Abstract
The Adriatic sturgeon, Acipenser naccarii (Bonaparte, 1836), is a critically endangered tetraploid endemism of the Adriatic region; it has been targeted, over the last 20 years, by different conservation programs based on controlled reproduction of captive breeders followed by the release of their juvenile offspring; its preservation would greatly benefit from the correct and coordinated management of the residual genetic variability available in the different captive stocks. In this sense, the setup of an efficient parental allocation procedure would allow identifying familiar groups and establishing informed breeding plans, effectively preserving genetic variation. However, being the species tetraploid, the analyses often deal with complex genome architecture and a preliminary evaluation of allele segregation patterns at different chromosomes is necessary to assess whether the species can be considered a pure tetraploid, as previously observed at some loci, or if a more complex situation is present. Here we study the segregation at 14 microsatellites loci in 12 familiar groups. Results support in different families the tetrasomic segregation pattern at 11 markers and the disomic segregation at three markers. The Adriatic sturgeon thus shows a mixed inheritance modality. In this species, and likely in other sturgeons, accurate knowledge of the loci used for paternity analysis is therefore required.
1. Introduction
Sturgeons are the most endangered group of species, according to the International Union for Conservation of Nature (IUCN, July 2021) (http://www.iucnredlist.org, accessed on 8 August 2022). For this reason, sturgeons are targeted by several conservation efforts that often include restocking programs with juveniles produced in captivity; these ex-situ conservation activities must necessarily be supported by studies aimed at preserving the residual genetic diversity through long-term breeding programs [1]. However, genetic analyses on sturgeons deal with complex genomes and various levels of ploidy, due to independent events of whole-genome duplication [2,3]. The first event of duplication took place in the Acipenseriformes’ common ancestor starting from sixty chromosomes. Then, secondary events of duplication occurred in the Pacific and Atlantic clades leading to a total of 240 chromosomes [2]. Finally, a third event led to the unique number of 360 chromosomes observable in Acipenser brevirostrum (Lesueur, 1818) [4]. The number of chromosomes associated with the distinct levels of ploidy has been the subject of an extensive debate between two main positions. The first argues that, since the condition with 120 chromosomes results from a duplication event in the common ancestor, the species with 120 and 240 chromosomes must be considered tetraploid and octaploid, respectively. The second position, taking into account the functional reduction of ploidy that follows whole genome duplications, attributes to the two groups a condition of diploidy and tetraploidy, respectively [5]. The two views use different criteria to define the nominal ploidy, the number of duplications and the functional activity of genes, respectively, and are both correct and fully compatible [3].
The Adriatic sturgeon, Acipenser naccarii (Bonaparte, 1836), a species with 240 chromosomes, is in general considered a functional tetraploid based on the analyses of 28S and 5S rDNA through in situ hybridization [6]; this is also confirmed by most of the microsatellites analysed in this species which mostly show up to 4 alleles per individuals. However, some loci consistently showed more than 4 alleles in many individuals [7]; this could be due to the duplication of the region of such microsatellites but, a still incomplete process of functional reduction following genome duplication cannot be excluded as already proposed also for other sturgeon species [8,9].
For these reasons, loci to be used for applied purposes, such as parental allocation and kinship analysis, should be carefully selected and their functional ploidy should be preliminarily assessed. Moreover, these applications require a careful preliminary investigation also on the segregation modalities which, in tetraploids, can be of different types. In fact, polyploidization can originate through the fusion of unreduced gametes at intraspecific or interspecific levels, leading to two types of conditions in tetraploids, called autotetraploidy and allotetraploidy, respectively. The origins of polyploidization have important implications on the segregation patterns of the alleles within gametes. In complete autotetraploids, there are always four homologous chromosomes, and random pairs of bivalents and quadrivalents are possible during meiosis (see Figure 1 for graphical support) [10]; this condition leads to tetrasomic inheritance, which means that all allelic combinations within gametes are possible; this is not the case in complete allotetraploids, where each tetrad is composed of two sets of homeologous chromosomes, originating from the two parental species. In this situation, the homeologous chromosomes do not form pairs, leading to disomic inheritance, where only four out of six allelic combinations are possible [10]; these are two extreme cases and intermediate conditions are observable when the chromosomes have different degrees of preferential pairing. Indeed, the inheritance may shift from tetrasomic to disomic or vice versa [10]. For example, in autotetraploids, fertility and karyotype stability can be negatively impacted by imperfect multivalent pairing, thus promoting diploidization and consequent shifting to disomy; on the contrary, in allotetraploids, the homeologous chromosomes from two distinct parental species could maintain some degree of genetic affinity permitting the competition with the homologous pair during meiotic interactions with a certain degree of tetrasomic inheritance [11]. Understanding the mechanisms of chromosomal segregation in a tetraploid species can have important conservation implications, for example when the development of parental allocation methods is required.
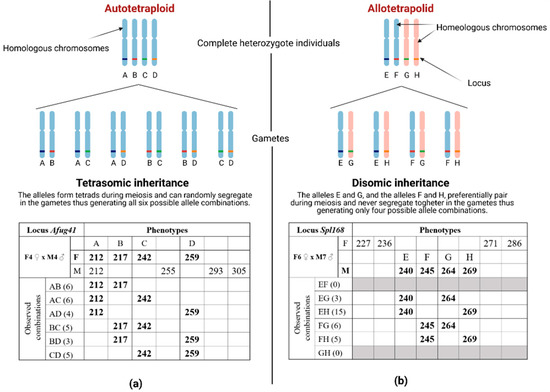
Figure 1.
Expected segregation patterns under pure disomy (a) or tetrasomy (b), respectively expected in Autotetraploid and Allotetraploid genomes.
In the Adriatic sturgeon, allele segregation was previously investigated at only 7 loci using microsatellite markers [12]. All loci showed a tetrasomic inheritance pattern, pointing at the probable autopolyploidization origin of this species. However, as secondary differentiation of some homologous chromosomes cannot be excluded and different segregation patterns can be followed by different chromosomes, we took advantage of the recent availability of new complete family groups and new isolated and tested microsatellite loci not yet explored, to provide a deeper insight into the mode of chromosome segregation in this species.
This is a key step not only to have a better understanding of the karyotype in the Adriatic sturgeon but also for the correct interpretation of the genetic analysis and parental allocation which in this species can have multiple applications. Firstly, the distinction of extremely rare individuals of wild origin from released ones. Secondly, the identification of groups of siblings existing in the different farms by assigning everyone to his pair of parents of the F0 generation. Finally, knowing the segregation patterns is crucial when the generation of virtual genotypes starting from observed genetic profiles is needed [13,14].
Thanks to the possibility of making 12 distinct crosses and raising their progeny separately within the project ENDEMIXIT (https://endemixit.com/, accessed on 8 August 2022), many new informative familiar groups became available for the analysis of segregation patterns at a much higher number of loci than those available in the past. The main purpose of the present study is therefore to exhaustively describe the modalities of microsatellite alleles inheritance in the Adriatic sturgeon with the following objectives: (a) verifying if the inferred pure tetrasomy is confirmed on a high number of loci, (b) providing a significant contribution to the management of the residua genetic diversity of this critically endangered species, and (c) shedding light on the functional ploidy level across its genome.
2. Materials and Methods
2.1. Samples and DNA Purification
Most samples analyzed in the present study were obtained from the aquaculture plant Storione Ticino (Cassolnovo, Italy). The reproduction of six females (F2, F4, F5, F6, F7, and F8) and six males (M2, M4, M5, M6, M7, and M8) was performed in different combinations. For each parent, a fin clip was collected for genotyping and, for each cross, the progeny was reared in captivity and spontaneously dead animals were collected and stored in ethanol. Moreover, five familiar groups (Nacc7 ♀ × Nacc5 ♂, Nacc8 ♀ × Nacc31 ♂, Nacc19 ♀ × Nacc17 ♂, Nacc19 ♀ × Nacc30 ♂, Nacc28 ♀ × Nacc23 ♂) derived from crosses performed in the past and for which fingerlings were available were also used. In total, 21 adult parents (12 parent pairs) and 376 fingerlings (with about 30 individuals per family) were collected (Table 1).

Table 1.
Screening of microsatellite loci.
Genomic DNA was extracted from breeder’s fin clips (10–100 mg) and from offspring’s muscular tissue, using Euroclone spinNAker Universal Genomic DNA mini kit (Euroclone) and stored at −20 °C till their processing for microsatellite analysis.
2.2. Selection of Loci and Genotyping
Loci analysed in the present study were selected based on the possibility of unambiguously tracing the genetic contribution of at least one of the two parents in at least one of the available families. For this reason, all loci in which there were no complete heterozygotes or there were individuals with more than four alleles were discarded. In fact, it is known that the Adriatic sturgeon presents a minority of loci at which more than four alleles were can be observed in different individuals; thus, segregation anomalies cannot be detected [7]; these extra numerary alleles could originate from duplication or from a locally unreduced octaploid condition. In fact, we recall that the Adriatic sturgeon is functionally a tetraploid but evolutionarily it is an octaploid [3]. Nevertheless, segregation anomalies were also observed in some individuals of each family that were accordingly excluded from the analysis.
All breeders were genotyped at 21 microsatellite loci (Table 1) [7,15,16,17,18,19,20] to select the informative Family/Locus combinations for the assessment of chromosomal segregation. For each selected combination, the progeny was also amplified and genotyped. Tracking segregation in the progeny requires the satisfaction of some features, such as (i) the complete heterozygosity (four different alleles) of at least one parent to ensure that each allele can unambiguously mark the segregation of its own chromosome and (ii) no more than an allele shared by the two parents to avoid ambiguity in following the alleles transmission to the progeny [12].
A total of 33 Family/Locus combinations were finally selected and approximately 30 fingerlings each were genotyped (Table 1). In 22 out of 33 case studies only one breeder of the parent pair was completely heterozygote and therefore informative to follow the segregation of the alleles, while for the remnant 11 Family/Locus combinations the segregation pattern was followed for both breeders of the parent pair as both they showed completely heterozygous genotypes with a single allele shared at most (Table 1).
Microsatellite loci were amplified from genomic DNA in a 10 µL reaction: 1X Master Mix Buffer (QIAGEN), 0.2 µM of each primer and about 50 ng of genomic DNA. Amplifications were performed on SimpliAmp Thermal Cycler (Thermo Fisher Scientific, Bologna, Italy). Amplifications were checked on 1.8% agarose gel in TBE1X stained with GelRed (BIOTIUM, Fremont, Canada, GelRed™ Nucleic Acid Stain). Genotyping was performed on ABI PRISM 310 Genetic Analyzer (external service, BMR Genomics, Padova, Italy). For microsatellite scoring the software GeneMarker Version 1.95 (SoftGenetics LLC, State College, PA, USA) was used.
2.3. Data Analyses
Scoring was performed by two operators, independently, with GeneMarker Version 1.95 (SoftGenetics LLC). The final dataset was carefully checked, and each individual was controlled for perfect Mendelian inheritance within his Family/Locus combination under the assumption that the observed genotypes at a given locus should be compatible with the inheritance of two allele copies from each parent. Individuals that for different possible reasons discussed later did not match this assumption were discarded. Since discordances with the above-accepted criterion can be attributed to anomalies in chromosomal segregation in the gametes of one parent, which are independent of what happened in the mating partner’s gametes, it was decided not to discard those animals whose segregation anomaly was due to a problem in the uninformative parent. Limiting our observations to those animals that met standard inheritance of two out of four alleles, we consciously excluded possible genotypes that were not necessarily derived from segregation anomalies such as the case of double reduction, a phenomenon widely observed in autotetraploid plants in which the four homolog chromosomes may form multivalents [21,22] in which identical alleles carried on the sister chromatids may enter the same gamete with consequent segregation of two copies of the same allele even if it was present in a single copy in the parental genome [23]. We have decided to limit ambiguities by disregarding this phenomenon and excluding all the individuals that could be compatible with it focusing on animals in which alleles were unambiguously traceable.
As proposed by Stift et al. (2008) [10], Likelihood Ratio Tests (LRT) with 1 df were applied to compare the null model of tetrasomy with the other alternative models intermediate between disomic and tetrasomic. For each parent-locus combination and each alternative inheritance model, the log-likelihood was estimated from constrained nonlinear regression models, using SPSS syntax as reported in the original reference [10]. The Sequential Bonferroni correction [24] was applied to adjust significance levels (p < 0.05) for multiple comparisons across loci and families.
3. Results
From 22 to 32 individuals for each family-locus combination were successfully genotyped. In some cases, a few animals were discarded for unreliable profiles or, after the allele scoring, due to the presence of segregation anomalies of alleles inherited from the informative parent (Table 2).
Table 2.
Likelihood Ratio Test.
At 11 (AfuG132, AfuG41, An20, Anac6784, Anac3133, AnacA6, AnacB11, AnacB7, AnacE4, AoxD234, AoxD241) out of 14 informative loci the LRT did not lead to the rejection of the null model of inheritance thus supporting the hypothesis of tetrasomy. At almost all these loci, indeed, all six allele combinations were observed in almost all tested families (Figure 1a, Table 2, Figure A1 of Appendix A). The only exceptions were the loci AoxD234 and Anac-3133 at a single family each showing only five allele combinations. However, in both cases, the strict disomic inheritance was excluded after correction for multiple tests (Table 2). In these two families, the sixth combination is expected to be detectable by increasing the sample size. Five (An20, AnacA6, AnacE4, AoxD234, and AoxD241) of the 11 loci showing tetrasomic inheritance were already tested to assess the inheritance pattern in different familiar groups of Adriatic sturgeon [12] and their tetrasomy has been here confirmed. Specifically, the locus AnacA6 here analysed at a single family showed only four allele combinations but, missing combinations are not compatible with disomic inheritance modality and even in this case the null model was not rejected.
On the contrary, a significant rejection of the null model was observed at three loci, Spl163, AoxD161, and Spl168, analysed respectively at one, three and six families and never analysed before in other studies. In some cases, the analysed families have both parents informative for segregation and agree in suggesting a disomic mode of inheritance (Table 2, Figure 1b, Figure A1 Appendix A). The disomic model fitted the observed allele combination frequencies significantly better than the null model, and the parameter τ equal to zero indicates a full disomic inheritance at almost all Family/Locus combinations. The only two exceptions were observed in the segregation of the alleles of female F8 at locus AoxD161 and male Nacc31 at locus Spl168 (Table 2). In these two cases, five combinations of alleles were present, the parameter τ assumed a very low value confirming a strong degree of preferential pairing during meiosis but the rejection of the null model was not significant after the Bonferroni correction, thus suggesting possible imperfect preferential pairing.
Expected allele combinations in tetrasomic and disomic mode of inheritance in tetraploids. (a) Observed allele combinations inherited from a complete heterozygote parent in autotetraploids in which tetrads are generated during meiosis and random segregation of allele pairs is expected. In brackets, the number of individuals carrying the relative allele combination is reported. (b) Observed allele combinations inherited from a complete heterozygote parent in allotetraploids in which a preferential pair between homologous chromosomes occurs. Only four possible gametes are expected. Alleles of the parent for which the segregation is reported in each table are marked in bold and by a letter used to indicate the observed combinations. Complete tables for all loci are reported in Appendix A. (Created with BioRender.com, accessed on 8 August 2022).
4. Discussion
The segregation pattern observed in the present study at most loci indicates that the Adriatic sturgeon can be considered predominantly tetraploid with a tetrasomic inheritance pattern. The presence of three disomic loci, however, indicates that the functional diversification process is at different stages in different parts of the genome, with some regions possibly still octaploid, most tetraploid, and some others in which the degree of divergence has gone up to a condition of double diploidy. We also observed the presence of two loci with a marked tendency to disomy with a few unexpected allele combinations, pointing to a possible imperfect preferential pairing expected at intermediate stages of functional diploidization [10,11]; this coexistence of different ploidy levels was previously described in other organisms. In plants, for example, the co-existence of different segregation patterns (e.g., tetrasomic and disomic) with different intermediate degrees of preferential pairing among chromosomes provides evidence of a process of functional reduction of ploidy which reasonably cannot occur simultaneously throughout the genome, but which is the result of a progressive differentiation [11,25].
As for the sturgeons, in other species the presence of different degrees of ploidy has been deduced based on the maximum number of alleles per individual present at the different loci [16] and, in some cases, the Mendelian transmission in the progeny has also been verified [15,26,27]. However, the selection of the cross/locus combinations to allow the traceability of every single allele and consequently to distinguish between the different modes of tetraploid segregation (e.g., disomic, tetrasomic or intermediate) has been conducted to our knowledge only on the Adriatic sturgeon.
The evidence that the level of homology between chromosomes within the Adriatic sturgeon genome is likely to vary from chromosome to chromosome and that different parts of the genome may consequently have different degrees of functional ploidy (2 to 8) should be considered when characterizing the genome of this species and probably of sturgeons in general. Genome assembly procedures must contemplate the possibility that different regions are present with a variable number of copies.
Whatever the evolutionary origin and implications of our results, which can be better investigated only when the genome of this species will be available, the identification of different inheritance pattern at different markers may have practical consequences for the conservation of the Adriatic sturgeon. In fact, our findings suggest that before developing parental allocation methods, a preliminary analysis of the loci used is recommended; this would ease the reconstruction of individual genealogies of animals kept as captive breeders and the reallocation of any individual recaptured after release. Another interesting and relatively unexplored aspect of sturgeon ex situ conservation in which a clear knowledge of the inheritance patterns at different loci could be useful is the monitoring of the genomic impact of breeding protocols. Captive breeding is usually performed following standardized protocols of hormonal induction of egg and milt release and fertilization is done in an excess of sperms. Then, the resulting viable progeny is released without verifying if the procedure used had some effect on their genomic asset, for example, by inducing aneuploidies, which are a common phenomenon in some captive sturgeon stocks [28]. The release in nature of genetically anomalous individuals should be avoided as the consequences that this can have on the following generations and on their reproductive efficiency is unknown and, in the case of animals with long generation times, could be revealed after decades. Random screening of familiar groups to verify the correct parents-to-progeny segregation at both disomic and tetrasomic loci could significantly contribute to reducing the potential impact of genomic anomalies on natural populations.
5. Conclusions
Thanks to the availability of some family groups not previously analyzed, it was possible to better investigate the patterns of chromosomal segregation in the polyploid Adriatic sturgeon. The picture that emerged is that of an extremely dynamic genome in which it is possible to find the co-existence of regions with different degrees of ploidy, some of which retain the legacy of ancient duplications and others show a dynamic reduction of functional ploidy; this pattern is probably shared with other polyploid sturgeons in which different patterns of segregation have been observed [27], but it is not known whether the same genomic regions are involved.
Additional studies are required to better characterize the distribution of ploidy across the genome; this will likely contribute to explaining some specific features of the sturgeons, such as the ability of species with different degrees of ploidy to hybridize and produce viable offspring [29]; this extreme genomic plasticity could somehow be linked to a high degree of genomic redundancy; it would also be very interesting to verify whether the regions with different segregation patterns are somehow related to the size of the chromosomes, which in sturgeons is known to be very variable, with a small number of large chromosomes and a high number of micro-chromosomes. However, until complete genomes of good quality are available, it is difficult to move beyond speculation on these topics.
Author Contributions
Conceptualization, S.D.P., E.B. and L.C.; methodology, S.D.P., E.B. and L.C.; investigation, S.D.P. and S.G.B.; writing—original draft preparation, S.D.P. and L.C.; writing—review and editing, E.B., V.H.M.-M. and G.B.; project administration, G.B. and L.C.; funding acquisition, G.B. and L.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Italian PRIN 2017, grant number 201794ZXTL.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of the University of Padova (OPBA)—(protocol code 421922 of 14 October 2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank the Province of Vicenza and the “Bacino Pesca B (VI)” for the hospitality in the “Oasi le Sorgenti” Natural Reserve and for the logistical support and the Storione Ticino aquaculture farm for performing all crosses that allowed this study.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A






Figure A1.
Total segregation results. Single-locus microsatellite inheritance for each studied sturgeon family. Microsatellite alleles of parents are reported as sizes in bp. For families in which both parents are informative, two different schemes are shown. The 4 alleles of the informative parents are highlighted in bold and marked with capital letters A, B, C, D for females and E, F, G, H for males, also used to label allele combinations observed in the progeny. The number of F1 in which each combination has been observed is reported in brackets. Missing combinations are marked in grey.
Figure A1.
Total segregation results. Single-locus microsatellite inheritance for each studied sturgeon family. Microsatellite alleles of parents are reported as sizes in bp. For families in which both parents are informative, two different schemes are shown. The 4 alleles of the informative parents are highlighted in bold and marked with capital letters A, B, C, D for females and E, F, G, H for males, also used to label allele combinations observed in the progeny. The number of F1 in which each combination has been observed is reported in brackets. Missing combinations are marked in grey.






References
- Ludwig, A. A Sturgeon View on Conservation Genetics. Eur. J. Wildl. Res. 2005, 52, 3–8. [Google Scholar] [CrossRef]
- Peng, Z.; Ludwig, A.; Wang, D.; Diogo, R.; Wei, Q.; He, S. Age and Biogeography of Major Clades in Sturgeons and Paddlefishes (Pisces: Acipenseriformes). Mol. Phylogenetics Evol. 2007, 42, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Fontana, F.; Congiu, L.; Mudrak, V.A.; Quattro, J.M.; Smith, T.I.; Ware, K.; Doroshov, S.I. Evidence of Hexaploid Karyotype in Shortnose Sturgeon. Genome 2008, 51, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Nam, Y.K.; Noh, J.K.; Park, C.H.; Chapman, F.A. Karyotype of North American Shortnose Sturgeon Acipenser Brevirostrum with the Highest Chromosome Number in the Acipenseriformes. Ichthyol. Res. 2005, 52, 94–97. [Google Scholar] [CrossRef]
- Fontana, F.; Zane, L.; Pepe, A.; Congiu, L. Polyploidy in Acipenseriformes: Cytogenetic and Molecular Approaches. In Fish Cytogenetics; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; Science Publisher, Inc.: Enfield, NH, USA, 2007; pp. 385–403. [Google Scholar]
- Fontana, F.; Lanfredi, M.; Chicca, M.; Congiu, L.; Tagliavini, J.; Rossi, R. Fluorescent in Situ Hybridization with rDNA Probes on Chromosomes of Acipenser ruthenus and Acipenser naccarii (Osteichthyes, Acipenseriformes). Genome 1999, 42, 1008–1012. [Google Scholar] [CrossRef]
- Boscari, E.; Vidotto, M.; Martini, D.; Papetti, C.; Ogden, R.; Congiu, L. Microsatellites from the Genome and the Transcriptome of the Tetraploid Adriatic Sturgeon, Acipenser naccarii (Bonaparte, 1836) and Cross-Species Applicability to the Diploid Beluga Sturgeon, Huso huso (Linnaeus, 1758). J. Appl. Icht. 2015, 31, 977–983. [Google Scholar] [CrossRef]
- Ludwig, A.; Belfiore, N.M.; Pitra, C.; Svirsky, V.; Jenneckens, I. Genome Duplication Events and Functional Reduction of Ploidy Levels in Sturgeon (Acipenser, Huso and Scaphirhynchus). Genetics 2001, 158, 1203–1215. [Google Scholar] [CrossRef]
- Havelka, M.; Hulák, M.; Bailie, D.A.; Prodöhl, P.A.; Flajšhans, M. Extensive Genome Duplications in Sturgeons: New Evidence from Microsatellite Data. J. Appl. Ichthyol. 2013, 29, 704–708. [Google Scholar] [CrossRef]
- Stift, M.; Berenos, C.; Kuperus, P.; van Tienderen, P.H. Segregation Models for Disomic, Tetrasomic and Intermediate Inheritance in Tetraploids: A General Procedure Applied to Rorippa (Yellow Cress) Microsatellite Data. Genetics 2008, 179, 2113–2123. [Google Scholar] [CrossRef]
- Soares, N.R.; Mollinari, M.; Oliveira, G.K.; Pereira, G.S.; Vieira, M.L.C. Meiosis in Polyploids and Implications for Genetic Mapping: A Review. Genes 2021, 12, 1517. [Google Scholar] [CrossRef]
- Boscari, E.; Barbisan, F.; Congiu, L. Inheritance pattern of Microsatellite Loci in the Polyploid Adriatic Sturgeon (Acipenser naccarii). Aquaculture 2011, 321, 223–229. [Google Scholar] [CrossRef]
- Congiu, L.; Pujolar, J.M.; Forlani, A.; Cenadelli, S.; Dupanloup, I.; Barbisan, F.; Galli, A.; Fontana, F. Managing Polyploidy in Ex Situ Conservation Genetics: The Case of the Critically Endangered Adriatic Sturgeon (Acipenser naccarii). PLoS ONE 2011, 6, e18249. [Google Scholar] [CrossRef] [PubMed]
- Boscari, E.; Pujolar, J.M.; Dupanloup, I.; Corradin, R.; Congiu, L. Captive Breeding Programs Based on Family Groups in Polyploid Sturgeons. PLoS ONE 2014, 9, e110951. [Google Scholar] [CrossRef] [PubMed]
- May, B.; Krueger, C.C.; Kincaid, H.L. Genetic Variation at Microsatellite Loci in Sturgeon: Primer Sequence Homology in Acipenser and Scaphirhynchus. Can. J. Fish. Aquat. Sci. 1997, 54, 1542–1547. [Google Scholar] [CrossRef]
- Welsh, A.B.; Blumberg, M.; May, B. Identification of Microsatellite Loci in Lake Sturgeon, Acipenser Fulvescens, And Their Variability in Green Sturgeon, A. medirostris. Mol. Ecol. Notes 2002, 3, 47–55. [Google Scholar] [CrossRef]
- Zane, L.; Patarnello, T.; Ludwig, A.; Fontana, F.; Congiu, L. Isolation and Characterization of Microsatellites in the Adriatic Sturgeon (Acipenser naccarii). Mol. Ecol. Notes 2002, 2, 586–588. [Google Scholar] [CrossRef]
- Forlani, A.; Fontana, F.; Congiu, L. Isolation of Microsatellite Loci from the Endemic and Endangered Adriatic Sturgeon (Acipenser naccarii). Conserv. Genet. 2007, 9, 461–463. [Google Scholar] [CrossRef]
- Henderson-Arzapalo, A.; King, T.L. Novel Microsatellite Markers for Atlantic Sturgeon (Acipenser oxyrinchus) Population De-Lineation and Broodstock Management. Mol. Ecol. Notes 2002, 2, 437–439. [Google Scholar] [CrossRef]
- McQuown, E.C.; Sloss, B.L.; Sheehan, R.J.; Rodzen, J.; Tranah, G.J.; May, B. Microsatellite Analysis of Genetic Variation in Sturgeon: New Primer Sequences for Scaphirhynchus and Acipenser. Trans. Am. Fish. Soc. 2000, 129, 1380–1388. [Google Scholar] [CrossRef]
- Sybenga, J. Chromosome Pairing Affinity and Quadrivalent Formation in Polyploids: Do Segmental Allopolyploids Exist? Genome 1996, 39, 1176–1184. [Google Scholar] [CrossRef]
- Lloyd, A.; Bomblies, K. Meiosis in Autopolyploid and Allopolyploid Arabidopsis. Curr. Opin. Plant Biol. 2016, 30, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Wright, L.; Ponce, O.; Chen, J.; Prashar, A.; Sanchez-Moran, E.; Luo, Z.; Compton, L. Varietal Variation and Chromosome Behaviour During Meiosis in Solanum Tuberosum. Heredity 2020, 125, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.D.; Van de Velde, J.D.; Novikova, P.Y. Inference of Polyploidy Origin and Inheritance Mode from Population Genomic Data. BioRxiv. [CrossRef]
- Pyatskowit, J.D.; Krueger, C.C.; Kincaid, H.L.; May, B. Inheritance of Microsatellite Loci in the Polyploid Lake Sturgeon (Acipenser fulvescens). Genome 2001, 44, 185–191. [Google Scholar] [CrossRef]
- Rodzen, J.A.; May, B. Inheritance of Microsatellite Loci in the White Sturgeon (Acipenser transmontanus). Genome 2002, 45, 1064–1076. [Google Scholar] [CrossRef]
- Schreier, A.D.; Van Eenennaam, J.P.; Anders, P.; Young, S.; Crossman, J. Spontaneous Autopolyploidy in the Acipenseriformes, with Recommendations for Management. Rev. Fish Biol. Fish. 2021, 31, 159–180. [Google Scholar] [CrossRef]
- Birstein, V.J. Sturgeon Species and Hybrids: Can Hybrids Produce Caviar. Environ. Policy Law 2002, 32, 210–214. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).