
The Potential Effect of Climate Change on the Distribution of Endemic Anurans from Mexico’s Tropical Dry Forest

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
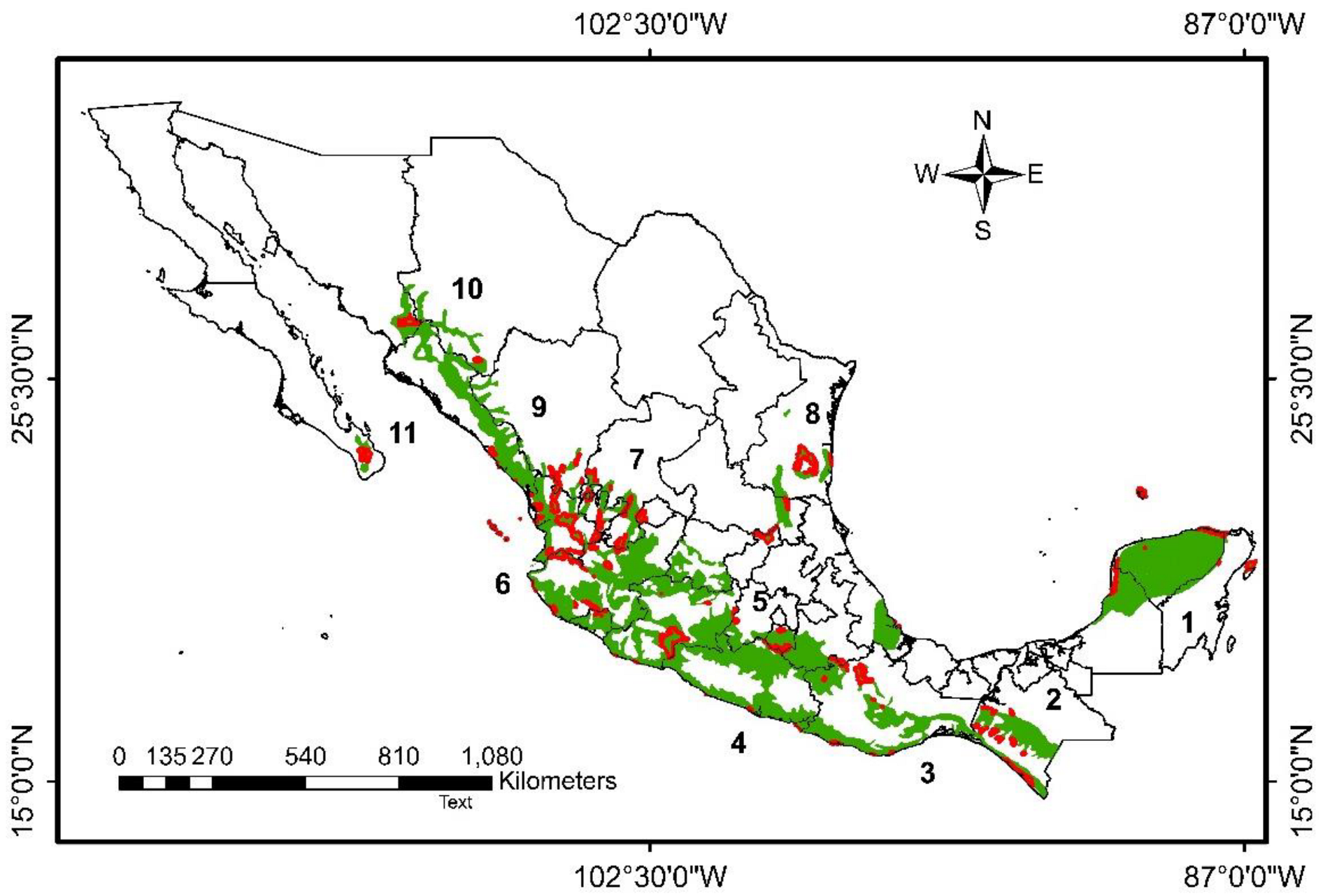
2.1. Study Area
2.2. Species Records and Occurrence Data
2.3. Environmental Data and Determination of the Accessible Area (M)
2.4. Potential Distribution Modeling
3. Results
3.1. Species and Model Performance
3.2. Current and Future Suitable Habitat
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Pörtner, H.O.; Scholes, R.J.; Agard, J.; Archer, E.; Arneth, A.; Bai, X.; Barnes, D.; Burrows, M.; Chan, L.; Cheung, W.L.; et al. IPBES-IPCC Co-Sponsored Workshop Report on Biodiversity and Climate Change. IPBES-IPCC Co-Sponsored Workshop Report on Biodiversity and Climate Change, 1st ed.; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2021; pp. 15–17. [Google Scholar] [CrossRef]
- Hansen, A.; Neilson, R.P.; Dale, V.H.; Flather, C.H.; Iverson, L.R.; Currie, D.J.; Shafer, S.; Cook, R.; Bartlein, P.J. Global changes in forest: Responses of species, communities and biomes. BioScience 2001, 51, 765–779. [Google Scholar] [CrossRef]
- Parmesan, C. Climate and species’ range. Nature 1996, 382, 765–766. [Google Scholar] [CrossRef]
- Parmesan, C.; Nils, R.; Stefanescus, C.; Hill, J.K.; Thomas, D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Polewards shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Biggs, R.; Simons, H.; Bakkenes, M.; Scholesa, R.J.; Eickhout, B.; Van Vuuren, D.; Alkemade, R. Scenarios of biodiversity loss in southern Africa in the 21st century. Glob. Environ. Chang. 2008, 18, 296–309. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and Status and trends of amphibian declines and extinction worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 15, 11466–11473. Available online: www.pnas.org/cgi/doi10.1073/pnas.0801921105 (accessed on 20 February 2022). [CrossRef] [PubMed]
- Abarca-Alvarado, J.G. Endangered amphibians: Threats and effective conservation strategies. Biocenosis 2021, 32, 133–145. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Raven, P.H. Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 13596–13602. Available online: https://www.pnas.org/doi/full/10.1073/pnas.1922686117 (accessed on 10 February 2022). [CrossRef] [PubMed]
- Araújo, M.B.; Thuiller, W.; Pearson, R.G. Climate warming and the decline of amphibians and reptiles in Europe. J. Biogeogr. 2006, 33, 1712–1728. [Google Scholar] [CrossRef]
- García, A.; Ortega-Huerta, M.A.; Martínez-Meyer, E. Potential distributional changes and conservation priorities of endemic amphibians in western Mexico as a result of climate change. Environ. Conserv. 2013, 47, 1–12. [Google Scholar] [CrossRef]
- Ochoa-Ochoa, L.; Rodríguez, P.; Mora, F.; Flores-Villela, O.; Whittaker, R. Climate change and amphibian diversity patterns in Mexico. Biol. Conserv. 2012, 150, 94–102. [Google Scholar] [CrossRef]
- World Wildlife Fundation (WWF). Impactos y Vulnerabilidad al Cambio Climático en México [en línea]. 2010. Available online: http://d2ouvy59p0dg6k.cloudfront.net/downloads/03_impactos_nacionales_e_internacionales_del_cambio_climatico.pdf (accessed on 5 March 2022).
- Wake, D.B.; Koo, M.S. Primer: Amphibians. Curr. Biol. 2018, 28, R1237–R1241. [Google Scholar] [CrossRef] [PubMed]
- Balderas-Valdivia, C.J.; González-Hernández, A. Inventario de la herpetofauna de México 2021. Herpetol. Mex. 2021, 2, 10–71. [Google Scholar]
- Parra-Olea, G.; Flores-Villela, O.; Mendoza-Almeralla, C. Biodiversidad de anfibios en México. Rev. Mex. Biodivers. Supl. 2014, 85, S460–S466. [Google Scholar] [CrossRef]
- Stuart, S.N.; Hoffmann, M.; Chanson, J.S.; Cox, N.A.; Berridge, R.J.; Ramani, P.; Young, B.E. Threatened Amphibians of the World, 1st ed.; Lynx Edicions: Barcelona, Spain, 2008; pp. 2–3. [Google Scholar]
- Balvanera, P.; Mass, M. Los Servicios Ecosistémicos Que Proveen las Selvas Secas. In Diversidad, Amenazas y Áreas Prioritarias Para la Conservación de las Selvas Secas del Pacífico de México, 1st ed.; Ceballos, G., Martínez, L., García, A., Espinoza, E., Bezaury Creel, J., Dirzo, R., Eds.; Fondo De Cultura Económica (FCE): Mexico City, México; Comisión Nacional para el Fomento y Uso de la Biodiversidad (CONABIO): Mexico City, México; Comisión Nacional de Áreas Naturales Protegidas (CONANP): Mexico City, México, 2010; pp. 251–270. ISBN 970-9000-38-1. [Google Scholar]
- Jaramillo, V.; García-Oliva, F.; Martínez-Yrízar, A. La selva seca y las perturbaciones antrópicas en un contexto funcional. In Diversidad, Amenazas y Áreas Prioritarias Para la Conservación de Las Sevas Secas del Pacífico de México, 1st ed.; Ceballos, G., Martínez, L., García, A., Espinoza, E., Bezaury Creel, J., Dirzo, R., Eds.; Fondo De Cultura Económica (FCE): Mexico City, México; Comisión Nacional para el Fomento y Uso de la Biodiversidad (CONABIO): Mexico City, México; Comisión Nacional de Áreas Naturales Protegidas (CONANP): Mexico City, México, 2010; pp. 235–250. ISBN 970-9000-38-1. [Google Scholar]
- Banda, R.K.; Delgado-Salinas, A.; Dexter, K.G.; Linares-Palomino, R.; Oliveira-Filho, A.; Prado, D.; Pullan, M.; Quintana, C.; Riina, R.; Rodriguez, M.G.M.; et al. Plant diversity patterns in neotropical dry forests and their conservation implications. Science 2016, 6306, 1383–1387. [Google Scholar] [CrossRef]
- Ceballos, G. Vertebrate diversity, ecology, and conservation in Neotropical dry forests. In Seasonally dry Tropical Forests, 1st ed.; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 195–220. [Google Scholar]
- Trejo, I. Las selvas secas del Pacífico mexicano. In Diversidad, Amenazas y Áreas Prioritarias Para la Conservación de las Selvas Secas del Pacífico de México, 1st ed.; Ceballos, G., Martínez, L., García, A., Espinoza, E., Bezaury Creel, J., Dirzo, R., Eds.; Fondo De Cultura Económica (FCE): Mexico City, México; Comisión Nacional para el Fomento y Uso de la Biodiversidad (CONABIO): Mexico City, México; Comisión Nacional de Áreas Naturales Protegidas (CONANP): Mexico City, México, 2010; pp. 41–52. ISBN 970-9000-38-1. [Google Scholar]
- Trejo-Vázquez, I. El clima de la selva baja caducifolia en México. Investig. Geográficas 1999, 39, 40–52. [Google Scholar] [CrossRef]
- Ceballos, G.; Valenzuela, D. Diversidad, ecología y conservación de los vertebrados de Latinoamérica. In Diversidad, Amenazas y Áreas Prioritarias Para la Conservación de las Selvas Secas del Pacífico de, 1st ed.; Ceballos, G., Martínez, L., García, A., Espinoza, E., Bezaury Creel, J., Dirzo, R., Eds.; Fondo De Cultura Económica (FCE): Mexico City, México; Comisión Nacional para el Fomento y Uso de la Biodiversidad (CONABIO): Mexico City, México; Comisión Nacional de Áreas Naturales Protegidas (CONANP): Mexico City, México, 2010; pp. 94–118. ISBN 970-9000-38-1. [Google Scholar]
- Huechacona-Ruiz, A.H.; Dupuy, J.M.; Schwartz, N.B.; Powers, J.S.; Reyes-García, C.; Tun-Dzul, F.; Hernández-Stefanoni, J.L. Mapping Tree Species Deciduousness of Tropical Dry Forests Combining Reflectance, Spectral Unmixing, and Texture Data from High-Resolution Imagery. Forest 2020, 11, 1234. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Benavides, E.; Breceda, A.; Anadón, J.D. Winners and losers in the predicted impact of climate change on cacti species in Baja California. Plant Ecol. 2021, 222, 29–44. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.A. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2015, 275, 73–77. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Griffies, S.M.; Winton, M.; Donner, L.J.; Horowitz, L.W.; Downes, S.M.; Farneti, R.; Gnanadesikan, A.; Hurlin, W.J.; Lee, H.; Liang, Z.; et al. The GFDL CM3 coupled climate model: Characteristics of the ocean and sea ice simulations. J. Clim. 2011, 24, 3520–3544. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetación Potencial. IV.8.2. Atlas Nacional de México. Vol II. Escala 1:4000000. Instituto de Geografía, UNAM. México. 1990. Available online: http://www.conabio.gob.mx/informacion/gis/ (accessed on 27 July 2021).
- Soberón, J.; Peterson, A.T. Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef]
- Guevara, L.; Gerstner, B.E.; Kass, J.M.; Anderson, R.P. Toward ecologically realistic predictions of species distributions: A cross-time example from tropical montane cloud forests. Glob. Chang. Biol. 2018, 24, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Wiensa, J.A.; Stralberga, D.; Jongsomjita, D.; Howella, C.A.; Snyderb, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Zimmermann, N.E. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Pérez-García, B.; Liria, J. Modelos de nicho ecológico fundamental para especies del género Thraulodes (Ephemeroptera: Leptophlebiidae: Atalophlebiinae). Rev. Mex. Biodivers. 2013, 84, 600–611. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Meza-Joya, F.L.; Rojas-Morales, J.A.; Ramos, E. Predicting distributions of rare species: The case of the false coral snake Rhinobothryum bovallii (Serpentes: Colubridae). Phyllomedusa 2020, 19, 141–164. [Google Scholar] [CrossRef]
- Berriozabal-Islas, C.; Mota Rodrigues, J.F.; Ramírez-Bautista, A.; Becerra-López, J.L.; Nieto-Montes de Oca, A. Effect of climate change in lizards of the genus Xenosaurus (Xenosauridae) based on projected changes in climatic suitability and climatic niche conservatism. Ecol. Evol. 2018, 8, 6860–6871. [Google Scholar] [CrossRef] [PubMed]
- Correia, L.O.; Siqueira Júnior, S.; Carneiro, P.L.S.; Bezerra, M.A. Evaluation of the use of Leptodactylus ocellatus (Anura: Leptodactylidae) frog tissues as bioindicator of metal contamination in Contas River, Northeastern Brazil. An. Acad. Bras. Ciênc 2014, 86, 1549–1561. [Google Scholar] [CrossRef]
- García, A. Reptiles y anfibios. In Diversidad, Amenazas y Áreas Prioritarias Para la Conservación de las Selvas Secas del Pacífico de México, 1st ed.; Ceballos, G., Martínez, L., García, A., Espinoza, E., Bezaury Creel, J., Dirzo, R., Eds.; Fondo De Cultura Económica (FCE): Mexico City, México; Comisión Nacional para el Fomento y Uso de la Biodiversidad (CONABIO): Mexico City, México; Comisión Nacional de Áreas Naturales Protegidas (CONANP): Mexico City, México, 2010; pp. 165–178. ISBN 970-9000-38-1. [Google Scholar]
- Prieto-Torres, D.A.; Navarro-Sigüenza, A.G.; Santiago-Alarcón, D.; Rojas-Soto, O.R. Response of the endangered tropical dry forests to climate change and the role of Mexican Protected Areas for their conservation. Glob. Chang. Biol. 2016, 22, 364–379. [Google Scholar] [CrossRef]
- Pounds, J.A.; Fogden, M.P.L.; Campbell, J.H. Biological response to climate change on a tropical mountain. Nature 1999, 398, 611–615. [Google Scholar] [CrossRef]
- IPCC (Intergovernmental Panel on Climate Change). Climate Change and Biodiversity. IPCC Technical Paper V. IPCC-WMO-UNEP. 2002. Available online: https://www.tnrf.org/files/E-INFO_IPCC_2002_Climate_Change_and_Biodiversity_0.pdf (accessed on 28 July 2021).
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef]
- Li, Y.; Cohen, J.M.; Rohr, J.R. Review and synthesis of the effects of climate change on amphibians. Integr. Zool. 2013, 8, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; Madison, D.M. Dryness increase predation risk in efts: Support for an amphibian decline hypothesis. Oecologia 2003, 135, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Theurillat, J.P. Monitoring Networks for Testing Model-Based Scenarios of Climate Change Impact on Mountain Plant Distribution. In Global Change and Mountain Regions. Advances in Global Change Research; Huber, U.M., Bugmann, H.K.M., Reasoner, M.A., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 467–476. [Google Scholar]
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species-climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Graham, C.H. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Chang. Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Bioclimatic Variables |
|---|---|
| Bio 1 | Annual mean temperature (°C) |
| Bio 2 | Mean diurnal range (mean of monthly (max temp–min temp)) (°C) |
| Bio 3 | Isothermality ((BIO2/BIO7) × 100) (°C) |
| Bio 4 | Temperature seasonality (standard deviation × 100) (°C) |
| Bio 5 | Maximum temperature of warmest month (°C) |
| Bio 6 | Minimum temperature of coldest month (°C) |
| Bio 7 | Temperature Annual Range (BIO5-BIO6) |
| Bio 8 | Mean temperature of wettest quarter (°C) |
| Bio 9 | Mean temperature of driest quarter (°C) |
| Bio 10 | Mean temperature of warmest quarter (°C) |
| Bio 11 | Mean temperature of coldest quarter (°C) |
| Bio 12 | Annual precipitation (mm) |
| Bio 13 | Precipitation of wettest month (mm) |
| Bio 14 | Precipitation of driest month (mm) |
| Bio 15 | Precipitation seasonality (standard deviation × 100) |
| Bio 16 | Precipitation of wettest quarter (mm) |
| Bio 17 | Precipitation of driest quarter (mm) |
| Bio 18 | Precipitation of warmest quarter (mm) |
| Bio 19 | Precipitation of coldest quarter (mm) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballesteros-Barrera, C.; Tapia-Pérez, O.; Zárate-Hernández, R.; Leyte-Manrique, A.; Martínez-Bernal, A.; Vargas-Miranda, B.; Martínez-Coronel, M.; Ortiz-Burgos, S. The Potential Effect of Climate Change on the Distribution of Endemic Anurans from Mexico’s Tropical Dry Forest. Diversity 2022, 14, 650. https://doi.org/10.3390/d14080650
Ballesteros-Barrera C, Tapia-Pérez O, Zárate-Hernández R, Leyte-Manrique A, Martínez-Bernal A, Vargas-Miranda B, Martínez-Coronel M, Ortiz-Burgos S. The Potential Effect of Climate Change on the Distribution of Endemic Anurans from Mexico’s Tropical Dry Forest. Diversity. 2022; 14(8):650. https://doi.org/10.3390/d14080650
Chicago/Turabian StyleBallesteros-Barrera, Claudia, Oscar Tapia-Pérez, Rocío Zárate-Hernández, Adrián Leyte-Manrique, Angélica Martínez-Bernal, Bárbara Vargas-Miranda, Matías Martínez-Coronel, and Selene Ortiz-Burgos. 2022. "The Potential Effect of Climate Change on the Distribution of Endemic Anurans from Mexico’s Tropical Dry Forest" Diversity 14, no. 8: 650. https://doi.org/10.3390/d14080650