
Deep Troglomorphy: New Arrhopalitidae (Collembola: Symphypleona) of Different Life Forms from the Snezhnaya Cave System in the Caucasus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Locality
2.2. Sampling
2.3. Microscopic Study
2.4. Measurements
2.5. Chaetotaxy Nomenclature
2.6. Abbreviations
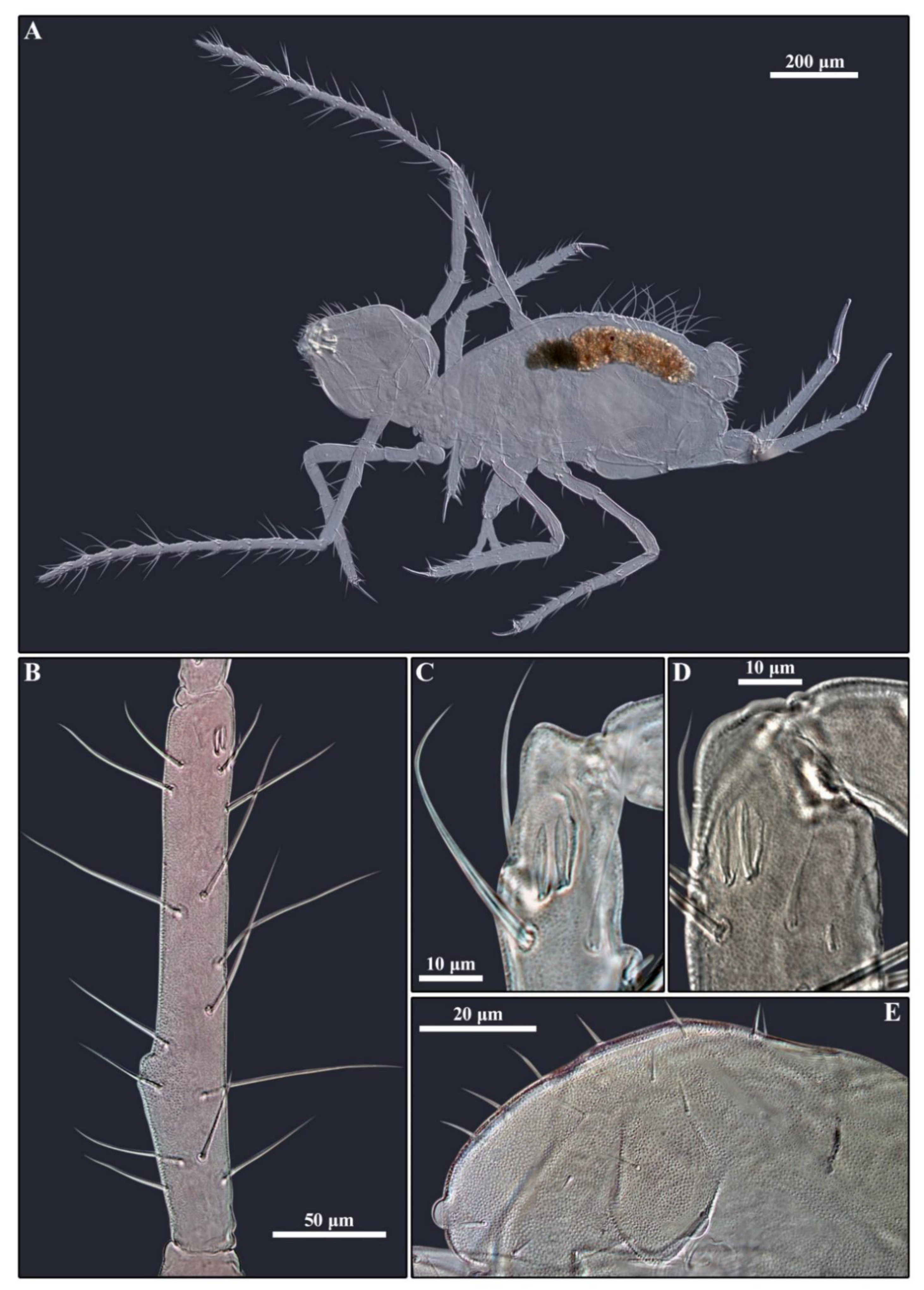
3. Results
3.1. Genus Arrhopalites Börner, 1906
3.2. Genus Pygmarrhopalites Vargovitsh, 2009
4. Discussion
4.1. Reasons of Different Troglomorphy Level in Arrhopalitidae
4.1.1. Ecological Evolutionary Trajectory of Certain Lineages
4.1.2. Evolutionary Age of Troglobiont Species
4.1.3. Habitat Preferences of Species in Subterranean Realm
4.2. Troglomorphic Features in Arrhopalitidae
4.2.1. Elongated Antennae with Multi-Subdivided Fourth Antennal Segment
4.2.2. Elongated Legs
4.2.3. Elongated and Slender Claws
4.2.4. Elongated Furca and Modified Lamellae of Mucro
4.2.5. Enlarged Sensory Organ of the Third Antennal Segment
4.2.6. Thinning of the Spine-like Setae on Head and Dens
4.2.7. Elongated Setae of Posterior Dorsal Complex
4.3. Life Forms in Cave-Dwelling Arrhopalitidae
4.3.1. Neustonic-Hygropetric Troglomorphs (Cave Water & Wet-Stone Walkers)
4.3.2. Atmobiont Troglomorphs (Cave Terrestrial Walkers)
4.3.3. Intermediate Troglomorphs (Cave Terrestrial & Water Walkers)
4.3.4. Intrasubstrate Troglomorphs (Cave-Dwelling within-Substrate Inhabitants)
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antić, D.Ž.; Makarov, S.E. The Caucasus as a major hotspot of biodiversity: Evidence from the millipede family Anthroleucosomatidae (Diplopoda, Chordeumatida). Zootaxa 2016, 4211, 1–205. [Google Scholar] [CrossRef] [PubMed]
- Fiera, C.; Arbea, J.I.; Vargovitsh, R.S.; Barjadze, S. A synthesis on troglobitic springtails in Europe. J. Zool. Syst. Evol. Res. 2021, 59, 1874–1890. [Google Scholar] [CrossRef]
- Turbanov, I.S.; Palatov, D.M.; Golovatch, S.I. The state of the art of biospeleology in Russia and other countries of the former Soviet Union: A review of the cave (endogean) invertebrate fauna. 2. Arachnida—Acknowledgments. Entomol. Rev. 2016, 96, 1297–1333. [Google Scholar] [CrossRef]
- Jordana, R.; Baquero, E.; Reboleira, S.; Sendra, A. Reviews of the genera Schaefferia Absolon, 1900, Deuteraphorura Absolon, 1901, Plutomurus Yosii, 1956 and the Anurida Laboulbène, 1865 species group without eyes, with the description of four new species of cave springtails (Collembola) from Krubera-Voronya cave, Arabika Massif, Abkhazia. Terr. Arthropod Rev. 2012, 5, 35–85. [Google Scholar] [CrossRef]
- Babenko, A.B. New species of springtails of the genus Typhlogastrura (Collembola, Hypogastruridae) from caves of the USSR. Zool. Zh. 1987, 66, 463–472. (In Russian) [Google Scholar]
- Vargovitsh, R.S. Cave water walker: An extremely troglomorphic Troglaphorura gladiator gen. et sp. nov. (Collembola, Onychiuridae) from Snezhnaya Cave in the Caucasus. Zootaxa 2019, 4619, 267–284. [Google Scholar] [CrossRef]
- Vargovitsh, R.S. Cavernicolous Arrhopalites abchasicus sp. nov. (Collembola: Symphypleona: Arrhopalitidae) from the West Caucasus with a key to the World species of the genus. Zootaxa 2013, 3666, 16–30. [Google Scholar] [CrossRef]
- Vargovitsh, R.S. Two new troglobiont Pygmarrhopalites species of the principalis group (Collembola: Arrhopalitidae) from the West Caucasus. Zootaxa 2017, 4250, 23–42. [Google Scholar] [CrossRef]
- Vargovitsh, R.S. New troglomorphic Arrhopalitidae (Collembola: Symphypleona) from the Western Caucasus. Zootaxa 2012, 3174, 1–21. [Google Scholar] [CrossRef]
- Barjadze, S.; Djanashvili, R. Checklist of the springtails (Collembola) of Georgia. Caucasian Entomol. Bull. 2008, 4, 187–193. [Google Scholar] [CrossRef]
- Klimchouk, A. Caucasus, Georgia. In Encyclopedia of Caves and Karst Science; Gunn, J., Ed.; Fitzroy Dearborn: Chicago, IL, USA, 2004; pp. 412–417. [Google Scholar]
- Vakhrushev, B.A.; Dublyansky, V.N.; Amelichev, G.N. Karst of the Bzybsky Ridge. The Western Caucasus; Peoples’ Friendship University of Russia: Moscow, Russia, 2001; pp. 1–165. (In Russian) [Google Scholar]
- Gusev, A.S. Hydrology of underground water of Khipsta Massif (Abkhazia). Probl. Geogr. 2018, 147, 107–133. (In Russian) [Google Scholar]
- Mavlyudov, B.R.; Shelepin, A.L. Systema Snezhnaya. In Atlas of Caves of Russia; Shelepin, A.L., Ed.; Russian Geographical Society: Moscow, Russia, 2019; pp. 751–762. (In Russian) [Google Scholar]
- Mavlyudov, B.R. The Snezhnaya-Mezhennogo-Illyuziya cave system in the western Caucasus. Bol. Geol. Min. 2016, 127, 219–235. [Google Scholar]
- Barber, H.S. Traps for cave-inhabiting insects. J. Elisha Mitchell Sci. Soc. 1931, 46, 259–267. [Google Scholar]
- Mercet, R.G. Los enemigos de los parásitos de las plantas. Trab. Mus. Nac. Ci. Nat. 1912, 6, 1–306. [Google Scholar]
- Betsch, J.-M.; Waller, A. Chaetotaxic nomenclature of the head, thorax and abdomen in Symphypleona (Insecta, Collembola). Acta Zool. Fenn. 1994, 195, 5–12. [Google Scholar]
- Fjellberg, A. The labial palp in Collembola. Zool. Anz. 1999, 237, 309–330. [Google Scholar]
- Vargovitsh, R.S. New cave Arrhopalitidae (Collembola: Symphypleona) from the Crimea (Ukraine). Zootaxa 2009, 2047, 1–47. [Google Scholar] [CrossRef]
- Bretfeld, G. The chaetotaxy of the small abdomen of the Symphypleona (Insecta, Collembola) and its phylogenetic interpretation. Acta Zool. Fenn. 1994, 195, 13–17. [Google Scholar]
- Betsch, J.-M. An ontogenetically focused chaetotaxical scheme in Symphypleona (Collembola): The 6th abdominal segment. Pedobiologia 1997, 41, 13–18. [Google Scholar]
- Nayrolles, P. Chétotaxie tibiotarsale des Collemboles Symphypléones. Trav. Lab. Ecobiol. Arthrop. Edaph. Toulouse 1988, 5, 1–19. [Google Scholar]
- Nayrolles, P. Chétotaxie furcale des Collemboles Symphypléones. Trav. Lab. Ecobiol. Arthrop. Edaph. Toulouse 1990, 6, 27–50. [Google Scholar]
- Nayrolles, P. La chétotaxie antennaire des Collemboles Symphypléones. Trav. Lab. Ecobiol. Arthrop. Edaph. Toulouse 1991, 6, 1–94. [Google Scholar]
- Yosii, R. Results of the speleological survey in South Korea 1966. IV. Cave Collembola of South Korea. Bull. Natl. Sci. Mus. Tokyo 1966, 9, 541–561. [Google Scholar]
- Bretfeld, G. Synopses on Palaearctic Collembola: Symphypleona; Staatliches Museum für Naturkunde Görlitz: Görlitz, Germany, 1999; Volume 2, pp. 1–318. [Google Scholar]
- Mazina, S.E.; Bazarova, E.P.; Bashirov, A.M.; Gabbasova, E.R.; Gusev, A.S.; Kopachevskiy, Y.Y.; Kochetkov, S.N.; Savinov, V.P.; Shadrin, V.O. Abiotic Components of an Ecosystem of a Snezhnaya (Illyuzia-Mezhonnogo-Snezhnaya) Cave: A Condition and Problems of Research. In Mineralogy of Technogenesis–2011; Potapov, S.S., Ed.; IMin UrO RAS: Miass, Russia, 2011; pp. 219–238. (In Russian) [Google Scholar]
- Stach, J. The species of the genus Arrhopalites occurring in European caves. Act. Mus. Hist. Nat. Acad. Polon. Litt. Sci. 1945, 1, 1–47. [Google Scholar]
- Christiansen, K.; Bellinger, P. Cave Arrhopalites new to science. J. Cave Karst Stud. 1996, 58, 168–180. [Google Scholar]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World (1996–2022). Available online: http://www.collembola.org (accessed on 1 August 2022).
- Christiansen, K. Morphological Adaptations. In Encyclopedia of Caves; Culver, D.C., White, W., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2005; pp. 386–397. [Google Scholar]
- Vandel, A. Biospeleology. The Biology of Cavernicolous Animals; Pergamon Press: Oxford, UK, 1965; pp. 1–524. [Google Scholar]
- Kováč, Ľ.; Parimuchová, A.; Miklisová, D. Distributional patterns of cave Collembola (Hexapoda) in association with habitat conditions, geography and subterranean refugia in the Western Carpathians. Biol. J. Linn. Soc. 2016, 119, 571–592. [Google Scholar] [CrossRef]
- Vargovich, R.S. Arrhopalites kristiani sp. n. (Collembola: Symphypleona: Arrhopalitidae) from a cave in Eastern Carpathians. Vestn. Zool. 2005, 39, 67–73. [Google Scholar]
- Nosek, J. A new springtail from the caves of the Moravský kras (Moravian Karst) Arrhopalites ruseki sp. n. Speleol. Vĕstn. 1975, 6, 45–48. [Google Scholar]
- Loksa, I. Faunistisch-systematische und ökologische Untersuchungen in der Lóczy-Höhle bei Balatonfüred (Biospeologica Hungarica, XI). Ann. Univ. Sci. Bp. Rolando Eötvös Nomin. Sect. Biol. 1960, 3, 253–266. [Google Scholar]
- Vargovitsh, R.S. Arrhopalites potapovi sp. nov. (Collembola, Symphypleona) from Russia. Zootaxa 2015, 3955, 101–112. [Google Scholar] [CrossRef]
- Vargovitsh, R.S.; Buşmachiu, G. A new species and new records of Arrhopalitidae (Collembola: Symphypleona) from the Republic of Moldova. Zootaxa 2015, 3973, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Vargovitsh, R.S.; Kahrarian, M. A new species of Arrhopalites Börner, 1906 (Collembola, Symphypleona, Arrhopalitidae) from Iran with an updated key to A. diversus group of species. Zootaxa 2020, 4759, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Zeppelini, D.; Brito, R.A.; Lima, E.C.A. Three new species of Collembola (Arthropoda: Hexapoda) from Central Brazilian shallow caves: Side effects of long term application of environmental law on conservation. Zootaxa 2018, 4500, 59–81. [Google Scholar] [CrossRef] [PubMed]
- Nosek, J. Arrhopalites slovacicus a new species of Collembola from the Domica Cave. Rev. Suisse Zool. 1975, 82, 599–602. [Google Scholar] [CrossRef]
- Christiansen, K. The genus Arrhopalites in the United States and Canada. Int. J. Speleol. 1966, 2, 43–73. [Google Scholar] [CrossRef]
- Yosii, R. Monographie zur Höhlencollembolen Japans; Contributions from the Biological Laboratory Kyoto University: Kyoto, Japan, 1956; Volume 3, pp. 1–109. [Google Scholar]
- Christiansen, K. Convergence and parallelism in cave Entomobryinae. Evolution 1961, 15, 288–301. [Google Scholar] [CrossRef]
- Thibaud, J.-M.; Massoud, Z. Un nouveau genre d’insectes collemboles Hypogastruridae cavernicole du Pays Basque. Mém. Biospéol. 1983, 10, 317–319. [Google Scholar]
- Deharveng, L. Collemboles cavernicoles VIII. Contribution a l’etude des Oncopoduridae. Bull. Soc. Entomol. Fr. 1988, 92, 133–147. [Google Scholar] [CrossRef]
- Deharveng, L.; Thibaud, J.-M. Bessoniella procera n. g., n. sp., nouvel Orchesellidae cavernicole relictuel des Pyrénées (Insecta, Collembola). Bull. Mus. Natl. Hist. Nat. Sér. 4 1989, 11, 397–405. [Google Scholar]
- Lukić, M.; Houssin, C.; Deharveng, L. A new relictual and highly troglomorphic species of Tomoceridae (Collembola) from a deep Croatian cave. ZooKeys 2010, 69, 1–16. [Google Scholar] [CrossRef]
- Loksa, I. Vier neue Höhlencollembolen aus Ungarn (Biospeologica Hungarica, XXIII). Opusc. Zool. Bp. 1967, 6, 289–296. [Google Scholar]
- Howarth, F.G.; Moldovan, O.T. Where Cave Animals Live. In Cave Ecology; Ecological Studies; Moldovan, O., Kováč, Ľ., Halse, S., Eds.; Springer: Cham, Switzerland, 2018; Volume 235, pp. 23–37. [Google Scholar] [CrossRef]
- Christiansen, K. Proposition pour la classification des animaux cavernicoles. Spelunca 1962, 2, 76–78. [Google Scholar]
- Christiansen, K. Bionomics of Collembola. Annu. Rev. Entomol. 1964, 9, 147–178. [Google Scholar] [CrossRef]
- Sket, B. The cave hygropetric—A little known habitat and its inhabitants. Arch. Hydrobiol. 2004, 160, 413–425. [Google Scholar] [CrossRef]
- Vargovich, R.S. A new species of Collembola of the genus Arrhopalites (Entognatha, Hexapoda) from a cave in Eastern Carpathians. Vestn. Zool. 1999, 33, 89–92. [Google Scholar]
- Zeppelini, D. The genus Arrhopalites Börner, 1906 (Collembola, Appendiciphora, Arrhopalitidae) in the Neotropical Region, with description of four new cave species from Brazil. Zootaxa 2006, 1124, 1–40. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Part | Arrhopalites profundus | Pygmarrhopalites rystsovi | ||||
|---|---|---|---|---|---|---|
| Females, n = 2 + 1? | Male | Females, n = 6 | Male | |||
| Holotype | n = 2 | n = 1 | Min–Max | Mean | n = 1 | |
| Total (without appendages) | ~1000 | 875; ? | 920 | 1000–1150 | 1098 | ~950 |
| Body | ~700 | 620; ? | 640 | 730–830 | 792 | ~700 |
| Head | 387 | 250; ? | 305 | 340–385 | 353 | 302 |
| Head dorsum longest seta | 25 | 26; ? | 23 | 21–24 | 23 | 18 |
| Eye diameter | 5.5 | ? | 5.8 | 11.6–13.8 | 12.7 | 13 |
| Antenna | 755 | ? | 660 | 1160–1270 | 1197 | 1130 |
| Ant I | 53 | ? | 49 | 51–56 | 54 | 53 |
| Ant II | 110 | ? | 102 | 146–160 | 151 | 138 |
| Ant III | 182 | ? | 140 | 252–288 | 264 | 249 |
| Ant IV | 440 | ? | 395 | 705–812 | 748 | 714 |
| Ant III organ rods | 15 | 15.2; ? | 14.8 | 10–11.6 | 11 | 11.5 |
| Tibiotarsus I | 257 | 240; ? | 218 | 333–368 | 348 | 323 |
| Tibiotarsus II | 250 | 240; ? | 213 | 326–364 | 340 | 323 |
| Tibiotarsus III | 302 | 305; ? | 255 | 405–456 | 422 | 398 |
| Claw I | 135 | 133; 148 | 107 | 80–88 | 83 | 81 |
| Claw II | 135 | 133; 146 | 106 | 79–87 | 82 | 77 |
| Claw III | 118 | 114; 131 | 93 | 67–80 | 73 | 70 |
| Empodium I | 88 | 80; 95 | 65 | 39–46 | 42 | 40 |
| Empodium II | 75 | 84; 93 | 60 | 41–45 | 43 | 39 |
| Empodium III | 72 | 65; 90 | 62 | 41–48 | 44 | 42 |
| Th II seta m1 | 30 | 31; ? | 30 | 18–23 | 21 | 18 |
| Th II sensilla a | ? | ? | 12,6 | 4.3–12 | 7.0 | ? |
| Th III sensilla a | 13 | ? | 14,7 | 16–17 | 16.4 | 16 |
| Trichobothria AB distance | 87 | ? | 72 | 41–56 | 50 | 50 |
| Trichobothria BC distance | 60 | ? | 54 | 57–82 | 72 | 72 |
| Trichobothria ABC angle | 97° | ? | 96° | 154–168° | 160° | 166° |
| Abd c1 seta | 11.5 | 9; ? | 10.2 | 31–41 | 34 | 35 |
| Abd IV longest (dI-dIII)-1 seta | 85 | 88; ? | 86 | 127–136 | 130 | 127 |
| Abd VI longest circumanal seta | 56 | 47; ? | 52 | 43–58 | 52 | 46 |
| Appendices anales, length | 37 | 34; ? | - | 28–31 | 30 | - |
| App. anales, basal shaft width | 2.8 | 2.3; ? | - | 2.2–2.4 | 2.3 | - |
| App. anales, subapical max. width | 2.8 | 2.1; ? | - | 2.3–5 | 3.6 | - |
| Manubrium | 231 | 224; 207 | 186 | 225–240 | 230 | 221 |
| Dens | 265 | 240; 260 | 223 | 307–350 | 320 | 302 |
| Mucro | 140 | 120; 112 | 108 | 155–168 | 160 | 157 |
| Ratio | Arrhopalites profundus | Pygmarrhopalites rystsovi | ||
|---|---|---|---|---|
| Females | Male | Females | Male | |
| Antenna/head | 1.95 | 2.16 | 3.40 | 3.74 |
| Ant II/I | 2.08 | 2.08 | 2.78 | 2.60 |
| Ant III/I | 3.43 | 2.86 | 4.87 | 4.70 |
| Ant IV/I | 8.30 | 8.06 | 13.81 | 13.47 |
| Head/tibiotarsus I | 1.27 | 1.20 | 1.02 | 0.93 |
| Tibiotarsus II/I | 0.99 | 0.84 | 0.98 | 1.00 |
| Tibiotarsus III/I | 1.22 | 1.17 | 1.21 | 1.23 |
| Tibiotarsus I/claw I | 1.85 | 2.38 | 4.18 | 3.99 |
| Tibiotarsus II/claw II | 1.83 | 2.01 | 4.16 | 4.19 |
| Tibiotarsus III/claw III | 2.62 | 2.74 | 5.83 | 5.69 |
| Claw I/empodium I | 1.60 | 1.65 | 1.99 | 2.03 |
| Claw II/empodium II | 1.69 | 1.77 | 1.91 | 1.97 |
| Claw III/empodium III | 1.70 | 1.50 | 1.65 | 1.67 |
| Dens/mucro | 1.95 | 2.06 | 2.00 | 1.95 |
| Trichobothria AB/BC distance | 1.45 | 1.33 | 0.69 | 0.69 |
| Abd IV dI-1/Th2 m1 seta | 2.84 | 2.87 | 6.44 | 7.06 |
| Abd IV dI-1/claw III | 0.75 | 0.92 | 1.80 | 1.81 |
| Longest Abd IV/circumanal seta | 1.70 | 1.65 | 2.52 | 2.76 |
| Body total/tibiotarsus III | 3.31 | 3.61 | 2.60 | 2.39 |
| Body total/furca | 1.54 | 1.78 | 1.55 | 1.40 |
| Body total/dI-1 seta | 10.85 | 10.70 | 8.44 | 7.48 |
| Body total/claw I | 6.99 | 8.60 | 13.45 | 11.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargovitsh, R.S. Deep Troglomorphy: New Arrhopalitidae (Collembola: Symphypleona) of Different Life Forms from the Snezhnaya Cave System in the Caucasus. Diversity 2022, 14, 678. https://doi.org/10.3390/d14080678
Vargovitsh RS. Deep Troglomorphy: New Arrhopalitidae (Collembola: Symphypleona) of Different Life Forms from the Snezhnaya Cave System in the Caucasus. Diversity. 2022; 14(8):678. https://doi.org/10.3390/d14080678
Chicago/Turabian StyleVargovitsh, Robert S. 2022. "Deep Troglomorphy: New Arrhopalitidae (Collembola: Symphypleona) of Different Life Forms from the Snezhnaya Cave System in the Caucasus" Diversity 14, no. 8: 678. https://doi.org/10.3390/d14080678