Abstract
The tribe Rhodniini is a monophyletic group composed of 24 species grouped in two genera: Rhodnius and Psammolestes. Rhodnius is a paraphyletic genus formed by 21 species. The event of paraphilia is supported by the greater evolutionary proximity of the species of the prolixus groups with the genus Psammolestes than with the other groups of Rhodnius. Based on this phenomenon, it was proposed the alteration of the genus of the species of Psammolestes to Rhodnius. Thus, we performed experimental crosses between Psammolestes spp. and R. neglectus to assess the degree of reproductive compatibility between Psammolestes and Rhodnius. Hybrids were not obtained for intergeneric crosses. Based on this, we have demonstrated that Rhodnius and Psammolestes have reproductive incompatibility. These data are important to confirm the validity of the specific status of Psammolestes, based on the biological concept of the species.
1. Introduction
Chagas disease is a neglected disease caused by the protozoan Trypanosoma cruzi (Chagas, 1909) (Kinetoplastida, Trypanosomatidae) which affects about 8 million people worldwide [1,2]. The main form of transmission of the protozoan is through hematophagous insects known as triatomines (Hemiptera, Triatominae) [1,2]. Currently 157 species of triatomines are known, divided in 18 genera and five tribes [3,4,5], being all considered as potential Chagas disease vectors.
The tribe Rhodniini Pinto, 1926 is a monophyletic group composed of 24 species grouped in two genera: Rhodnius Stål, 1859 with long thin legs and a long head, living mainly in palm trees, and Psammolestes Bergroth, 1911 having a short head, strong legs, wide femora, a very wide rostrum (the widest in all the subfamily) and living in nests of birds [6]. However, there are several reports of these genera living in sympatry in natural conditions, as P. tertius Lent & Jurberg, 1965 living in sympathy with R. neglectus Lent, 1954 in nests of Phacellodomus ruber (Vieillot, 1817) (Passeriformes, Furnariidae) in the federal district, Brazil [7] and P. arthuri (Pinto, 1926) found in sympatry with R. prolixus Stål, 1859 on bird nests in Anzoátegui state, Venezuela [P. rufifrons (Wied-Neuwied, 1821) and Icterus nigrogularis (Hahn, 1819) (Passeriformes, Icteridae)] [8].
Rhodnius is a paraphyletic genus formed by 21 species divided into three groups: pallescens, pictipes and prolixus groups [3,9]. The event of paraphilia is supported by the greater evolutionary proximity of the species of the prolixus groups with the genus Psammolestes than with the other groups of Rhodnius [10,11,12]. Based on this phenomenon, Hypsa et al. [13] proposed the alteration of the genus of the species of Psammolestes to Rhodnius.
Although the change in the generic status of Psammolestes was suggested by these molecular studies [13] (based on the phylogenetic species concept [14]), from the morphological and ecological divergences [6] the genus Psammolestes was kept as valid (considering the phenetic and ecological species concepts [15]). However, recently Fileé et al. [12], using more complex phylogenetic analyzes with mitochondrial and nuclear data (phylogenetic species concept [14]), again recommended the reclassification of species from Psammolestes to Rhodnius.
Thus, considering that studies of hybridization can help to understand the taxonomy and systematics of species (based on the biological species concept [16,17]), can be used to analyze the isolating mechanisms that limit gene flow between species, as well as experimental hybridization can be employed to establish the role of natural hybridization in generating new genetic variants (that may lead to adaptive evolution and/or in founding new evolutionary lineages) [18,19], we performed experimental crosses between Psammolestes spp. and R. neglectus to assess the degree of intergeneric reproductive compatibility between Psammolestes and Rhodnius.
2. Materials and Methods
In order to evaluate the genetic and reproductive compatibility [16] between the genus of the Rhodniini tribe, reciprocal crossing experiments were conducted between R. neglectus (origin: Brazil, São Paulo, Jaboticabal, second generation) and P. coreodes (Bergroth, 1911) (origin: Brazil, Mato Grosso do Sul, Corumbá, second generation) and between R. neglectus and P. tertius (origin: Brazil, Bahia, Castro Alves, third generation). The choice of R. neglectus among the species of the prolixus groups was based on the phylogenetic proximity of the species with Psammolestes spp. [10]. The crossing experiments were conducted in the Triatominae insectary of the School of Pharmaceutical Sciences, São Paulo State University (UNESP), Araraquara, São Paulo, Brazil, in according to the experiments of Neves et al. [20] and Ravazi et al. [21]: the insects were sexed as 5th instar nymphs [22], and males and females were kept separately until they reached the adult stage in order to cross adult virgins [23]. For the crosses, 3 couples from each set were placed in plastic jars (5 cm in diameter ×10 cm in height) and kept at room temperature. Furthermore, intragenerics crosses were also performed for control. Weekly, the triatomines were fed on duck blood, the eggs were collected throughout the female’s oviposition periods (as this period may vary between species, for standardization, we set the “female’s oviposition periods” as “four months after the first oviposition”) and the egg fertility rate was calculated.
3. Results and Discussion
Hybrids were not obtained for intergeneric crosses between Rhodnius and Psammolestes (Table 1, Figure 1B [24]). On the other hand, intrageneric crosses between species of Rhodnius (R. prolixus x R. neglectus (Figure 1A [24]), R. prolixus x R. robustus Larrousse, 1927 and R. pictipes Stål, 1872 x R. prolixus [25]) and between species of Psammolestes (P. coreodes x P. tertius (Figure 1C [24]) [21]) resulted in the formation of hybrids (which were made unfeasible by post-zygotic barriers) [21,25].

Table 1.
Results of the experimental crosses performed between Rhodnius and Psammolestes.
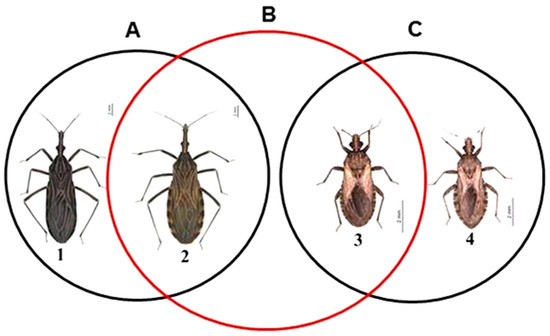
Figure 1.
Scheme representing the intrageneric (A,C) and intergeneric (B) crosses of the Rhodniini tribe. Note that the black circles represent reproductive compatibility and hybrid formation between Rhodnius spp. (A) and between Psammolestes spp. (C) and the red circle demonstrates reproductive incompatibility and absence of hybrid formation between the Rhodnius and Psammolestes genera (B). 1: R. prolixus, 2: R. neglectus, 3: P. tertius and 4: P. coreodes. Bar: 2 mm.
Events that result in pre-zygotic reproductive isolation in Triatominae are more common between species with different chromosome numbers (Triatoma brasiliensis brasiliensis Neiva, 1911 x T. melanocephala Neiva and Pinto, 1923, and T. b. brasiliensis x T. vitticeps (Stål, 1859) [20]), between species that are very distant from the phylogenetic point of view ([26]), or between species of different genera (T. b. brasiliensis x Panstrongylus tibiamaculatus (Pinto, 1926), T. infestans (Klug, 1834) x R. neglectus, T. infestans x R. prolixus, T. sordida (Stål, 1859) x R. prolixus and T. sordida x R. neglectus [20,26]). Recently, Cesaretto et al. [27] crossed Meccus Stål, 1859 and Triatoma Laporte, 1832 species and, due to the absence of a pre-zygotic barrier, confirmed the synonymization of Meccus with Triatoma.
The chromosome evolution of the Rhodniini tribe was quite homogenous from a numerical and structural point of view and resulted in several identical characteristics for most Rhodnius and Psammolestes species: karyotype 2n = 22, sex determination system (XY), heterochromatin pattern (C-bands restricted to the Y sex chromosome for most species}, AT and CG base pair composition (X sex chromosome CMA and DAPI and Y DAPI and CMA for most species) and arrangement and location of the 45S rDNA probes (markings restricted to sex chromosomes) [27,28,29,30]. These similar chromosomal characteristics, together with the phylogenetic relationship of the species of both genera [9,10,11,12,13], could result in the formation of hybrids. However, our results demonstrate that there is reproductive barrier between Rhodnius and Psammolestes (Table 1), confirming the generic status of Psammolestes.
We suggest that after the divergence of the Rhodnius and Psammolestes genera from the common ancestor, genotypic modifications (at the genomic level, which did not lead to changes in the numerical and structural composition of the chromosomes [27,28,29,30]) and phenotypic [6] resulted in pre-zygotic reproductive isolation. Although the genitalia of Rhodnius and Psammolestes show differences (Rhodnius: i. pygophore with median process pointed, blunt, bifurcate or subrectangular; ii. articulatory apparatus with basal bridge well developed; iii. basal plate struts transformed into two short, basally connected convolute structures and iv. genitalia of female truncate, eighth and ninth tergites subvertical; Psammolestes: i. pygophore with short median spinelike process, pointed or rounded apically; ii. articulatory apparatus with basal plate bridge well developed; iii. pedicel about as long as wide; iv. basal plate struts transformed into short curved or convoluted structures and v. genitalia of female truncate, last tergites subvertical [6]), intergeneric copulations were observed (suggesting, therefore, the possible absence of pre-zygotic reproductive isolation due to mechanical incompatibility). These observations are of great taxonomic and evolutionary importance, since it supports the particularities observed between the genera (since there is no intergeneric gene flow) and, above all, it corroborates the generic status of Psammolestes questioned by Hypsa et al. [13] and Fileé et al. [12] based on the biological species concept [16,17].
4. Conclusions
Based on this, we have demonstrated that Rhodnius and Psammolestes have reproductive incompatibility. These data, together with the morphological and ecological data [6] are important to confirm the validity of the specific status of Psammolestes, based on the biological, phenotypic and ecological concepts of the species.
Author Contributions
Conceptualization, A.R., J.d.O., and K.C.C.A.; methodology, A.R., J.d.O., F.F.M., Y.V.d.R., A.B.B.d.O., and K.C.C.A.; formal analysis, A.R., J.d.O., F.F.M., Y.V.d.R., A.B.B.d.O., C.G., M.T.V.d.A.-O., J.A.d.R. and K.C.C.A.; investigation, A.R., J.d.O., F.F.M., Y.V.d.R., A.B.B.d.O., C.G., M.T.V.d.A.-O., J.A.d.R. and K.C.C.A.; resources, A.R., F.F.M., Y.V.d.R., A.B.B.d.O., C.G., M.T.V.d.A.-O., J.A.d.R. and K.C.C.A.; writing—original draft preparation, A.R. and K.C.C.A.; writing—review and editing, A.R., J.d.O., F.F.M., Y.V.d.R., A.B.B.d.O., C.G., M.T.V.d.A.-O., J.A.d.R. and K.C.C.A.; supervision, K.C.C.A., J.d.O. and M.T.V.d.A.-O.; project administration, K.C.C.A., and M.T.V.d.A.-O.; funding acquisition, K.C.C.A., C.G., J.A.d.R., and M.T.V.d.A.-O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Coordination for the Improvement of Higher Education Personnel, Brazil (CAPES)—Finance Code 001, Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Carlos Chagas Filho Research Foundation of the State of Rio de Janeiro (FAPERJ).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data are within the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization. Available online: https://www.who.int/health-topics/chagas-disease#tab=tab_1 (accessed on 23 March 2022).
- Pan American Health Organization. Available online: https://www.paho.org/en/topics/chagas-disease (accessed on 23 March 2022).
- Alevi, K.C.C.; Oliveira, J.; Rocha, D.S.; Galvão, C. Trends in Taxonomy of Chagas Disease Vectors (Hemiptera, Reduviidae, Triatominae): From Linnaean to Integrative Taxonomy. Pathogens 2021, 10, 1627. [Google Scholar] [CrossRef] [PubMed]
- Oliveira Correia, J.P.S.; Gil-Santana, H.R.; Dale, C.; Galvão, C. Triatoma guazu Lent and Wygodzinsky Is a Junior Synonym of Triatoma williami Galvão, Souza and Lima. Insects 2022, 13, 591. [Google Scholar] [CrossRef] [PubMed]
- Gil-Santana, H.R.; Chavez, T.; Pita, S.; Panzera, F.; Galvão, C. Panstrongylus noireaui, a remarkable new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. ZooKeys 2022, 1104, 203–225. [Google Scholar] [CrossRef]
- Lent, H.; Wygodzinsky, P. Revision of the Triatominae (Hemiptera: Reduviidae) and their significance as vector of Chagas’s disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 123–520. [Google Scholar]
- Gurgel-Gonçalves, R.; Duarte, M.A.; Dias, R.E.; Torre, P.A.R.; Romaña, C.A.; Cuba-Cuba, C.A. Spatial distribution of Triatominae populations (Hemiptera: Reduviidae) in Mauritia flexuosa palm trees in Federal District of Brazil. Rev. Soc. Bras. Med. Trop. 2004, 37, 241–247. [Google Scholar] [CrossRef]
- Cruz-Guzmán, P.; Morocoima, A.; Chique, J.; RamonisQuintero, J.; Uzcátegui, M.; Carrasco, H. Psammolestes arthuri naturally infected with Trypanosoma cruzi found in sympatry with Rhodnius prolixus and Triatoma maculata on bird nests in Anzoátegui State, Venezuela. Saber 2014, 26, 428–440. [Google Scholar]
- Justi, S.; Galvão, C. The evolutionary origin of diversity in Chagas disease vectors. Trends Parasitol. 2017, 33, 42–52. [Google Scholar] [CrossRef]
- Justi, S.A.; Galvão, C.; Schrago, C.G. Geological changes of the Americas and their influence on the diversification of the Neotropical kissing bugs (Hemiptera: Reduviidae: Triatominae). PLoS Negl. Trop. Dis. 2016, 10, e0004527. [Google Scholar] [CrossRef]
- de Paula, A.S.; Barreto, C.; Telmo, M.C.M.; Diotaiuti, L.; Galvão, C. Historical biogeography and the evolution of hematophagy in Rhodniini (Heteroptera: Reduviidae: Triatominae). Front. Ecol. Evol. 2021, 9, 660151. [Google Scholar] [CrossRef]
- Filée, J.; Merle, M.; Bastide, H.; Mougel, F.; Bérenger, J.M.; Folly-Ramos, E.; Almeida, C.E.; Harry, M. Phylogenomics for Chagas Disease Vectors of the Rhodnius Genus (Hemiptera, Triatominae): What We Learn From Mito-Nuclear Conflicts and Recommendations. Front. Ecol. Evol. 2022, 9, 750317. [Google Scholar] [CrossRef]
- Hypsa, V.; Tietz, D.; Zrzavy, J.; Rego, R.O.; Galvão, C.; Jurberg, J. Phylogeny and biogeography of Triatominae (Hemiptera, Reduviidae): Molecular evidence of a New World origin of the asiatic clade. Mol. Phylog. Evol. 2012, 23, 447–457. [Google Scholar] [CrossRef]
- Cracraft, J. Species Concepts and Speciation Analysis. Curr. Ornithol. 1983, 1, 159–187. [Google Scholar]
- de Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, E. Populações, Espécies e Evolução, 1st ed.; Editora Nacional: São Paulo, Brasil, 1963; 485p. [Google Scholar]
- Mayr, E. Populations, Species, and Evolution; Harvard University Press: Cambridge, MA, USA, 1970; 453p. [Google Scholar]
- Arnold, M.L. Natural Hybridization and Evolution; Oxford University Press: Oxford, MS, USA, 1997; 215p. [Google Scholar]
- Pérez, R.; Hernández, M.; Quintero, O.; Scovortzoff, E.; Canale, D.; Méndez, L.; Cohanoff, C.; Martino, M.; Panzera, F. Cytogenetic analysis of experimental hybrids in species of Triatominae (Hemiptera-Reduviidae). Genetica 2005, 125, 261–270. [Google Scholar] [CrossRef]
- Neves, S.J.M.; Sousa, P.S.; Oliveira, J.; Ravazi, A.; Madeira, F.F.; Reis, Y.V. Prezygotic isolation confirms the exclusion of Triatoma melanocephala, T. vitticeps and T. tibiamaculata of the T. brasiliensis subcomplex (Hemiptera, Triatominae). Infect. Genet. Evol. 2020, 79, 104149. [Google Scholar] [CrossRef]
- Ravazi, A.; Oliveira, J.; Campos, F.F.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Galvão, C.; Alevi, K.C.C. Trends in evolution of the Rhodniini tribe (Hemiptera, Triatominae): Experimental crosses between Psammolestes tertius Lent & Jurberg, 1965 and P. coreodes Bergroth, 1911 and analysis of the reproductive isolating mechanisms. Parasit. Vec. 2021, 14, 350. [Google Scholar]
- Rosa, J.A.; Barata, J.M.S.; Barelli, N.; Santos, J.L.F.; Belda Neto, F.M. Sexual distinction between 5th instar nymphs of six species (Hemiptera: Reduviidae). Mem. Inst. Oswaldo Cruz 1992, 87, 257–264. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Grant-Guillén, Y.; Delgadillo-Aceves, I.N.; Zumaya-Estrada, F.A.; Rocha-Chávez, G.; Salazar-Schettino, P.M.; Alejandre-Aguilar, R.; Villalobos, G.; Nogueda-Torres, B. Biological and genetic aspects of crosses between phylogenetically close species of Mexican Triatomines (Hemiptera: Reduviidae). J. Med. Entomol. 2011, 48, 705–707. [Google Scholar] [CrossRef]
- Carcavallo, R.U.; Girón, I.G.; Jurberg, J.; Galvao, C.; Noireau, F.; Canale, D. Mutations, Hybrids and Teratologies. In Atlas of Chagas Disease Vectors in the Americas; Carcavallo, R.U., Girón, I.G., Jurberg, J., Lent, H., Eds.; Editora FIOCRUZ: Rio de Janeiro, Brazil, 1998; pp. 515–536, Coleção de Triatominae da FCFAR–UNESP Araraquara; Available online: https://www2.fcfar.unesp.br/#!/triatominae (accessed on 9 July 2022).
- Perlowagora-Szumlewics, A.; Correia, M.V. Induction of male sterility manipulation of genetic mechanisms present in vector species of Chagas disease (remarks on integrating sterile-male release with insecticidal control measures against vectors of Chagas disease). Rev. Inst. Med. Trop São Paulo 1972, 14, 360–371. [Google Scholar]
- Cesaretto, N.R.; Oliveira, J.; Ravazi, A.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Vicente, R.D.; Cristal, D.C.; Galvão, C.; Azeredo-Oliveira, M.T.V.; et al. Trends in taxonomy of Triatomini (Hemiptera, Reduviidae, Triatominae): Reproductive compatibility reinforces the synonymization of Meccus Stål, 1859 with Triatoma Laporte, 1832. Parasit Vect. 2021, 14, 340. [Google Scholar] [CrossRef]
- Alevi, K.C.C.; Moreira, F.F.F.; Jurberg, J.; Azeredo-Oliveira, M.T.V. Description of the diploid chromosome set of Triatoma pintodiasi (Hemiptera, Triatominae). Genet. Mol. Res. 2015, 15, gmr.15026343. [Google Scholar] [CrossRef]
- Alevi, K.C.C.; Oliveira, J.; Rosa, J.A.; Azeredo-Oliveira, M.T.V. Karyotype Evolution of Chagas Disease Vectors (Hemiptera, Triatominae). Am. J. Trop Med. Hyg. 2018, 99, 87–89. [Google Scholar] [CrossRef]
- Pita, S.; Panzera, F.; Ferrandis, I.; Galvão, C.; Gómez-Palacio, A.; Panzera, Y. Chromosomal divergence and evolutionary inferences in Rhodniini based on the chromosomal location of ribosomal genes. Mem. Inst. Oswaldo Cruz. 2013, 108, 376–382. [Google Scholar] [CrossRef]
- Pita, S.; Lorite, P.; Cuadrado, A.; Panzera, Y.; Oliveira, J.; Alevi, K.C.C.; Rosa, J.A.; Freitas, S.P.C.; Gómez-Palacio, A.; Solari, A.; et al. High chromosomal mobility of ribosomal clusters in holocentric chromosomes of Triatominae, vectors of Chagas disease (Hemiptera-Reduviidae). Med. Vet. Entomol. 2022, 36, 66–80. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).