

The Effects of Tropical Elevations and Associated Habitat Changes on Firefly (Coleoptera: Lampyridae) Diversity in Malaysia


Abstract
1. Introduction
Malaysian Fireflies across Elevation as a Case Study
2. Materials and Methods
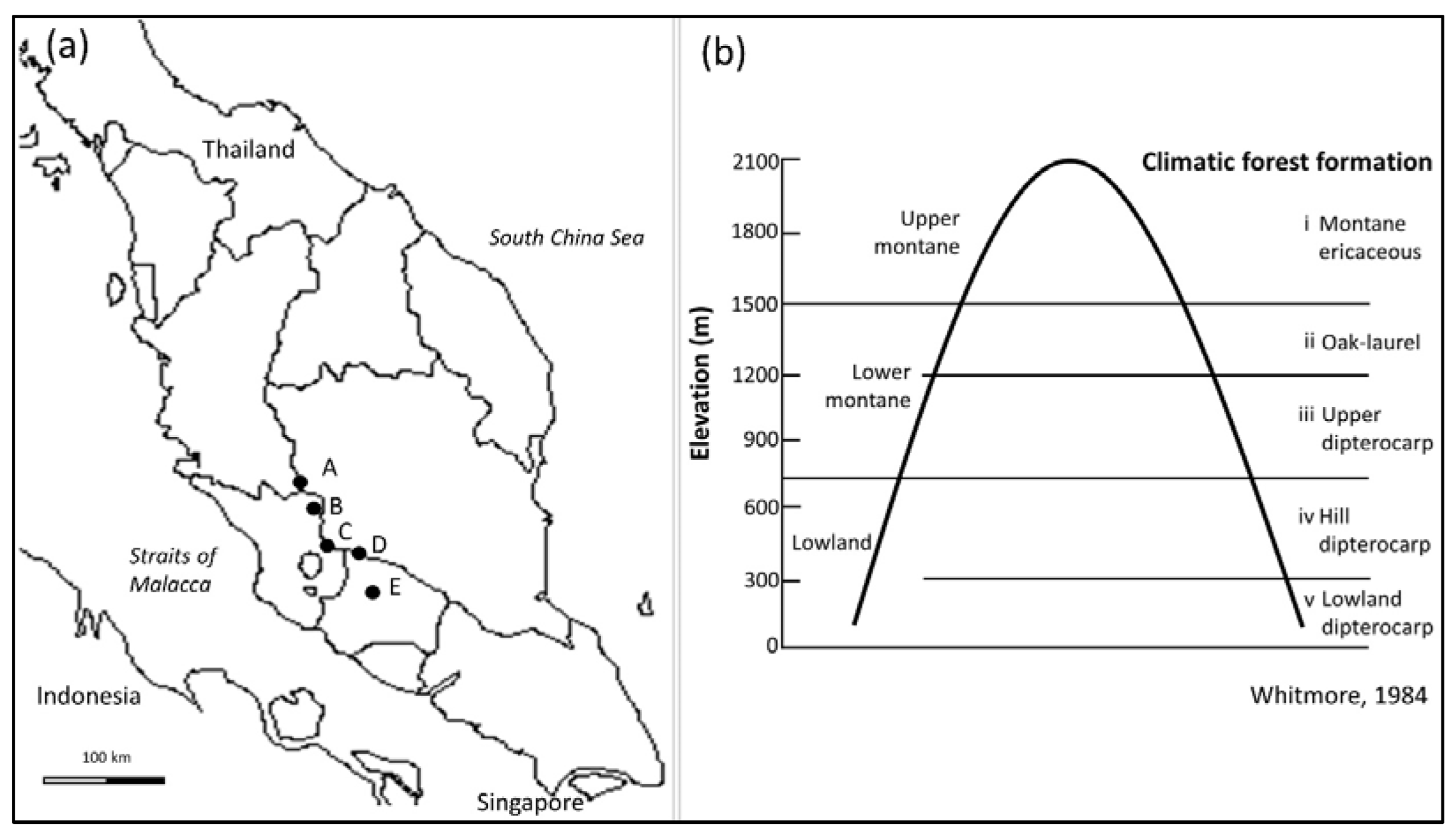
2.1. Study Area
2.2. Firefly Sampling and Identification
2.3. Habitat Characteristics’ Recording
2.4. Data Analysis
2.4.1. Species Beta-Diversity Analysis
2.4.2. Species-Diversity–Habitat Relationships
3. Results
3.1. Sampling Effort and Species Found
3.2. Firefly Elevational Ranges

{kind=link}
{kind=link}
| Elevation Bands, m a.s.l. | Firefly Species Found (Average no.) | Forest Types |
|---|---|---|
| 1900 | - | Mountain ericaceous |
| 1850 | - | |
| 1800 | Drilaster sp. (1.00), Pyrocoelia sp. 1 (1.00) | |
| 1750 | Curtos costipennis (1.00), Drilaster sp. (1.00), Pyrocoelia fumigata (3.00) | |
| 1700 | Pyrocoelia sp. 1 (3.00), Pyrocoelia sp. 3 (3.00) | |
| 1650 | - | |
| 1600 | Curtos obscuricolor (1.00), Pyrocoelia sp. 3 (3.00) | |
| 1550 | - | |
| 1500 | - | Oak-laurel |
| 1450 | - | |
| 1400 | Abscondita berembun * (1.00), Luciola jengai *,† (1.00) | |
| 1350 | - | |
| 1300 | - | |
| 1250 | Curtos obscuricolor (0.33), Curtos costipennis (0.33) | |
| 1200 | Curtos costipennis (0.33), Stenocladius sp. 1 † (0.33) | Upper hill dipterocarp |
| 1150 | Curtos costipennis (0.33) | |
| 1100 | Curtos costipennis (0.33) | |
| 1050 | - | |
| 1000 | Abscondita berembun * (1.20), Stenocladius sp. (0.20) | |
| 950 | Stenocladius sp. 1 † (0.20) | |
| 900 | Abscondita pallescens (0.20) | |
| 850 | Abscondita pallescens (0.40), Diaphanes sp. 1 (0.20), Stenocladius sp. 1 † (0.20) | |
| 800 | Abscondita pallescens (0.20) | Hill dipterocarp |
| 750 | Abscondita pallescens (0.60) | |
| 700 | Abscondita pallescens (0.20) | |
| 650 | Diaphanes sp. 1 (0.20), Pyrocoelia sp. 2 (0.20), | |
| 600 | Diaphanes sp. 2 (0.20), Pygoluciola dunguna * (0.20) | |
| 550 | Abscondita pallescens (0.20), Colophotia brevis (0.20), Curtos sp. 1 (0.20), Stenocladius sp. 2 (0.40) | |
| 500 | Abscondita pallescens (0.20), Colophotia brevis (0.20), Pygoluciola dunguna * (0.20) | |
| 450 | Colophotia brevis (0.40), Stenocladius sp. 2 (0.40) | |
| 400 | Pyrocoelia sp. 2 (0.20) | |
| 350 | Abscondita pallescens (0.20), Luciola jengai *† (0.40), Pygoluciola dunguna * (1.80), Stenocladius sp. 2 (0.20) | |
| 300 | Abscondita pallescens (0.20), Diaphanes sp. 1 (0.20), Diaphanes sp. 2 (0.40), Luciola jengai *† (0.20), Pygoluciola dunguna * (0.80) | Lowland dipterocarp |
| 250 | Colophotia brevis (1.00), Luciola pallidipes (1.33), Stenocladius sp. 1 (1.00), Stenocladius sp. 2 (0.20) | |
| 200 | Colophotia brevis (1.00), Pyrocoelia sp. 2 (1.00), Stenocladius sp. 1 † (3.00) |
3.3. Species Turnover (β-Diversity)—Multivariate Generalised Linear Model Method
3.4. Species Turnover (β-Diversity)—Distance Matrix Methods
3.5. Species-Diversity–Habitat Relationships
4. Discussion
4.1. Firefly Elevational Ranges
4.2. β-Diversity of Fireflies between Mountains and between Forest Types
4.3. Firefly Species Diversity and Habitat Relationships
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lomolino, M.V. Elevation Gradients of Species-Density: Historical and Prospective Views. Glob. Ecol. Biogeogr. 2001, 10, 3–13. [Google Scholar] [CrossRef]
- Guo, Q.; Kelt, D.A.; Sun, Z.; Liu, H.; Hu, L.; Ren, H.; Wen, J. Global Variation in Elevational Diversity Patterns. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, C.; Rapini, A.; Santos Damascena, L.; De Marco Junior, P. The Worrying Future of the Endemic Flora of a Tropical Mountain Range under Climate Change. Flora 2016, 218, 1–10. [Google Scholar] [CrossRef]
- Chaves, A.V.; Freitas, G.H.S.; Vasconcelos, M.F.; Santos, F. Biogeographic Patterns, Origin and Speciation of the Endemic Birds from Eastern Brazilian Mountaintops: A Review. Syst. Biodivers. 2015, 13, 1–16. [Google Scholar] [CrossRef]
- Macedo, M.V.; Monteiro, R.F.; Flinte, V.; Almeida-Neto, M.; Khattar, G.; da Silveira, L.F.L.; Araújo, C.d.O.; Araújo, R.d.O.; Colares, C.; Gomes, C.V.S.; et al. Insect Elevational Specialization in a Tropical Biodiversity Hotspot. Insect Conserv. Divers. 2017, 11, 240–254. [Google Scholar] [CrossRef]
- Freeman, B.G.; Class Freeman, A.M. Rapid Upslope Shifts in New Guinean Birds Illustrate Strong Distributional Responses of Tropical Montane Species to Global Warming. Proc. Natl. Acad. Sci. USA 2014, 111, 4490–4494. [Google Scholar] [CrossRef]
- Molina-Martínez, A.; León-Cortés, J.L.; Regan, H.M.; Lewis, O.T.; Navarrete, D.; Caballero, U.; Luis-Martínez, A. Changes in Butterfly Distributions and Species Assemblages on a Neotropical Mountain Range in Response to Global Warming and Anthropogenic Land Use. Divers. Distrib. 2016, 22, 1085–1098. [Google Scholar] [CrossRef]
- Cusack, D.F.; Karpman, J.; Ashdown, D.; Cao, Q.; Ciochina, M.; Halterman, S.; Lydon, S.; Neupane, A. Global Change Effects on Humid Tropical Forests: Evidence for Biogeochemical and Biodiversity Shifts at an Ecosystem Scale. Rev. Geophys. 2016, 54, 523–610. [Google Scholar] [CrossRef]
- Zhou, X.; Fu, Y.; Zhou, L.; Li, B.; Luo, Y. An Imperative Need for Global Change Research in Tropical Forests. Tree Physiol. 2013, 33, 903–912. [Google Scholar] [CrossRef]
- Menendez, R.; Gonzalez-Megias, A.; Jay-Robert, P.; Marquez-Ferrando, R. Climate Change and Elevational Range Shifts: Evidence from Dung Beetles in Two European Mountain Ranges. Glob. Ecol. Biogeogr. 2014, 23, 646–657. [Google Scholar] [CrossRef]
- García-López, A.; Micó, E.; Galante, E. From Lowlands to Highlands: Searching for Elevational Patterns of Species Richness and Distribution of Scarab Beetles in Costa Rica. Divers. Distrib. 2012, 18, 543–553. [Google Scholar] [CrossRef]
- Sheldon, K.S.; Huey, R.B.; Kaspari, M.; Sanders, N.J. Fifty Years of Mountain Passes: A Perspective on Dan Janzen’s Classic Article. Am. Nat. 2018, 191, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Raxworthy, C.J.; Pearson, R.G.; Rabibisoa, N.; Rakotondrazafy, A.M.; Ramanamanjato, J.B.; Raselimanana, A.P.; Wu, S.; Nussbaum, R.A.; Stone, D.A. Extinction Vulnerability of Tropical Montane Endemism from Warming and Upslope Displacement: A Preliminary Appraisal for the Highest Massif in Madagascar. Glob. Chang. Biol. 2008, 14, 1703–1720. [Google Scholar] [CrossRef]
- Nada, B.; Mohd Salleh, S.; Khirul Faizal, O. A Survey of Fireflies (Coleoptera: Lampyridae) in Fraser’s Hill, Pahang. In Forest R&D: Meeting National and Global Needs, Proceedings of the Conference on Forestry and Forest Products Research 2013, Kuala Lumpur, Malaysia, 12–13 November 2013; Rahim, S., Lim, H.F., Huda Farhana, M.M., Mahmudin, S., Eds.; Forest Research Institute Malaysia: Kuala Lumpur, Malaysia, 2014; pp. 14–19. [Google Scholar]
- Wong, C.H. In the Blink of a Firefly. Malays. Nat. 2014, 26–32. [Google Scholar]
- Chow, A.; Chong, J.H.; Cook, M.; White, D. Vanishing Fireflies: A Citizen-Science Project Promoting Scientific Inquiry and Environmental Stewardship. Sci. Educ. Civ. Engagem. 2014, 6, 23–31. [Google Scholar]
- Bonney, R.; Dickinson, J.L. Overview of Citizen Science. In Citizen Science: Public Participation in Environmental Research; Dickinson, J.L., Bonney, R., Eds.; Cornell University Press: Ithaca, NY, USA, 2012; pp. 19–26. [Google Scholar]
- Xing, Y. Local Environment Attachment and the Possibility of Using Citizen Science Approaches to Measure Firefly Populations in Time and Place. Unpublished Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2012; p. 170. [Google Scholar]
- Viviani, V.R.; Rosa, S.P.; Martins, M.A. Aspisoma Lineatum (Gyllenhal) (Coleoptera: Lampyridae) Firefly: Description of the Immatures, Biological, and Ecological Aspects. Neotrop. Entomol. 2012, 41, 89–94. [Google Scholar] [CrossRef]
- Fu, X.H.; Meyer-Rochow, V.B. Larvae of the Firefly Pyrocoelia Pectoralis (Coleoptera: Lampyridae) as Possible Biological Agents to Control the Land Snail Bradybaena Ravida. Biol. Control 2013, 65, 176–183. [Google Scholar] [CrossRef]
- Fu, X.H.; Ballantyne, L.A.; Lambkin, C.L. The External Larval Morphology of Aquatic and Terrestrial Luciolinae Fireflies (Coleoptera: Lampyridae). Zootaxa 2012, 3405, 1–34. [Google Scholar] [CrossRef]
- Ballantyne, L.A.; Lambkin, C.L.; Luan, X.; Boontop, Y.; Nak-Eiam, S.; Pimpasalee, S.; Silalom, S.; Thancharoen, A. Further Studies on South Eastern Asian Luciolinae: 1. Sclerotia Ballantyne, a New Genus of Fireflies with Back Swimming Larvae 2. Triangulara Pimpasalee, a New Genus from Thailand (Coleoptera: Lampyridae). Zootaxa 2016, 4170, 201–249. [Google Scholar] [CrossRef]
- Fu, X.H.; Meyer-Rochow, V.B. An Investigation into the Morphological and Behavioral Adaptations of the Aquatic Larvae of Aquatica Leii (Coleoptera: Lampyridae) to Prey upon Freshwater Snails That Serve as Intermediate Hosts for the Liver Fluke. Biol. Control 2012, 62, 127–134. [Google Scholar] [CrossRef]
- Jeng, M.-L.; Lai, J.; Yang, P.-S.; Sato, M. Revision of the Genus Diaphanes Motschulsky (Coleoptera, Lampyridae, Lampyrinae) of Taiwan. Jpn. J. Syst. Entomol. 2001, 7, 203–235. [Google Scholar]
- Fu, X.H.; South, A.; Lewis, S.M. Sexual Dimorphism, Mating Systems, and Nuptial Gifts in Two Asian Fireflies (Coleoptera: Lampyridae). J. Insect Physiol. 2012, 58, 1485–1492. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.M.; Schlindwein, M.N.; Viviani, V.R. Survey of Bioluminescent Coleoptera in the Atlantic Rain Forest of Serra Da Paranapiacaba in São Paulo State (Brazil). Biota Neotrop. 2016, 16, e0045. [Google Scholar] [CrossRef]
- Firebaugh, A.; Haynes, K.J. Experimental Tests of Light-Pollution Impacts on Nocturnal Insect Courtship and Dispersal. Oecologia 2016, 182, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Kazama, S.; Matsumoto, S.; Ranjan, S.P.; Hamamoto, H.; Sawamoto, M. Characterization of Firefly Habitat Using a Geographical Information System with Hydrological Simulation. Ecol. Modell. 2007, 209, 392–400. [Google Scholar] [CrossRef]
- Kakehashi, K.; Kuranishi, R.B.; Kamata, N. Estimation of Dispersal Ability Responding to Environmental Conditions: Larval Dispersal of the Flightless Firefly, Luciola Parvula (Coleoptera: Lampyridae). Ecol. Res. 2014, 29, 779–787. [Google Scholar] [CrossRef]
- Smith, B.W. Firefly Diversity in Colombia: Patterns Across a Dynamic Landscape. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2009; p. 123. [Google Scholar]
- Wattanachaiyingcharoen, W.; Nak-Eiam, S.; Phanmuangma, W.; Boonkiaew, S.; Nimlob, N. Species Diversity of Firefly (Coleoptera: Lampyridae) in the Highlands of Northern Thailand. NU Int. J. Sci. 2016, 13, 24–32. [Google Scholar]
- Fu, X.H.; Ballantyne, L.A. Taxonomy and Behaviour of Lucioline Fireflies (Coleoptera: Lampyridae: Luciolinae) with Redefinition and New Species of Pygoluciola Wittmer from Mainland China and Review of Luciola LaPorte. Zootaxa 2008, 1733, 1–44. [Google Scholar] [CrossRef]
- Jeng, M.-L.; Engel, M.S.; Yang, P.-S. Discovery of the Female of Pyrocoelia Prolongata in Taiwan (Coleoptera, Lampyridae). Zookeys 2011, 116, 49–57. [Google Scholar] [CrossRef]
- Jamaludin, S.; Sayang, M.D.; Wan Zawiah, W.Z.; Abdul Aziz, J. Trends in Peninsular Malaysia Rainfall Data during the Southwest Monsoon and Northeast Monsoon Seasons: 1975–2004. Sains Malays. 2010, 39, 533–542. [Google Scholar]
- Whitmore, T.C. Tropical Rain Forest of the Far East; Oxford University Press: Oxford, UK, 1984; p. 352. [Google Scholar]
- Timeanddate.Com. Available online: www.timeanddate.com (accessed on 12 September 2015).
- Ballantyne, L.A.; Lambkin, C.L. Systematics and Phylogenetics of Indo-Pacific Luciolinae Fireflies (Coleoptera: Lampyridae) and the Description of New Genera. Zootaxa 2013, 3653, 1–162. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, L.A.; Fu, X.H.; Lambkin, C.L.; Jeng, M.-L.; Faust, L.F.; Wijekoon, W.M.C.D.; Li, D.; Zhu, T. Studies on South-East Asian Fireflies: Abscondita, a New Genus with Details of Life History, Flashing Patterns and Behaviour of Abs. Chinensis (L.) and Abs. Terminalis (Olivier) (Coleoptera: Lampyridae: Luciolinae). Zootaxa 2013, 3721, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Jeng, M.-L.; Lai, J.; Yang, P.-S.; Sato, M. On the Validity of the Generic Name Pyrocoelia Gorham (Coleoptera, Lampyridae, Lampyrinae) with Review of Taiwanese Species. Jpn. J. Syst. Entomol. 1999, 5, 347–362. [Google Scholar]
- Jeng, M.-L.; Yang, P.-S.; Engel, M.S. The Firefly Genus Vesta in Taiwan (Coleoptera: Lampyridae). J. Kans. Entomol. Soc. 2007, 80, 265–280. [Google Scholar] [CrossRef]
- Kawashima, I.; Satou, F.; Sato, M. The Lampyrid Genus Drilaster (Coleoptera, Lampyridae, Ototretinae) of the Ryukyu Archipelago, Southwest Japan. Jpn. J. Syst. Entomol. 2005, 11, 225–262. [Google Scholar]
- Lemmon, P.E. A Spherical Densiometer for Estimating Forest Overstory Density. For. Sci. 1956, 2, 314–320. [Google Scholar]
- Baselga, A.; Orme, C.D.L. Betapart: An R Package for the Study of Beta Diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, R.B.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package Vegan: Community Ecology Package, Version 2.5-2. R Package. 2018. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 15 June 2018).
- Hsieh, T.C.; Ma, K.H.; Chao, A. Package ‘INEXT’ Version 2.0.15. R Package. 2018. Available online: http://chao.stat.nthu.edu.tw/wordpress/software_download/ (accessed on 30 July 2018).
- Fox, J.; Weisberg, S.; Price, B.; Adler, D.; Bates, D.; Baud-Bovy, G.; Bolker, B.; Ellison, S.; Firth, D.; Friendly, M.; et al. Package Car: Companion to Applied Regression, Version 3.0-0. R Package. 2018. Available online: https://CRAN.R-project.org/package=car (accessed on 1 May 2018).
- Fox, J.; Friendly, M.; Monette, G. Package Heplots: Visualising Hypothesis Tests in Multivariate Linear Models, Version 1.3-5. R Package. 2018. Available online: https://CRAN.R-project.org/package=heplots (accessed on 1 May 2018).
- Ballantyne, L.A.; Lambkin, C.L.; Ho, J.-Z.; Jusoh, W.F.A.; Nada, B.; Nak-Eiam, S.; Thancharoen, A.; Wattanachaiyingcharoen, W.; Yiu, V. The Luciolinae of S. E. Asia and the Australopacific Region: A Revisionary Checklist (Coleoptera: Lampyridae) Including Description of Three New Genera and 13 New Species. Zootaxa 2019, 4687, 1–174. [Google Scholar] [CrossRef]
- Nada, B.; Ballantyne, L.A. A New Species of Pygoluciola Wittmer with Unusual Abdominal Configuration, from Lowland Dipterocarp Forest in Peninsular Malaysia (Coleoptera: Lampyridae: Luciolinae). Zootaxa 2018, 4455, 343–362. [Google Scholar] [CrossRef]
- Saiful, I. Effects of Selective Logging on Tree Species Composition, Richness and Diversity in a Hill Dipterocarp Forest in Malaysia. J. Trop. For. Sci. 2014, 26, 188–202. [Google Scholar]
- Abdullah, S.A.; Hezri, A.A. From Forest Landscape to Agricultural Landscape in the Developing Tropical Country of Malaysia: Pattern, Process, and Their Significance on Policy. Environ. Manage. 2008, 42, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Peh, K.S.-H.; de Jong, J.; Sodhi, N.S.; Lim, S.L.-H.; Yap, C.A.-M. Lowland Rainforest Avifauna and Human Disturbance: Persistence of Primary Forest Birds in Selectively Logged Forests and Mixed-Rural Habitats of Southern Peninsular Malaysia. Biol. Conserv. 2005, 123, 489–505. [Google Scholar] [CrossRef]
- Hamid, W.A.; Rahman, S.B.W.A. Comparison Results of Forest Cover Mapping of Peninsular Malaysia Using Geospatial Technology. IOP Conf. Ser. Earth Environ. Sci. 2016, 37, 012027. [Google Scholar] [CrossRef]
- Şekercioğlu, H.; Primack, R.B.; Wormworth, J. The Effects of Climate Change on Tropical Birds. Biol. Conserv. 2012, 148, 1–18. [Google Scholar] [CrossRef]
- Brodie, J.; Post, E.; Laurance, W.F. Climate Change and Tropical Biodiversity: A New Focus. Trends Ecol. Evol. 2012, 27, 145–150. [Google Scholar] [CrossRef]
- Beier, P.; Brost, B. Use of Land Facets to Plan for Climate Change: Conserving the Arenas, Not the Actors. Conserv. Biol. 2010, 24, 701–710. [Google Scholar] [CrossRef]
- Ghalambor, C.K.; Huey, R.B.; Martin, P.R.; Tewksbury, J.J.; Wang, G. Are Mountain Passes Higher in the Tropics? Janzen’s Hypothesis Revisited. Integr. Comp. Biol. 2006, 46, 5–17. [Google Scholar] [CrossRef]
- Janzen, D.H. Why Mountain Passes Are Higher in the Tropics. Am. Nat. 1967, 101, 233–249. [Google Scholar] [CrossRef]
- Ueda, I.; Shinoda, F.; Kamaya, H. Temperature-Dependent Effects of High Pressure on the Bioluminescence of Firefly Luciferase. Biophys. J. 1994, 66, 2107–2110. [Google Scholar] [CrossRef]
- Dreisig, H. Environmental Control of the Daily Onset of Luminescent Activity in Glowworms and Fireflies (Coleoptera: Lampyridae). Oecologia 1975, 18, 85–99. [Google Scholar] [CrossRef]
- Iguchi, Y. Temperature-Dependent Geographic Variation in the Flashes of the Firefly Luciola Cruciata (Coleoptera: Lampyridae). J. Nat. Hist. 2010, 44, 861–867. [Google Scholar] [CrossRef]
- Iguchi, Y. The Ecological Impact of an Introduced Population on a Native Population in the Firefly Luciola Cruciata (Coleoptera: Lampyridae). Biodivers. Conserv. 2009, 18, 2119–2126. [Google Scholar] [CrossRef]
- Sharma, U.; Goswami, A.; Phukan, M.; Chandra Rajbongshi, S.; Gohain Barua, A. Temperature Dependence of the Flash Duration of the Firefly Luciola Praeusta. Photochem. Photobiol. Sci. 2014, 13, 1788–1792. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Amano, T.; Katoh, K.; Higuchi, H. The Habitat Requirement of the Genji-Firefly Luciola Cruciata (Coleoptera: Lampyridae), a Representative Endemic Species of Japanese Rural Landscapes. Biodivers. Conserv. 2006, 15, 191–203. [Google Scholar] [CrossRef]
- Yuma, M. Effect of Rainfall on the Long-Term Population Dynamics of the Aquatic Firefly Luciola Cruciata. Entomol. Sci. 2007, 10, 237–244. [Google Scholar] [CrossRef]
- Ashton, P.S. Florisitic Zonation of Tree Communities on Wet Tropical Mountains Revisited. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 87–104. [Google Scholar] [CrossRef]
- Ballantyne, L.A.; Lambkin, C.L. A Phylogenetic Reassessment of the Rare S.E. Asian Firefly Genus Pygoluciola Wittmer (Coleoptera: Lampyridae: Luciolinae). Raffles Bull. Zool. 2006, 54, 21–48. [Google Scholar]
- Ballantyne, L.A.; Lambkin, C.L.; Boontop, Y.; Jusoh, W.F.A. Revisional Studies on the Luciolinae Fireflies of Asia (Coleoptera: Lampyridae): 1. The Genus Pyrophanes Olivier with Two New Species. 2. Four New Species of Pteroptyx Olivier and 3. A New Genus Inflata Boontop, with Luciola Indica. Zootaxa 2015, 3959, 1–84. [Google Scholar] [CrossRef]
- Nada, B.; Ballantyne, L.A.; Jusoh, W.F.A. Description of the Larva of a Firefly Species, Pygoluciola Dunguna Nada (Coleoptera: Lampyridae). Zootaxa 2021, 4920, 528–542. [Google Scholar] [CrossRef]
- Blatrix, R.; Lebas, C.; Galkowski, C.; Wegnez, P.; Pimenta, R.; Morichon, D. Vegetation Cover and Elevation Drive Diversity and Composition of Ant Communities (Hymenoptera: Formicidae) in a Mediterranean Ecosystem. Myrmecol. News 2016, 22, 119–127. [Google Scholar]
- Bhardwaj, M.; Uniyal, V.P.; Sanyal, A.K.; Singh, A.P. Butterfly Communities along an Elevational Gradient in the Tons Valley, Western Himalayas: Implications of Rapid Assessment for Insect Conservation. J. Asia. Pac. Entomol. 2012, 15, 207–217. [Google Scholar] [CrossRef]
- Carneiro, E.; Mielke, O.H.H.; Casagrande, M.M.; Fiedler, K. Skipper Richness (Hesperiidae) Along Elevational Gradients in Brazilian Atlantic Forest. Neotrop. Entomol. 2014, 43, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, D.; Tocco, C.; Longoni, A.; Mammola, S.; Palestrini, C.; Rolando, A. Nesting Strategies Affect Altitudinal Distribution and Habitat Use in Alpine Dung Beetle Communities. Ecol. Entomol. 2015, 40, 372–380. [Google Scholar] [CrossRef]
- Nakamura, A.; Kitching, R.L.; Cao, M.; Creedy, T.J.; Fayle, T.M.; Freiberg, M.; Hewitt, C.N.; Itioka, T.; Koh, L.P.; Ma, K. Forests and Their Canopies: Achievements and Horizons in Canopy Science. Trends Ecol. Evol. 2017, 32, 438–451. [Google Scholar] [CrossRef]
- Azevedo-Ramos, C.; De Carvalho, O.; Do Amaral, B.D. Short-Term Effects of Reduced-Impact Logging on Eastern Amazon Fauna. For. Ecol. Manage. 2006, 232, 26–35. [Google Scholar] [CrossRef]
- França, F.M.; Louzada, J.; Barlow, J. Selective Logging Effects on ‘Brown World’ Faecal-Detritus Pathway in Tropical Forests: A Case Study from Amazonia Using Dung Beetles. For. Ecol. Manage. 2018, 410, 136–143. [Google Scholar] [CrossRef]
- Hosaka, T.; Niino, M.; Kon, M.; Ochi, T.; Yamada, T.; Fletcher, C.; Okuda, T. Impacts of Small-Scale Clearings Due to Selective Logging on Dung Beetle Communities. Biotropica 2014, 46, 720–731. [Google Scholar] [CrossRef]
- Kaufmann, T. Ecological and Biological Studies on the West African Firefly Luciola discicollis (Coleoptera: Lampyridae). Ann. Entomol. Soc. Am. 1965, 58, 414–426. [Google Scholar] [CrossRef]


| Term | Res.Df | Df.diff | Dev | Pr (>Dev) |
|---|---|---|---|---|
| (Intercept) | 15 | |||
| Mountain | 11 | 1 | 4179.9 | 0.108 |
| Forest type | 7 | 1 | 4159.3 | 0.001 |
| Term | DF | SS | R2 | F | Pr (>Dev) |
|---|---|---|---|---|---|
| Mountain | 4 | 1.7638 | 0.33130 | 2.4132 | 0.02 |
| Forest type | 4 | 2.2809 | 0.42844 | 3.1208 | 0.005 |
| Residual | 7 | 1.2790 | 0.24025 | ||
| Total | 15 | 5.3237 | 1.00000 |
| Term | Partial η2 | Sum Sq | Df | F Value | Pr (>F) |
|---|---|---|---|---|---|
| Forest type | 0.3253 | 2.3011 | 4 | 0.8438 | 0.5395 |
| Canopy closure | 0.1519 | 0.8546 | 1 | 1.253 | 0.2998 |
| Leaf litter depth | 0.0561 | 0.2834 | 1 | 0.4157 | 0.5396 |
| Understorey height | 0.0154 | 0.0746 | 1 | 0.1095 | 0.7504 |
| Number of large trees | 0.0018 | 0.0086 | 1 | 0.0126 | 0.9137 |
| Residuals | NA | 4.7725 | 7 | NA | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nada, B.; Beckerman, A.P.; Evans, K.L.; Ballantyne, L.A. The Effects of Tropical Elevations and Associated Habitat Changes on Firefly (Coleoptera: Lampyridae) Diversity in Malaysia. Diversity 2023, 15, 79. https://doi.org/10.3390/d15010079
Nada B, Beckerman AP, Evans KL, Ballantyne LA. The Effects of Tropical Elevations and Associated Habitat Changes on Firefly (Coleoptera: Lampyridae) Diversity in Malaysia. Diversity. 2023; 15(1):79. https://doi.org/10.3390/d15010079
Chicago/Turabian StyleNada, B., Andrew P. Beckerman, Karl L. Evans, and Lesley A. Ballantyne. 2023. "The Effects of Tropical Elevations and Associated Habitat Changes on Firefly (Coleoptera: Lampyridae) Diversity in Malaysia" Diversity 15, no. 1: 79. https://doi.org/10.3390/d15010079
APA StyleNada, B., Beckerman, A. P., Evans, K. L., & Ballantyne, L. A. (2023). The Effects of Tropical Elevations and Associated Habitat Changes on Firefly (Coleoptera: Lampyridae) Diversity in Malaysia. Diversity, 15(1), 79. https://doi.org/10.3390/d15010079