
Species Distribution Modeling Reveals Recent Shifts in Suitable Habitat for Six North American Cypripedium spp. (Orchidaceae)

Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roe, G. In defense of Milankovitch. Geophys. Res. Lett. 2006, 33, L24703. [Google Scholar] [CrossRef] [Green Version]
- Eliseev, A.V.; Mokhoc, I.I. Influence of volcanic activity on climate change in the past several centuries: Assessments with a climate model of intermediate complexity. Atmos. Ocean. Phys. 2008, 44, 671–683. [Google Scholar] [CrossRef]
- Montañez, I.P.; McElwain, J.C.; Poilsen, C.J.; White, J.D.; DiMichele, W.A.; Wilson, J.P.; Griggs, G.; Hren, M.T. Climate, , and terrestrial carbon cycle linkages during late Palaeozoic glacial-interglacial cycles. Nat. Geosci. 2016, 9, 824–828. [Google Scholar] [CrossRef]
- PAGES 2k Consortium. Continental-scale temperature variability during the past two millennia. Nat. Geosci. 2013, 6, 339–346. [Google Scholar] [CrossRef]
- Neukom, R.; Steiger, N.; Gómez-Navarro, J.J.; Wang, J.; Wener, J.P. Werner. No evidence for globally coherent warm and cold periods over the preindustrial common era. Nature 2019, 571, 550–554. [Google Scholar] [CrossRef]
- Thornton, P.E.; Thorton, M.M.; Mayer, B.W.; Wei, Y.; Devaralonda, R.; Vose, R.S.; Cook, R.B. Daymet: Daily Surface Weather Data on a 1-km Grid of North America, Version 3; ORNL DAAC: Oak Ridge, TN, USA, 2016. [Google Scholar] [CrossRef]
- Jump, A.S.; Peñuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Franks, S.J.; Weber, J.J.; Aitken, S.N. Evolutionary and plastic responses to climate change in terrestrial plant populations. Evol. Appl. 2014, 7, 123–139. [Google Scholar] [CrossRef]
- Christmas, M.J.; Breed, M.F.; Lowe, A.J. Constraints to and conservation implications for climate change adaptation in plants. Conserv. Genet. 2016, 17, 305–320. [Google Scholar]
- Hamrick, J.L. Response of forest trees to global environmental changes. For. Ecol. Manag. 2014, 197, 323–335. [Google Scholar] [CrossRef]
- Kremer, A. Long-distance gene flow and adaptation of forest trees to rapid climate change. Ecol. Lett. 2012, 15, 378–392. [Google Scholar] [CrossRef] [Green Version]
- Lenoir, J.; Svenning, J.-C. Climate-related range shifts–a global multidimensional synthesis and new research directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Steinbauer, M.J. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- McGillivray, F.; Hudson, I.L.; Lowe, A.J. Herbarium collections and photographic images: Alternative data sources for phenological research. Biol. Conserv. 2010, 157, 172–177. [Google Scholar]
- Melles, S.J.; Fortin, M.-J.; Lindsay, K.; Badzinski, D. Expanding northward: Influence of climate change, forest connectivity, and population processes on a threatened species’ range shift. Glob. Chang. Biol. 2011, 17, 17–31. [Google Scholar] [CrossRef]
- Robbirt, K.M.; Davy, A.J.; Hutchings, M.J.; Roberts, D.L. Validation of biological collections as a source of phenological data for use in climate change studies: A case study with the orchid Ophrys sphegodes. J. Ecol. 2011, 99, 235–241. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Rymer, P.D.; Byrne, M.; Ruthrof, K.X.; Whinam, J.; McGeoch, M.; Bergstrom, D.M.; Guerin, G.R.; Sparrow, B.; Joseph, L.; et al. Impacts of recent climate change on terrestrial flora and fauna: Some emerging Australian examples. Austral. Ecol. 2019, 44, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.E.; Goulden, M.L. Goulden. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 11823–11826. [Google Scholar] [CrossRef] [Green Version]
- Crimmins, S.M.; Dobrowski, S.Z.; Greenberg, J.A.; Abatzoglou, J.T.; Mynsberge, A.R. Changes in climatic water balance drive downhill shifts in plant species’ optimum elevations. Science 2011, 331, 258–261. [Google Scholar] [CrossRef]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrío-Dillon, G.; de Ruffray, P.; Vidal, C.; Pierrat, J.-C.; Gégout, J.-C. Changes in plant community composition lag behind climate warming in lowland forest. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef]
- Nicholé, F.; Brzosko, E.; Till-Bottraud, I. Population viability analysis of Cypripedium calceolus in a protected area: Longevity, stability and persistence. J. Ecol. 2005, 93, 716–726. [Google Scholar] [CrossRef]
- Dressler, R.L. The Orchids: Natural History and Classification; England Harvard University Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Rasmussen, H. Terrestrial Orchids: From Seed to Mycotrophic Plant; Cambridge University Press: New York, NY, USA, 1995. [Google Scholar]
- Arditti, J.; Ghani, K.A. Tansley review no. 110: Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, S.; Cafasso, D.; Pellegrino, G.; Musacchio, A.; Widmer, A. Fine-scale phylogeographical analysis of Mediterranean Anacamptis palustris (Orchidaceae) populations based on chloroplast minisatellite and microsatellite variation. Mol. Ecol. 2003, 12, 2783–2792. [Google Scholar] [CrossRef]
- Trapnell, D.W.; Hamrick, J.L. Partitioning nuclear and chloroplast variation at multiple spatial scales in the Neotropical epiphytic orchid, Laelia rubescens. Mol. Ecol. 2004, 13, 2655–2666. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Trapnell, D.W. Using population genetic analyses to understand patterns of seed dispersal. Acta Oecol. 2011, 37, 641649. [Google Scholar] [CrossRef]
- Kartzinel, T.R.; Shefferson, R.P.; Trapnell, D.W. Relative importance of pollen and seed dispersal across a Neotropical mountain landscape for an epiphytic orchid. Mol. Ecol. 2013, 22, 6048–6059. [Google Scholar] [CrossRef]
- Broeck, A.V.; Landuyt, W.V.; Cox, K.; Bruyn, L.D.; Gyselings, R.; Oostermeijer, G.; Valentin, B.; Bozic, G.; Dolinar, B.; Illyés, Z.; et al. High levels of effective long-distance dispersal may blur ecotypic divergence in a rare terrestrial orchid. BMC Ecol. 2014, 14, 20. [Google Scholar]
- Minasiewicz, J.; Znaniecka, J.M.; Górniak, M.; Kawiński, A. Spatial genetic structure of an endangered orchid Cypripedium calceolus (Orhcidaceae) at a regional scale: Limited gene flow in a fragmented landscape. Conserv. Genet. 2018, 19, 1149–1460. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, D.W.; Hamrick, J.L.; Smallwood, P.A.; Kartzinel, T.R.; Ishibashi, C.D.; Quigley, C.T.C. Phylogeography of the Neotropical epiphytic orchid, Brassavola nodosa: Evidence for a secondary contact zone in northwestern Costa Rica. Heredity 2019, 123, 662–674. [Google Scholar] [CrossRef]
- Kotilínek, M.; Tĕšitelová, T.; Košnar, J.; Fibich, P.; Hemrová, L.; Koutecký, P.; Münzber-gová, Z.; Jersáková, J. Seed dispersal and realized gene flow of two forest orchids in a fragmented landscape. Plant Biol. 2020, 22, 522–532. [Google Scholar] [CrossRef]
- Trapnell, D.W.; Hamrick, J.L.; Ishibashi, C.D.; Kartzinel, T.R. Genetic inference of epiphytic orchid colonization; it may only take one. Mol. Ecol. 2013, 22, 3680–3692. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species, Version 2020-1; IUCN: Grand, Switzerland, 2020; Available online: https://www.iucnredlilst.org (accessed on 3 August 2021).
- Cribb, P. The Genus Cypripedium; Timber Press: Portland, OR, USA, 1997. [Google Scholar]
- Alexander, E.B.; Coleman, R.G.; Keeler-Wolf, T.; Harrison, S.P. Serpentine Geoecology of Western North America: Geology, Soils, and Vegetation; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Coleman, R.A. The Cypripedium of the United States and Canada. Part III: Arietinum, californicum, fasciculatum, guttatum, yatabeanum and X alaskanum. Orchids 2018, 87, 522–529. [Google Scholar]
- Stoutamire, W.P. Flower biology of the Lady’s-slippers (Orchidaceae: Cypripedium). Mich. Bot. 1967, 6, 159–175. [Google Scholar]
- Argue, C.L. The Pollination of North American Orchids: Volume 1: North of Florida and Mexico; Springer: New York, NY, USA, 2012. [Google Scholar]
- Withner, C.L. The Orchids: A Scientific Survey; The Ronald Press Co.: New York, NY, USA, 1959. [Google Scholar]
- Shefferson, R.P.; Bunch, W.; Cowden, C.C.; Lee, Y.-I.; Kartzinel, T.R.; Yukawa, T.; Downing, J.; Jiang, H. Does evolutionary history determine specificity in broad ecological interactions? J. Ecol. 2019, 107, 1582–1593. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: New York, NY, USA, 2010. [Google Scholar]
- Shefferson, R.P.; Jacquemyn, H.; Kull, T.; Hutchings, M.J. The demography of terrestrial orchid: Life history, population dynamics and conservation. Bot. J. Linn. Soc. 2020, 192, 315–332. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Warren, I.I.; Pulliam, H.R. Life history costs make perfect sprouting maladaptive in two herbaceous perennials. J. Ecol. 2014, 102, 1318–1328. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- GBIF.org GBIF Occurrence Download. 2018. Available online: https://doi.org/10.15468/dl.dujw5l (accessed on 2 February 2020).
- Pebesma, E.J.; Bivand, R.S. 2005. Classes and Methods for Spatial Data in R-R News 5: 2. Available online: https://cran.r-project.org/doc/Rnews (accessed on 20 September 2018).
- R: A language and environment for statistical computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 17 September 2018).
- Hijman, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling, R Package Version 1.1-4; 2017. Available online: https://CRAN.R-project.org/package=dismo (accessed on 5 August 2018).
- Hengl, T.; de Jesus, J.M.; Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marscallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Siuja, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- ESA. Land Cover CCI Produce User Guider Version 2. Tech. Rep. 2017. Available online: Maps.elie.ucl.ac.be/CCI/viewer/download/ESACCI-LC-Ph2-PUGv2_2.0.pdf (accessed on 1 April 2022).
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Clifford, P.; Richardson, S.; Hermon, D. Assessing the significance of the correlation between two spatial processes. Biometrics 1989, 45, 123–134. [Google Scholar] [CrossRef]
- Dutilleul, P. Modifying the t test for assessing the correlation between two spatial processes. Biometrics 1993, 49, 305–314. [Google Scholar] [CrossRef]
- Vallejos, R.; Osorio, F.; Bevilacqua, M. Spatial Relationships between Two Georeferenced Variables: With Applications in R.; Springer: New York, NY, USA, 2020. [Google Scholar]
- Liu, C.; Newell, G.; White, M. On the selection of thresholds for predicting species occurrence with presence-only data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Kou, X.; Li, Q.; Liu, S. Quantifying species’ range shifts in relation to climate change: A case study of Abies spp. in China. PLoS ONE 2011, 6, e23115. [Google Scholar] [CrossRef]
- Amante, C.; Eakins, B.W. ETOPO1 Arc-Minute Global Relief Model: Procedures, Data Sources and Analysis; NOAA: Washington, DC, USA, 2009. [Google Scholar]
- Patton, D.R. A diversity index for quantifying habitat “edge”. Wildl. Soc. Bull. 1975, 3, 171–173. [Google Scholar]
- Forman, R.T.T.; Godron, M. Landscape Ecology; John Wiley and Sons: New York, NY, USA, 1986. [Google Scholar]
- Cooke, S.S. The Edaphic Ecology of Two Western North American Composite Species. Ph.D. Thesis, University of Washington, Seattle, WA, USA, 18 August 1994. [Google Scholar]
- Armstrong, J.K.; Huenneke, L.F. Spatial and temporal variation in species composition in California grasslands: The interaction of drought and substratum. In The Vegetation of Ultramafic (Serpentine) Soils; Baker, A.J.M., Proctor, J., Reeves, R.D., Eds.; Andover: Hampshire, UK, 1992; pp. 213–233. [Google Scholar]
- Grime, J.P.; Brown, V.K.; Thompson, K.; Masters, G.J.; Hiller, S.H.; Clarke, I.P.; Askew, A.P.; Corker, D.; Kielty, J.P. The response of two contrasting limestone grasslands to simulated climate change. Science 2000, 289, 762–765. [Google Scholar] [CrossRef]
- Grime, J.P.; Fridley, J.D.; Askew, A.P.; Thompson, K.; Hodgson, J.G.; Bennett, C.R. Long-term resistance to simulated climate change in an infertile grassland. Proc. Natl. Acad. Sci. USA 2008, 105, 10028–10032. [Google Scholar] [CrossRef] [Green Version]
- Hughes, R.; Konrad, B.; Smirnoff, N.; Macnair, M.R. The role of drought tolerance in serpentine tolerance in the Mimulus guttatus Fisher ex DC. complex. S. Afr. J. Sci. 2001, 97, 581–586. [Google Scholar]
- Damschen, E.I.; Harrison, S.; Ackerly, D.D.; Fernandez-Going, B.M.; Anacker, B.L. Endemic plant communities on special soils: Early victims or hardy survivors of climate change? J. Ecol. 2012, 100, 1122–1130. [Google Scholar] [CrossRef]
- Fernandez-Going, B.M.; Anacker, B.L.; Harrison, S.P. Temporal variability in California grasslands: Soil type and species functional traits mediate response to precipitation. Ecology 2012, 93, 2104–2114. [Google Scholar] [CrossRef]
- Wear, D.N. Land use. In Southern Forest Resource Assessment. General Technical Report; Wear, D.N., Greis, J.G., Eds.; USDA, Forest Service: Washington, DC, USA, 2002; pp. 153–174. [Google Scholar]
- Young, A.; Boyle, T.; Brown, T. The population genetic consequences of habitat fragmentation for plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef]
- Laurance, W.F.; Lovejoy, T.E.; Vasconcelos, H.L.; Bruna, E.M.; Didham, R.K.; Stouffer, P.C.; Gascon, C.; Bierregaard, R.O.; Laurance, S.G.; Sampaio, E. Ecosystem decay of Amazonian forest fragments: A 22-year investigation. Conserv. Biol. 2002, 16, 605–618. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.H. Extinction risk in fragmented habitat. Anim. Conserv. 2004, 7, 181–191. [Google Scholar] [CrossRef]




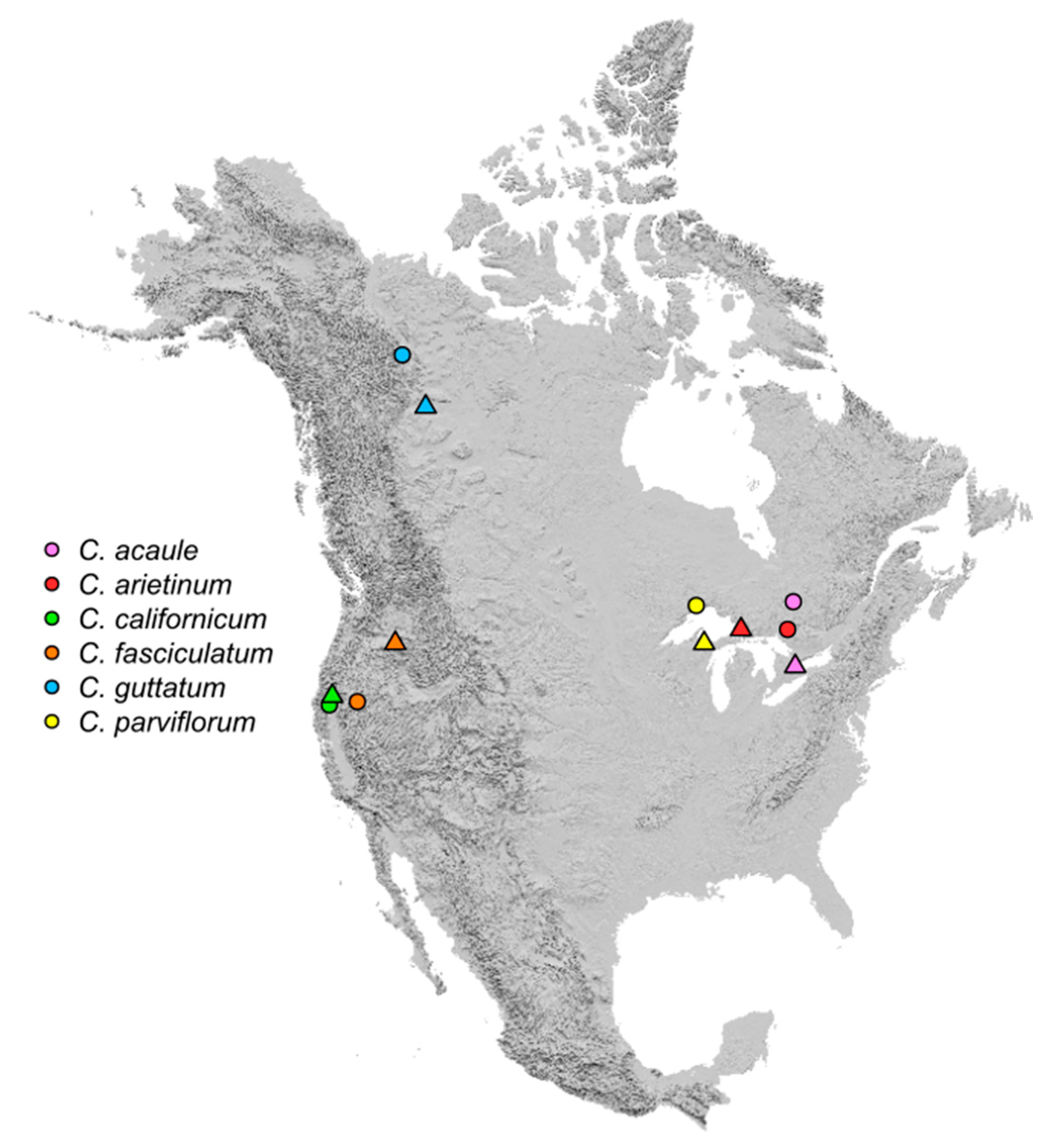{kind=link}
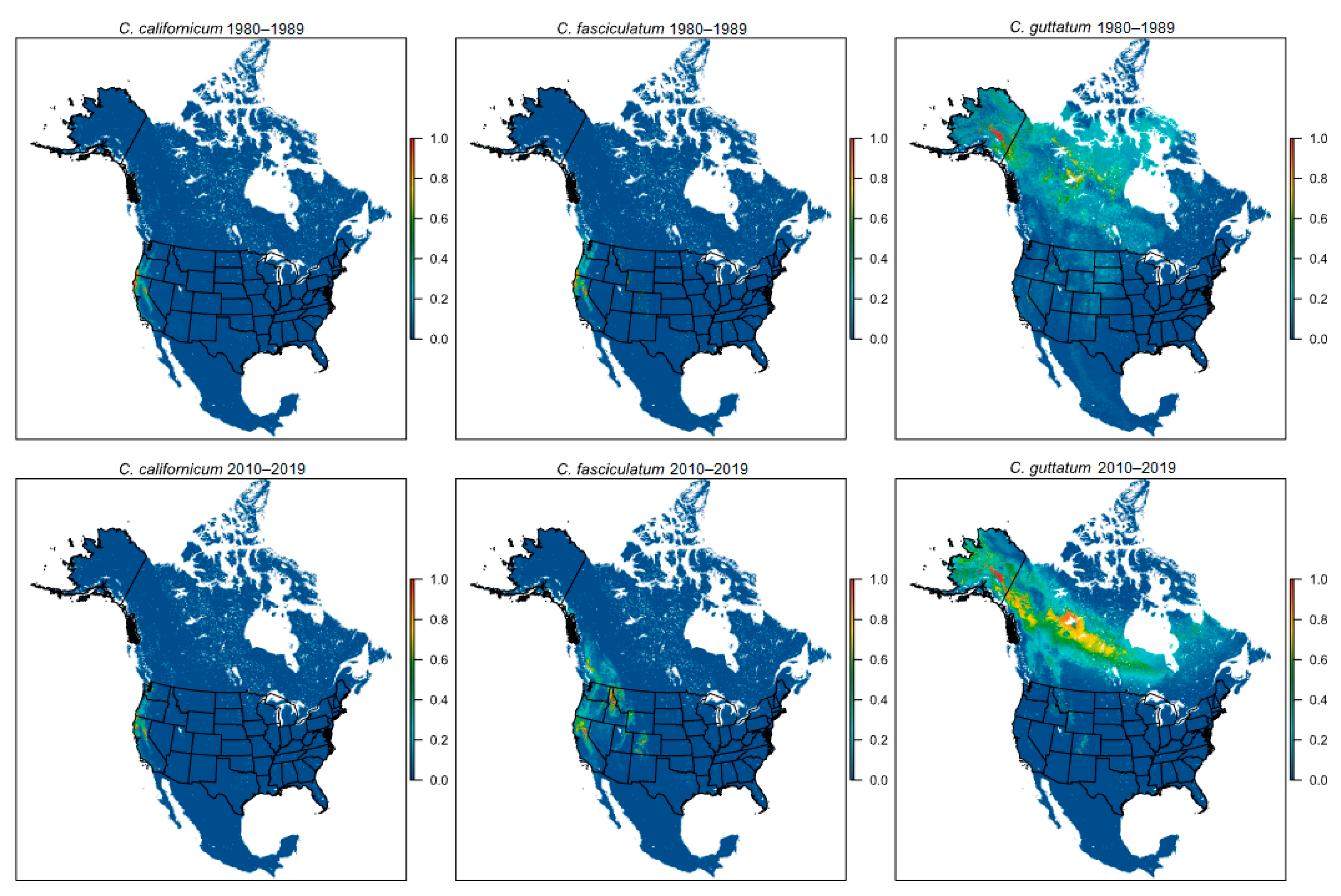{kind=link}
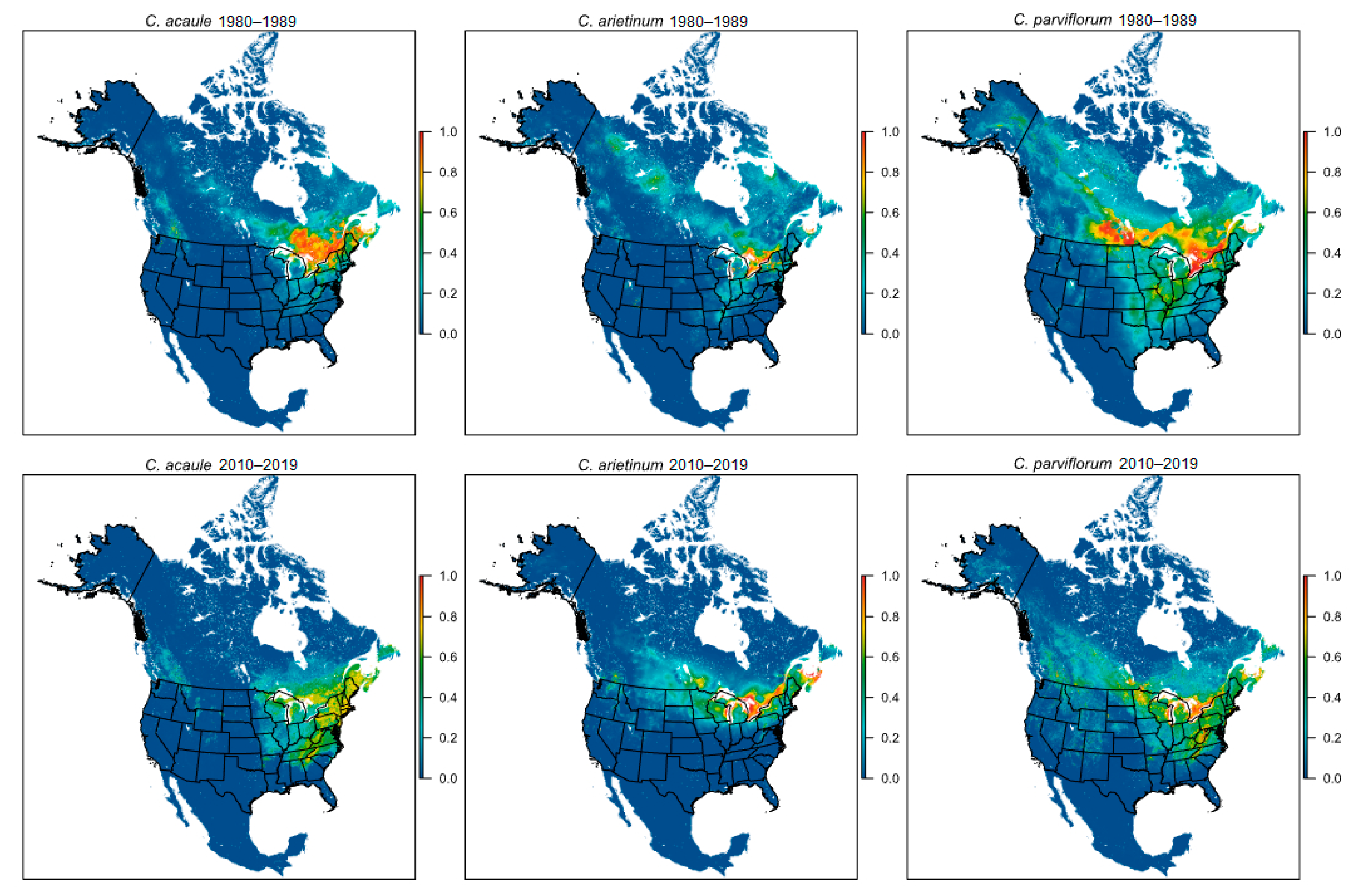{kind=link}
| Species | N (ETI) | N (LTI) | PCC (ETI) | PCC (LTI) | Ω | Δ Area (ha) | %Δ Area | Centroid Shift (km) | Δ Elevation (m) | %Δ Elevation | %Δ SIP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| West | |||||||||||
| C. californicum | 35 | 78 | 71.4% | 92.3% | 47.4% | 564 | 9.1% | 67 | −139 | −14.7% | 179.2% |
| C. fasciculatum | 17 | 53 | 64.7% | 94.3% | 4.2% | 79,599 | 2062.2% | 501 | 321 | 24.8% | 40.1% |
| C. guttatum | 12 | 34 | 91.7% | 82.4% | 10.7% | 104,193 | 408.0% | 411 | 84 | 20.1% | −66.0% |
| Mean | 21.3 | 55.0 | 75.93% | 89.67% | 20.77% | 61,452.0 | 826.43% | 326.3 | 88.6 | 10.07% | 51.10% |
| East | |||||||||||
| C. acaule | 82 | 4654 | 81.7% | 97.2% | 39.7% | 41,249 | 26.0% | 466 | 4 | 1.1% | 93.7% |
| C. arietinum | 11 | 91 | 72.7% | 93.4% | 11.5% | 114,714 | 553.5% | 336 | 88 | 35.8% | −81.7% |
| C. parviflorum | 81 | 1560 | 67.9% | 92.7% | 23.9% | 156,983 | 111.2% | 279 | 106 | 32.3% | 83.8% |
| Mean | 58.0 | 2101.7 | 74.10% | 94.43% | 25.03% | 104,315.3 | 230.23% | 360.3 | 66.0 | 23.07% | 31.93% |
| Overall mean | 39.7 | 1078.4 | 75.02% | 92.05% | 22.90% | 82,883.7 | 528.33% | 343.3 | 77.3 | 16.57% | 41.52% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smallwood, P.A.; Trapnell, D.W. Species Distribution Modeling Reveals Recent Shifts in Suitable Habitat for Six North American Cypripedium spp. (Orchidaceae). Diversity 2022, 14, 694. https://doi.org/10.3390/d14090694
Smallwood PA, Trapnell DW. Species Distribution Modeling Reveals Recent Shifts in Suitable Habitat for Six North American Cypripedium spp. (Orchidaceae). Diversity. 2022; 14(9):694. https://doi.org/10.3390/d14090694
Chicago/Turabian StyleSmallwood, Patrick A., and Dorset W. Trapnell. 2022. "Species Distribution Modeling Reveals Recent Shifts in Suitable Habitat for Six North American Cypripedium spp. (Orchidaceae)" Diversity 14, no. 9: 694. https://doi.org/10.3390/d14090694
APA StyleSmallwood, P. A., & Trapnell, D. W. (2022). Species Distribution Modeling Reveals Recent Shifts in Suitable Habitat for Six North American Cypripedium spp. (Orchidaceae). Diversity, 14(9), 694. https://doi.org/10.3390/d14090694