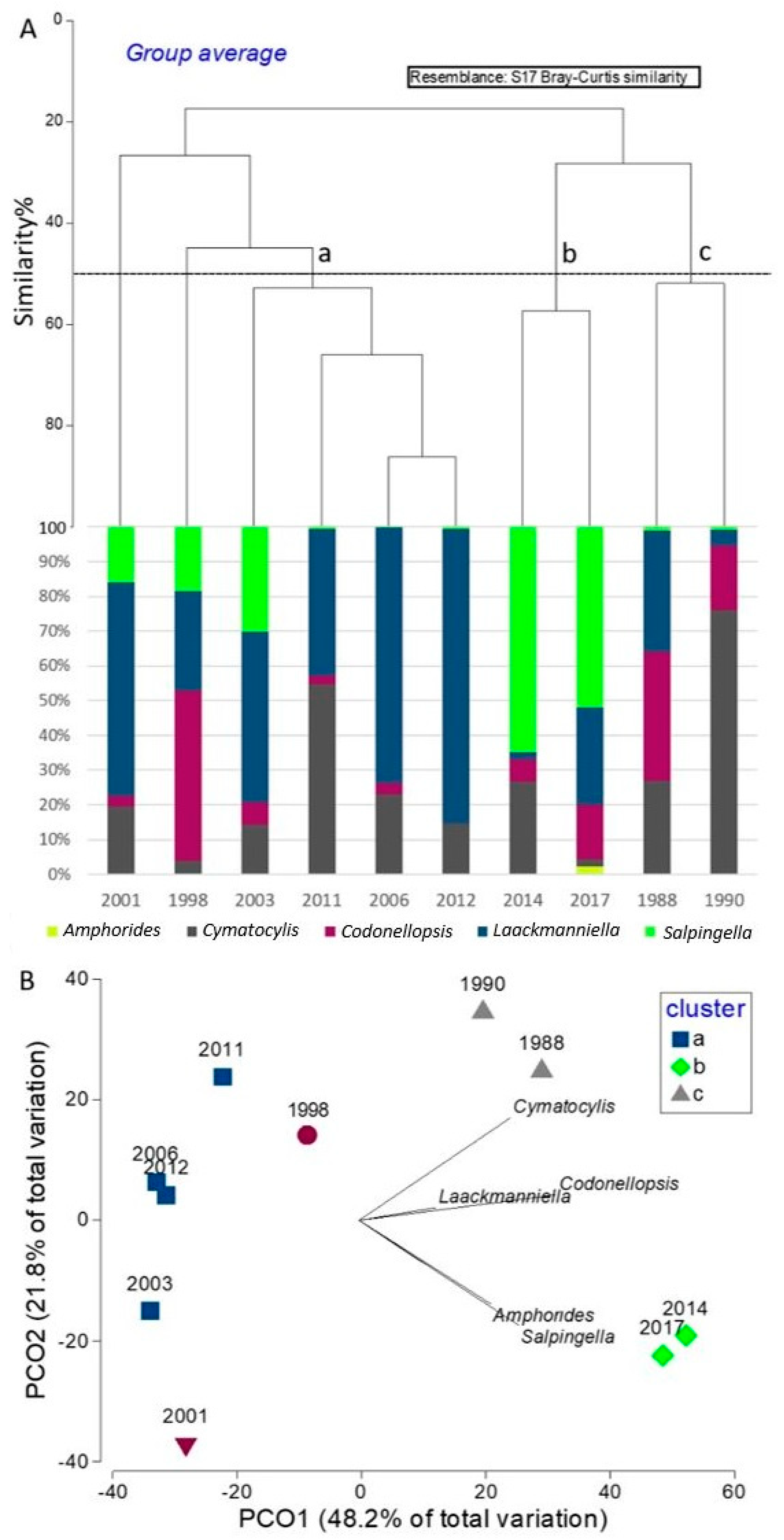
With reference to the diverse composition recorded in the different years (except 1994, and December samples of the campaigns 1997/98 and 2011/12), the cluster analysis (
Figure 9A) and PCO plot (
Figure 9B) were performed on the compositions of the populations in the different years and identified three significant groups (PERMANOVA
p < 0.05) over the slice of 50% in similarity: in the cluster A, four years (2003, 2006, 2011, 2012) were characterized in the simper analysis output (
Table 6) by highest relative abundances of the genera
Laackmaniella and
Cymatocylis. In cluster B, the years 2014 and 2017 were characterized by the highest abundances and a high percentage of genera
Salpingella and
Codonellopsis within the population; finally, cluster C (1988 and 1990), was characterized by intermediate abundances with high percentages of
Cymatocylis and
Codonellopsis. The years 1998 (showing the highest percentage in
Codonellopsis) and 2001 (showing the lowest total abundances) were not grouped in any cluster.
4. Discussion
Over eleven summer expeditions from 1988 to 2017 we have detected only nine tintinnids species belonging to five genera, therefore confirming the low tintinnids diversity in TNB winter polynya area, and in the Antarctic waters in general [
41,
54,
55,
56,
57]. Previously, in TNB a richer list comprising 14 species was reported [
58]. This difference is mainly due to a recent taxonomic rearrangement [
50]. Indeed,
Cymatocylis ecaudata Kofoid and Campbell 1929 detected in [
54] is now a synonym of
C. drygalskii and
C. folliculus Kofoid and Campbell 1929 as well as of
C. nobilis.
Cymatocylis conica (Laackmann) Kofoid and Campbell 1929 and
C. flava Laackmann 1910 are grouped into the species
C. vanhoeffeni, while
C. glans Kofoid and Campbell 1929 and
C. subconica Kofoid and Campbell 1929 are grouped into the species
C. cristallina.
Laackmanniella prolongata (Laackmann) Kofoid and Campbell 1929 is now a synonym of
L. naviculaefera [
50].
Excluding 1994, which had only one station, on average three species (
C. drygalskii,
C. gaussi, and
L. naviculaefera) were recorded for each year of the study period. These species are common in the Southern Ocean [
59] and they are often dominant in coastal tintinnid communities [
26]. In TNB polynya, they can be considered key species and their presence and fluctuation in abundance should be considered an important signal of possible changes in the whole plankton community.
Only in 1997/98 and 2011/12, we sampled in December, January, and February. On these two occasions, we recorded fewer species and lower abundances in early December compared to the January–February period, confirming a seasonal pattern and late development of the tintinnid community observed [
40,
60]. In December 1997 and 2011, tintinnids’ community was particularly scarce, whereas a significant increase over the period January–February was recorded.
The phytoplankton community in the Ross Sea is dominated by the haptophytes
Phaeocystis antarctica and diatoms, and it shows different temporal and spatial patterns [
18]. In TNB, the other functional groups (dinoflagellates, cryptophytes, cyanophytes, chlorophytes) are poorly represented and are considered to have a minor role in the Antarctic food web [
18,
20,
61,
62]. Generally, in the TNB during December there is a
Phaeocystis-dominated inshore area and a small-diatoms-dominated community at the retreating ice edge. Later in the season, the herbivorous food web is more active, whereas in late February the consumers’ community shifts toward smaller-sized organisms (namely protists), and the food web is considered mistivourus [
60]. Considering this as the usual seasonal food web structure development in the study area [
55,
63,
64], we decided to omit December samples from the multiyear comparisons (1994, 1997, and 2011).
A clear decreasing tintinnids abundance till 2012 is evident when considering the January–February annual average, which started during the first sampled year (1988) and dramatically dropped in January 2001. The general decreasing trend (despite the limitation due to the data collected over a long period and in different sites) might be due to the analogous decreasing trend of phytoplankton biomass, which was detected from 1997 onward [
11,
65]. In many studies, it has been demonstrated that microzooplankton (and consequently tintinnids) abundances are tightly coupled with chlorophyll concentration [
55,
57,
58,
66,
67,
68,
69]. Arrigo and van Dijken [
11] suggested that chlorophyll concentrations are mainly controlled by ice coverage. Low chlorophyll concentrations are measured in areas with less extension of ice-free waters. Consequently, we can assume that the increasing ice coverage since 1978 reported by Stammerjohn and Smith [
69] could have affected the phytoplankton biomass, which in turn controls micrograzers (namely protists) abundances. Particularly in the 1997–1998 and 2000–2001 austral summers, chlorophyll
a concentration was low over the whole season, and the phytoplankton blooms were less extended and delayed by almost two months, peaking only in February [
11]. In addition to the interannual fluctuation, which is probably related to ENSO (El Niño-Southern Oscillations) oscillations [
11], the 2000–2001 austral summer was impacted by the drifting of the immense iceberg B 15, which impeded the normal circulation pattern in the western Ross Sea, thus strongly affecting primary production and higher trophic levels through a cascading effect [
69]. In the austral summer 2002–2003 Harangozo and Connolley [
70] registered the minimum extent of open water in the Ross Sea, which again could have lowered primary production and consequently phytoplankton biomass. Moreover, minimum values of tintinnids biomass were detected in 2001 and they remained very low until 2014.
On the contrary, the increase in abundance in 2014 and 2017 can be related to the increase in the surface layer temperature, which may cause an increase in the buoyancy of the surface waters relative to the underlying layer and consequently decrease the vertical exchange.
The timing and magnitude of primary productivity in the Ross Sea has been changing during the last years, with a general increase in the area of the Ross Sea polynya [
71,
72]. These changes may have influenced the tintinnid community. In the last period (2014 and 2017) we noticed an increase in the abundance of the dominant species (
C. drygalskii,
C. gaussi, and
L. naviculaefera), and of the
Salpingella genus. Moreover,
A. laackmanni was detected for the first time in the area.
Salpingella and
A. laackmanni are two taxa not peculiar to Antarctica but widespread in the Southern Ocean [
41,
59] and they both present a similar oral diameter (LOD). The variation in the LOD of tintinnids can reflect the size spectrum of food items available. Previous studies showed that the species assemblage of the Southern Ocean appears to be distributed bimodally with peaks at about 45 and 120 µm, suggesting that most species likely use prey of 10–15 µm or 30 µm [
59]. The most abundant species detected in this study (
C. drygalskii,
C. gaussi, and
L. naviculaefera) fall perfectly into these two classes and the recent increase in small diatoms in TNB [
20] could explain their increase in abundance. On the contrary, the
Salpingella genus and
A. laackmanni present both a smaller oral diameter (LOD between 15 to 20 µm), and they are supposed to feed on prey < 10 µm. The TNB community size structure in 2017, together with the high percentage of microphytoplankton (39%), showed high values also for nano-phytoplankton (39%) and pico-phytoplankton (22%) [
73] as possible prey for tintinnids with smaller LOD.
The different abundances of tintinnids in TNB can be linked to the higher biomass of diatoms reported near the coastline [
20,
26,
73]. Previous studies showed
L. naviculaefera and
C. gaussi more abundant in the stations close to the coast and
Cymatocylis in the offshore stations [
58]. Additionally, our results showed that
L. naviculaefera was more abundant in the stations closer to the coast, while other species showed different tendencies. We speculate that these differences are due to the small sampling scale employed in our study, which means that in general our samples can be compared to the coastal area samples in other studies, (e.g., [
73]).
During the summer the area presents a typical vertical structure, with the DSW at the bottom and the lightest Antarctic surface water (AASW) occupying the top 100 m layer [
73]. The higher abundance of tintinnids in the layer 0–100 m is confirmed also by previous studies where
Salpingella,
C. gaussi, and
L. naviculaefera were present in higher abundance at the surface, while
C. drygalskii and
C. convallaria were mainly found at intermediate depths [
42,
74]. Liang et al. [
42] showed that
A. laackmanni had no clear depth pattern, notwithstanding in our study this species was mainly found in the 10–100 m layer.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}