
Effects of Mustard Invasions on Soil Microbial Abundances and Fungal Assemblages in Southern California


 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Collection and Processing
2.3. Greenhouse Experiment
2.4. Analyses
2.4.1. Soil Abiotic and Biotic Properties
2.4.2. Fungal Assemblage Analyses
2.4.3. Germination Analyses
3. Results
3.1. Soil Properties
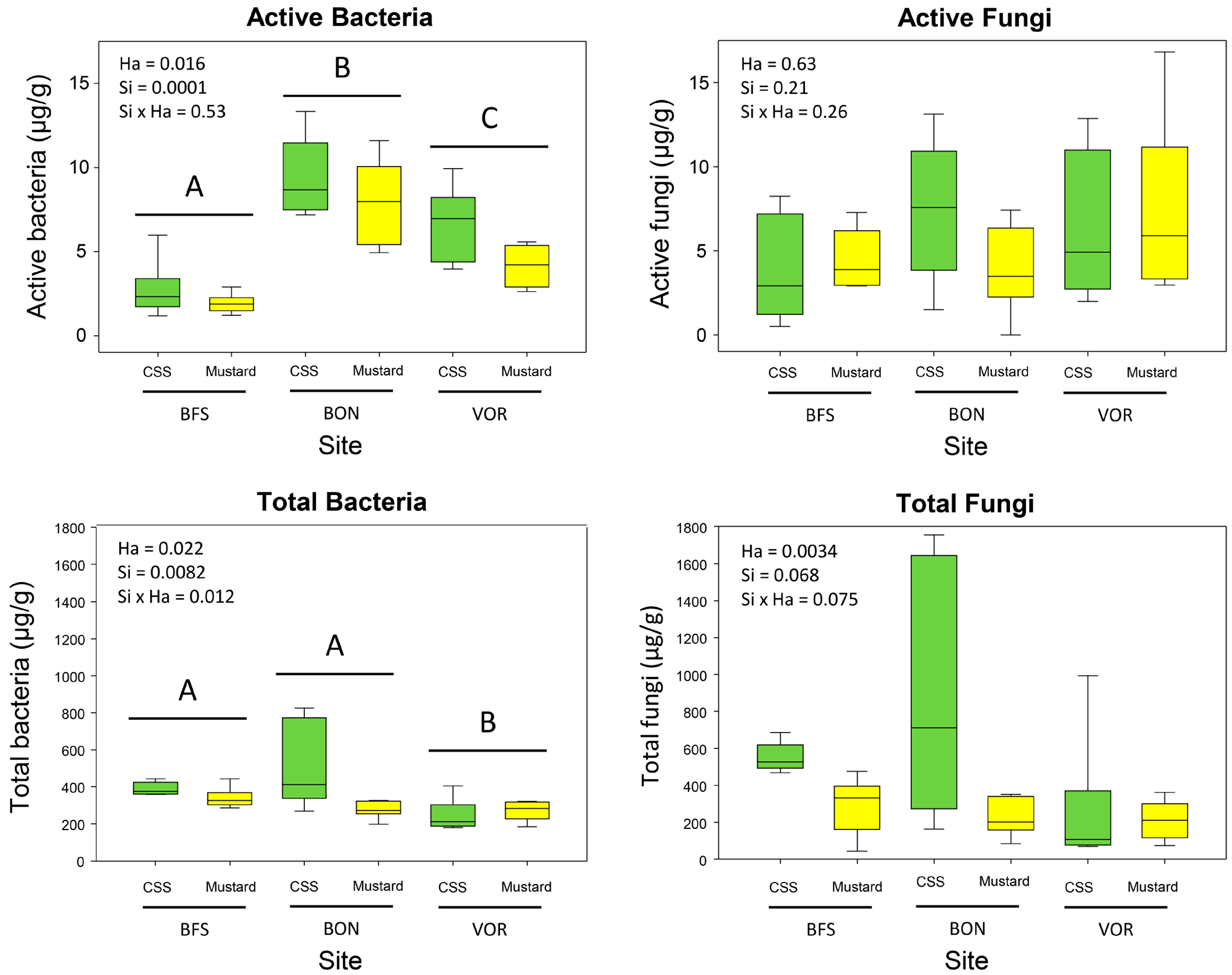
3.2. Microbial Abundances
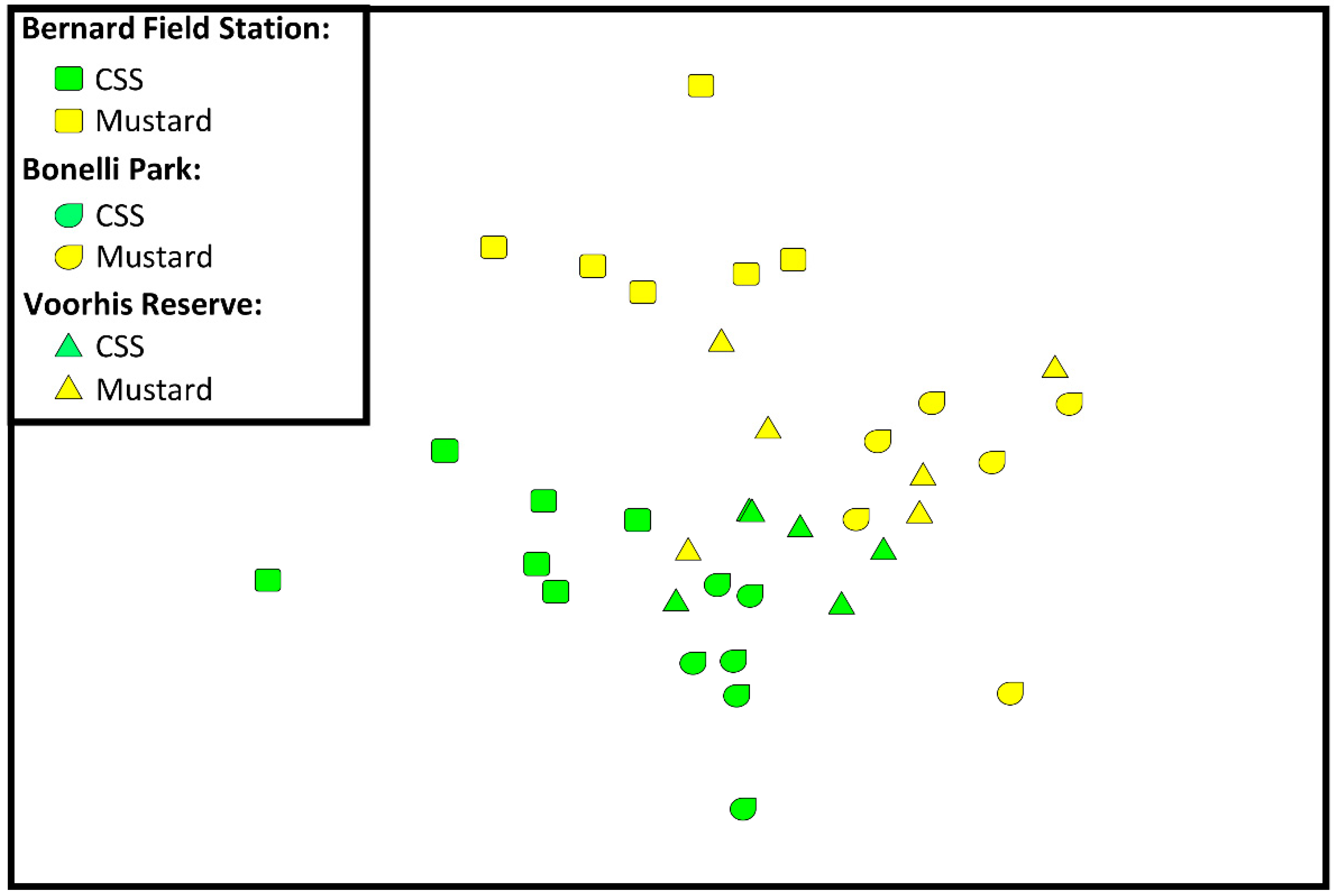
3.3. Fungal Assemblages
3.4. Seedling Germination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buerdsell, S.L.; Milligan, B.G.; Lehnhoff, E.A. Invasive plant benefits a native plant through plant-soil feedback but remains the superior competitor. NeoBiota 2021, 64, 119–136. [Google Scholar] [CrossRef]
- Caspi, T.; Estrada, L.; Dowling, A.V.; Su, E.; Leshchinskiy, M.; Cavalcanti, A.R.O.; Crane, E.J.; Robins, C.R.; Meyer, W.M. Carbon and nitrogen in the topsoils of inceptisols and mollisols under native sage scrub and non-native grasslands in Southern California. Geoderma Reg. 2018, 14, e00172. [Google Scholar] [CrossRef]
- Caspi, T.; Hartz, L.A.; Soto Villa, A.E.; Loesberg, J.A.; Robins, C.R.; Meyer, W.M. Impacts of invasive annuals on soil carbon and nitrogen storage in Southern California depend on the identity of the invader. Ecol. Evol. 2019, 9, 4980–4993. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, D.; Casagrande, G.; Carmona-Galindo, V.D. Effects of black mustard allelopathy on the fitness and life history strategies of buffalo gourd in southern california. Bios 2016, 87, 98–103. [Google Scholar] [CrossRef]
- Verbeek, J.D.; Kotanen, P.M. Soil-mediated impacts of an invasive thistle inhibit the recruitment of certain native plants. Oecologia 2019, 190, 619–628. [Google Scholar] [CrossRef]
- Andrews, H.M.; Jenerette, G.D. Exotic grass litter modulates seasonal pulse dynamics of CO2 and N2O, but not NO, in Mediterranean-type coastal sage scrub at the wildland-urban interface. Plant Soil 2020, 456, 339–353. [Google Scholar] [CrossRef]
- Bell, J.K.; Siciliano, S.D.; Lamb, E.G. A Survey of invasive plants on grassland soil microbial communities and ecosystem services. Sci Data 2020, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Eldridge, D.J.; Travers, S.K.; Val, J.; Ding, J.; Wang, J.-T.; Singh, B.K.; Delgado-Baquerizo, M. Experimental evidence of strong relationships between soil microbial communities and plant germination. J. Ecol. 2021, 109, 2488–2498. [Google Scholar] [CrossRef]
- Pickett, B.; Irvine, I.C.; Bullock, E.; Arogyaswamy, K.; Aronson, E. Legacy effects of invasive grass impact soil microbes and native shrub growth. Invasive Plant Sci. Manag. 2019, 12, 22–35. [Google Scholar] [CrossRef]
- Singh, M.; Meyer, W.M. Plant-soil feedback effects on germination and growth of native and non-native species common across Southern California. Diversity 2020, 12, 217. [Google Scholar] [CrossRef]
- Cox, R.D.; Allen, E.B. Composition of soil seed banks in Southern California coastal sage scrub and adjacent exotic grassland. Plant Ecol. 2008, 198, 37–46. [Google Scholar] [CrossRef]
- Dong, H.; Liu, T.; Liu, Z.; Song, Z. Fate of the soil seed bank of giant ragweed and its significance in preventing and controlling its invasion in grasslands. Ecol. Evol. 2020, 10, 4854–4866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Ge, X.; Fu, Z.; Liu, J. Alternanthera Philoxeroides invasion affects the soil seed bank of reed community. Environ. Exp. Bot. 2020, 180, 104196. [Google Scholar] [CrossRef]
- Puritty, C.E.; Esch, E.H.; Castro, S.P.; Ryan, E.M.; Lipson, D.A.; Cleland, E.E. Drought in Southern California coastal sage scrub reduces herbaceous biomass of exotic species more than native species, but exotic growth recovers quickly when drought ends. Plant Ecol. 2019, 220, 151–169. [Google Scholar] [CrossRef]
- Rundel, P.W. Sage Scrub. In Terrestrial Vegetation of California, 3rd ed.; Barbour, M.G., Keeler-Wolf, T., Schoenherr, A.A., Eds.; University of California Press: Oakland, CA, USA, 2007; pp. 208–228. [Google Scholar]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 2017, 67, 534–545. [Google Scholar] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [PubMed]
- Reid, T.S.; Murphy, D.D. Providing a regional context for local conservation action: A natural community conservation plan for the Southern California coastal sage scrub. BioScience 1995, 45 (Suppl. 1), S84–S90. [Google Scholar] [CrossRef]
- Talluto, M.V.; Suding, K.N. Historical change in coastal sage scrub in Southern California, USA in relation to fire frequency and air pollution. Landsc. Ecol. 2008, 23, 803–815. [Google Scholar] [CrossRef]
- Cox, R.D.; Preston, K.L.; Johnson, R.F.; Minnich, R.A.; Allen, E.B. Influence of landscape-scale variables on vegetation conversion to exotic annual grassland in Southern California, USA. Glob. Ecol. Conserv. 2014, 2, 190–203. [Google Scholar] [CrossRef] [Green Version]
- Finks, S.S.; Weihe, C.; Kimball, S.; Allison, S.D.; Martiny, A.C.; Treseder, K.K.; Martiny, J.B.H. Microbial community response to a decade of simulated global changes depends on the plant community. Elem. Sci. Anthr. 2021, 9, 00124. [Google Scholar] [CrossRef]
- Goldstein, L.J.; Suding, K.N. Intra-annual rainfall regime shifts competitive interactions between coastal sage scrub and invasive grasses. Ecology 2014, 95, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.V.; Wren, I.F.; Herman, D.J.; Firestone, M.K. Plant invasion alters nitrogen cycling by modifying the soil nitrifying community. Ecol. Lett. 2005, 8, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Oduor, A.; van Kleunen, M.; Stift, M. Allelopathic effects of native and invasive Brassica nigra do not support the novel-weapons hypothesis. Am. J. Bot. 2020, 107, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Turk, M.A.; Tawaha, A.M. Allelopathic effect of black mustard (Brassica nigra L.) on germination and growth of wild oat (Avena fatua L.). Crop Prot. 2003, 22, 673–677. [Google Scholar] [CrossRef]
- Tawaha, A.M.; Turk, M.A. Allelopathic Effects of black mustard (Brassica nigra) on germination and growth of wild barley (Hordeum spontaneum). J. Agron. Crop Sci. 2003, 189, 298–303. [Google Scholar] [CrossRef]
- Oduor, A.M.O.; Lankau, R.A.; Strauss, S.Y.; Gómez, J.M. Introduced Brassica Nigra populations exhibit greater growth and herbivore resistance but less tolerance than native populations in the native range. New Phytol. 2011, 191, 536–544. [Google Scholar] [CrossRef]
- Rivera-Vega, L.J.; Krosse, S.; de Graaf, R.M.; Garvi, J.; Garvi-Bode, R.D.; van Dam, N.M. Allelopathic effects of glucosinolate breakdown products in Hanza (Boscia senegalensis (Pers.) Lam.) processing waste water. Front. Plant Sci. 2015, 6, 532. [Google Scholar] [CrossRef] [Green Version]
- Poon, G.T.; Maherali, H. Competitive interactions between a nonmycorrhizal invasive plant, Alliaria petiolata, and a suite of mycorrhizal grassland, old field, and forest species. PeerJ 2015, 3, e1090. [Google Scholar] [CrossRef] [Green Version]
- Stinson, K.A.; Campbell, S.A.; Powell, J.R.; Wolfe, B.E.; Callaway, R.M.; Thelen, G.C.; Hallett, S.G.; Prati, D.; Klironomos, J.N. Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms. PLoS Biol. 2006, 4, e140. [Google Scholar] [CrossRef]
- Blossey, B.; Nuzzo, V.; Dávalos, A. Climate and rapid local adaptation as drivers of germination and seed bank dynamics of Alliaria petiolata (Garlic Mustard) in North America. J. Ecol. 2017, 105, 1485–1495. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of allelopathy on plant performance: A meta-analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef] [PubMed]
- Loesberg, J.A.; Meyer, W.M. Granivory in California sage scrub: Implications for common plant invaders and ecosystem conservation. Plant Ecol. 2021, 222, 1089–1100. [Google Scholar] [CrossRef]
- Orrock, J.L.; Witter, M.S.; Reichman, O.J. Apparent competition with an exotic plant reduces native plant establishment. Ecology 2008, 89, 1168–1174. [Google Scholar] [CrossRef]
- Burke, D.J.; Chan, C.R. Effects of the invasive plant Garlic Mustard (Alliaria petiolata) on bacterial communities in a Northern Hardwood Forest soil. Can. J. Microbiol. 2010, 56, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Cox, E.; Cavalcanti, A.R.O.; Crane, E.J.; Meyer, W.M. Soil bacterial assemblage responses to wildfire in low elevation Southern California habitats. PLoS ONE 2022, 17, e0266256. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Wang, J.; Li, D.; Wu, F.; Zhou, X. Rotations with Indian Mustard and Wild Rocket suppressed cucumber Fusarium Wilt disease and changed rhizosphere bacterial communities. Microorganisms 2019, 7, 57. [Google Scholar] [CrossRef] [Green Version]
- Pérez Castro, S.; Cleland, E.E.; Wagner, R.; Sawad, R.A.; Lipson, D.A. Soil microbial responses to drought and exotic plants shift carbon metabolism. ISME J. 2019, 13, 1776–1787. [Google Scholar] [CrossRef]
- Kirkpatrick, J.B.; Hutchinson, C.F. The community composition of Californian coastal sage scrub. Plant Ecol. 1977, 35, 21–33. [Google Scholar] [CrossRef]
- Kirkpatrick, J.B.; Hutchinson, C.F. The environmental relationships of Californian coastal sage scrub and some of its component communities and species. J. Biogeogr. 1980, 7, 23–38. [Google Scholar] [CrossRef]
- Paysen, T.E.; Derby, J.A.; Black, H.; Bleich, V.C.; Mincks, J.W. A Vegetation Classification System Applied to Southern California; General Techical Report PSW-45; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Albany, CA, USA, 1980; Volume 45, 33p. [Google Scholar]
- Mooney, H.; Zavaleta, E. (Eds.) Ecosystems of California; University of California Press: Oakland, CA, USA, 2016. [Google Scholar]
- Schnürer, J.; Rosswall, T. Fluorescein Diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babiuk, L.A.; Paul, E.A. The use of fluorescein isothiocyanate in the determination of the bacterial biomass of grassland soil. Can. J. Microbiol. 1970, 16, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, J.A.; Paul, E.A. Conversion of biovolume measurements of soil organisms, grown under various moisture tensions, to biomass and their nutrient content. Appl. Environ. Microbiol. 1979, 37, 686–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R.; Gorley, R.N. PRIMER V6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Bolyen, D.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Rivers, A.R.; Weber, K.C.; Gardner, T.G.; Liu, S.; Armstrong, S.D. ITSxpress: Software to rapidly trim internally transcribed spacer sequences with quality scores for marker gene analysis. F1000Research 2018, 7, 1418. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome. 2018, 6, 90. [Google Scholar] [CrossRef]
- Abarenkov, K.; Zirk, A.; Põldmaa, K.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Adojaan, K.; Kõljalg, U. Third-party annotations: Linking PlutoF platform and the ELIXIR Contextual Data ClearingHouse for the reporting of source material annotation gaps and inaccuracies. Biodivers. Inf. Sci. Stand. 2021, 5, e74249. [Google Scholar] [CrossRef]
- Fouquier, J.; Rideout, J.R.; Bolyen, E.; Chase, J.; Shiffer, A.; McDonald, D.; Knight, R.; Caporaso, J.G.; Kelley, S.T. Ghost-tree: Creating hybrid-gene phylogenetic trees for diversity analyses. Microbiome 2016, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef]
- Aime, M.C.; Toome, M.; McLaughlin, D.J. Pucciniomycotina. In Systematics and Evolution: Part A; McLaughlin, D.J., Spatafora, J.W., Eds.; The Mycota; Springer: Berlin/Heidelberg, Germany, 2014; pp. 271–294. [Google Scholar]
- Francioli, D.; Lentendu, G.; Lewin, S.; Kolb, S. DNA Metabarcoding for the characterization of terrestrial microbiota—Pitfalls and solutions. Microorganisms 2021, 9, 361. [Google Scholar] [CrossRef] [PubMed]
- Öpik, M.; Davison, J.; Moora, M.; Zobel, M. DNA-based detection and identification of Glomeromycota: The virtual taxonomy of environmental sequences. Botany 2014, 92, 135–147. [Google Scholar] [CrossRef]
- Yuste, J.C.; Fernandez-Gonzalez, A.J.; Fernandez-Lopez, M.; Ogaya, R.; Penuelas, J.; Lloret, F. Functional diversification within bacterial lineages promotes wide functional overlapping between taxonomic groups in a Mediterranean forest soil. FEMS Microbiol. Ecol. 2014, 90, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Pakpour, S.; Klironomos, J. The invasive plant, Brassica nigra, degrades local mycorrhizas across a wide geographical landscape. R. Soc. Open Sci. 2015, 2, 150300. [Google Scholar] [CrossRef] [Green Version]
- Schreiner, R.P.; Koide, R.T. Mustards, mustard oils and mycorrhizas. New Phytol. 1993, 123, 107–113. [Google Scholar] [CrossRef]
- Bozzolo, F.H.; Lipson, D.A. Differential responses of native and exotic coastal sage scrub plant species to N additions and the soil microbial community. Plant Soil 2013, 371, 37–51. [Google Scholar] [CrossRef]
- Miller, E.C.; Perron, G.G.; Collins, C.D. Plant-driven changes in soil microbial communities influence seed germination through negative feedbacks. Ecol. Evol. 2019, 9, 9298–9311. [Google Scholar] [CrossRef] [Green Version]
- Yelenik, S.G.; Levine, J.M. The role of Plant–Soil Feedbacks in driving native-species recovery. Ecology 2011, 92, 66–74. [Google Scholar] [CrossRef]
- Allison, V.J.; Miller, R.M.; Jastrow, J.D.; Matamala, R.; Zak, D.R. Changes in soil microbial community structure in a tallgrass prairie chronosequence. Soil Sci. Soc. Am. J. 2005, 69, 1412–1421. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Leff, J.W.; Adams, B.J.; Nielsen, U.N.; Bates, S.T.; Lauber, C.L.; Owens, S.; Gilbert, J.A.; Wall, D.H.; Caporaso, J.G. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proc. Natl. Acad. Sci. USA 2012, 109, 21390–21395. [Google Scholar] [CrossRef]
- Manning, P. The impact of nitrogen enrichment on ecosystems and their services. In Soil Ecology and Ecosystem Services; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Wardle, D.A. The influence of biotic interactions on soil biodiversity. Ecol. Lett. 2006, 9, 870–886. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Site | Sediment Size | |||||
|---|---|---|---|---|---|---|
| Habitat | pH † | SOM (%) | Soil Moisture (%) † | Sand (%) | Silt (%) | Clay (%) |
| Bernard Field Station | ||||||
| CSS | 5.2 ± 0.33 | 3.0 ± 0.65 | 0.4 ± 0.13 | 81 | 14 | 6 |
| Mustard | 6.1 ± 0.22 | 3.3 ± 0.99 | 0.4 ± 0.25 | 78 | 15 | 7 |
| Bonelli Park | ||||||
| CSS | 5.9 ± 0.49 | 12 ± 3.5 | 4.2 ± 0.86 | 45 | 26 | 29 |
| Mustard | 6.6 ± 0.44 | 9 ± 1.1 | 2.4 ± 0.46 | 78 | 11 | 11 |
| Voorhis Reserve | ||||||
| CSS | 6.5 ± 0.53 | 5.0 ± 0.95 a | 1.5 ± 0.29 | 72 | 15 | 13 |
| Mustard | 6.5 ± 0.76 | 9 ± 1.99 b | 1.7 ± 0.63 | 65 | 18 | 18 |
| Function | Bernard Field Station | Bonelli Park | Voorhis Reserve | Habitat | Site | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Taxon | CSS | Mustard | CSS | Mustard | CSS | Mustard | F1 | p | F2 | p |
| Ascomycota | 80 ± 11 | 50 ± 24 | 60 ± 21 | 40 ± 23 | 60 ± 18 | 70 ± 15 | 7.1 | 0.01 | 1.5 | 0.22 |
| Basidiomycota | 4 ± 3.2 | 30 ± 29 | 10 ± 27 | 20 ± 31 | 10 ± 13 | 10 ± 13 | 1.6 | 0.22 | 0.07 | 0.93 |
| Plant Pathogens | ||||||||||
| Pucciniomycotina | 0.3 ± 0.31 | 0.12 ± 0.098 | 0.3 ± 0.39 | 0.1 ± 0.29 | 0.8 ± 0.61 | 0.2 ± 0.13 | 6.6 | 0.01 | 2.4 | 0.10 |
| EMF | ||||||||||
| Agaricales | 0.1 ± 0.10 | 20 ± 30 | 10 ± 16 | 10 ± 32 | 0 ± 5.5 | 10 ± 14 | 2.8 | 0.11 | 0.12 | 0.89 |
| Boletales | 0 | 0.003 ± 0.0082 | 0.3 ± 0.56 | 0.02 ± 0.047 | 0 | 0 | 1.3 | 0.28 | 2.0 | 0.04 |
| Cantharellales | 1 ± 3.3 | 7 ± 6.7 | 0.1 ± 0.21 | 2 ± 4.7 | 5 ± 8.3 | 0.9 ± 0.93 | 0.46 | 0.51 | 1.1 | 0.35 |
| Pezizales | 0 | 0.4 ± 0.37 | 0.1 ± 0.22 | 4 ± 6.0 | 0.01 ± 0.016 | 1 ± 1.6 | 4.1 | 0.02 | 1.3 | 0.29 |
| Helotiales | 3 ± 6.0 | 1.1 ± 0.90 | 2 ± 2.6 | 0.5 ± 0.48 | 0.9 ± 0.73 | 1 ± 1.2 | 0.92 | 0.42 | 0.42 | 0.79 |
| AMF | ||||||||||
| Glomeromycota | 0.02 ± 0.053 | 0.05 ± 0.090 | 0 | 0 | 0 | 0 ± 0.10 | 1.5 | 0.26 | 1.1 | 0.36 |
| Plant Identification | Bernard Field Station | Bonelli Park | Voorhis Reserve | |||
| CSS | Mustard | CSS | Mustard | CSS | Mustard | |
| Plant Species Totals | ||||||
| Brassicaceaespp. 1 | 1 | 21 | 3 | 80 | 1 | 60 |
| Nicotiana glauca1 | 0 | 6 | 0 | 0 | 1 | 24 |
| Non-native grasses (Poaceae spp.) 1 | 0 | 0 | 0 | 0 | 4 | 2 |
| Melilotus officinalis1 | 0 | 0 | 4 | 0 | 0 | 0 |
| Amsinckia intermedia1 | 0 | 2 | 0 | 0 | 0 | 0 |
| Centaurea melitensis1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Marrubium vulgare1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Total invasive plants | 1 | 29 | 8 | 80 | 6 | 87 |
| Pseudognaphalium californicum | 1 | 1 | 1 | 2 | 3 | 1 |
| Artemisia californica | 0 | 0 | 0 | 1 | 1 | 1 |
| Crassula connata | 0 | 1 | 0 | 0 | 0 | 0 |
| Plagiobothrys sp. | 0 | 1 | 0 | 0 | 0 | 0 |
| Total native plants | 1 | 3 | 1 | 3 | 4 | 2 |
| Solanacaea spp. 2 | 0 | 0 | 0 | 0 | 0 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wakefield, Z.R.; Cavalcanti, A.R.O.; Driessen, L.; Jaramillo, A.; Crane, E.J., III; Richetta, G.; Meyer, W.M., III. Effects of Mustard Invasions on Soil Microbial Abundances and Fungal Assemblages in Southern California. Diversity 2023, 15, 50. https://doi.org/10.3390/d15010050
Wakefield ZR, Cavalcanti ARO, Driessen L, Jaramillo A, Crane EJ III, Richetta G, Meyer WM III. Effects of Mustard Invasions on Soil Microbial Abundances and Fungal Assemblages in Southern California. Diversity. 2023; 15(1):50. https://doi.org/10.3390/d15010050
Chicago/Turabian StyleWakefield, Zachary R., André R. O. Cavalcanti, Lucía Driessen, Ana Jaramillo, Edward J. Crane, III, Giuliano Richetta, and Wallace M. Meyer, III. 2023. "Effects of Mustard Invasions on Soil Microbial Abundances and Fungal Assemblages in Southern California" Diversity 15, no. 1: 50. https://doi.org/10.3390/d15010050
APA StyleWakefield, Z. R., Cavalcanti, A. R. O., Driessen, L., Jaramillo, A., Crane, E. J., III, Richetta, G., & Meyer, W. M., III. (2023). Effects of Mustard Invasions on Soil Microbial Abundances and Fungal Assemblages in Southern California. Diversity, 15(1), 50. https://doi.org/10.3390/d15010050