Abstract
A new, very small-sized species of Arthroleptis is described from western Rwanda. The new species occurs locally endemic in Nyungwe Forest and Cyamudongo Forest, where it inhabits the leaf litter of montane forests between 1800 and 2200 m a.s.l. It differs from all other members of the genus by a unique combination of morphological characters, including size (SVL of adult males 16.0–16.5 mm), skin texture, length of hindlimbs, and ventral colour pattern; by characteristics of its advertisement call that consists of a single note lasting 17.4 ± 6.4 [11–32] ms and has a dominant frequency of 5861 ± 188 [5531–6029] Hz; and also in the sequence of the 16S rRNA gene that differs from available homologous sequences of other species of the genus by an uncorrected p-distance of at least 4.6%. Details of the natural history as well as two additional call types are described.
1. Introduction
The genus Arthroleptis Smith, 1849 contains about 48 species that are distributed in almost all parts of sub-Saharan Africa [1,2]. There are several taxonomic problems and disagreement with respect to the validity of particular species and probably a number of cryptic species are currently unrecognized [3,4,5,6,7]. Historically, several small-sized members of the genus had been placed in a separate genus, Schoutedenella de Witte, 1921 [3,4,8,9], but the validity of the genus had not been accepted by most authors (e.g., [5,10,11]). Phylogenetic analyses have repeatedly demonstrated that small-sized Arthroleptis species considered to belong to Schoutedenella do not form a distinct clade separated from larger species and Schoutedenella is now widely recognized as a junior synonym of Arthroleptis [6,12]. A number of small Arthroleptis species have been described from the mountains and the graben valley of the Albertine Rift [3,8,13,14,15,16], and three species of Arthroleptis have been recorded from Rwanda [17]. Arthroleptis adolfifriederici Nieden, 1911 was described from the “Bugoiewald” and the “Rugegewald” (nowadays Gishwati and Nyungwe Forest, respectively) in Rwanda and is easily identified by its large size, smooth dorsum and long and slender limbs [13,17]. Arthroleptis schubotzi Nieden, 1911 has its type locality at “Usumbura” (=Bujumbura, Burundi) and was later also recorded from southwestern Tanzania, eastern Democratic Republic of the Congo, and western Rwanda [8,13,15,17,18,19]. It is a small-sized species (SVL of male and female < 24 mm) with a granular dorsal skin, a grey venter with blackish network and reddish thighs [17]. I repeatedly collected another very small species of Arthroleptis in Nyungwe and Cyamudongo Forest (Arthroleptis sp. [17]) with shagreened dorsal skin and a yellowish brown ventral colouration with yellow longitudinal stripes. Herein, I present the results of morphological, bioacoustics, and genetic comparisons of this species with its congeners and address its taxonomic status.
2. Materials and Methods
Collection of specimens and comparative data: Specimens of Arthroleptis examined for this study (Table A1) were collected in Rwanda on several trips between September 2010 and December 2018. Specimens were collected by hand, photographed in life, and euthanized using a chlorobutanol solution. Tissue samples for molecular analyses were taken from liver or thigh muscle and stored in 98% ethanol. Specimens were fixed and stored in 70% ethanol and are deposited in the herpetological collection of the Zoologisches Forschungsmuseum Alexander Koenig, Bonn, Germany (ZFMK), and Staatliches Museum für Naturkunde, Stuttgart, Germany (SMNS), and the working collection of the author (JMD). Comparative material (Table A1), including the types of most of the relevant species, was examined in the Royal Museum for Central Africa, Tervuren, Belgium (RMCA), and the Museum für Naturkunde, Leibniz-Institut für Evolutions- und Biodiversitätsforschung an der Humboldt-Universität zu Berlin, Germany (ZMB).
Morphological examination: I took the following measurements from specimens with a digital calliper (to the nearest 0.1 mm): snout-vent length (SVL, from tip of snout to vent); head length (HL, distance from posterior end of jaw to tip of snout); head width (HW, measured at widest point); horizontal eye diameter (ED); horizontal tympanum diameter (TD); snout length (SL, distance between anterior margin of eye and tip of snout); eye-to-nostril distance (EN, distance between anterior margin of eye and centre of nostril); nostril-to-snout distance (NS, distance between centre of nostril and tip of snout); internarial distance (NN, distance between centres of nostrils); intereye distance (EE, distance between anterior margins of eyes); upper eyelid width (EW, measured at widest point); interorbital distance (IO, shortest distance between upper eyelids); tibiofibula length (TFL, measured with both knee and tibio-tarsal articulation flexed); thigh length (THL, from knee to vent with thigh held perpendicularly to median body plane); foot length (FOL, distance from proximal end of inner metatarsal tubercle to tip of fourth toe); hand length (HND, measured from proximal end of inner metacarpal tubercle to tip of third finger); length of third finger (3FL, from metacarpus to tip of third finger). Additionally, I recorded the following qualitative characters: size and shape of thenar and palmar tubercles; presence of maxillary, premaxillary and mandibular teeth; presence of vomer ridges; shape of supratympanic fold; presence, extent and shape of dermal ridges of dorsum. Sex of males was determined by presence of transparent skin in the gular region and elongate third fingers, and examination of the gonads. Sex of females was determined by presence of ova. Data on colour pattern were obtained from specimens photographed in life. Additional data on morphological characters used for comparison were taken from [1,3,5,7,8,13,14,15,16,20,21,22].
Phylogenetic analyses: DNA was extracted from the tissue samples using Qiagen DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. Polymerase Chain Reaction (PCR) was used to amplify a fragment of approximately 550 base pairs of the 16S mitochondrial rRNA using standard primers: 16SAL (50-CGCCTGTTTACTAAAAACAT-30), and 16SBH (50-CCGGTCTGAACTCAGATCACGT-30). Amplification followed the standard PCR conditions [23] with the following thermal cycle profile: 120 s at 94 °C, followed by 33 cycles of 94 °C for 30 s, 49 °C (12S)/53 C (16S) for 30 s, and extension at 65 °C for 60 s. All amplified PCR products were verified using electrophoresis on a 1.4% agarose gel stained with ethidium bromide. PCR products were purified using High pure PCR Product Purification Kit (Roche Diagnostics, Rotkreuz, Switzerland). Sequencing of both strands was performed with the DYEnamic ET Terminator Cycle Sequencing Premixkit (GE Healthcare, Munich, Germany) for sequencing reactions run on a Mega-BACE 1000 automated sequencer (GE Healthcare). Chromas lite 2.1.1 software (Technelysium Pty Ltd., Brisbane, Australia) and read the chromatograms of the sequences. The obtained sequences were compared with those in GenBank using a standard nucleotide-nucleotide BLAST search. The homologous sequences most similar to the sequences of the Rwandan species of Arthroleptis were downloaded and incorporated in an alignment. A sequence of Leptopelis karissimbensis from Rwanda was used as an outgroup. Sequences used in the study are listed in Table A2. The sequences were aligned using the MUSCLE algorithm [24] implemented in MEGA version X [25]. The final alignment contained 24 sequences and was 563 bp in length. Pairwise distances were calculated in MEGA version X. The general time-reversible model with proportion of invariable sites and gamma-distributed rate variation among sites (GTR + I + G) was chosen as the best-fitting model of sequence evolution on the basis of the Akaike information criterion as implemented in jModelTest 2 [26] and was applied in a maximum likelihood (ML) analysis, performed in MEGA version X with heuristic searches with stepwise addition and TBR branch-swapping algorithm, generating 1000 bootstrap replicates.
Bioacoustics: Advertisement calls were recorded as uncompressed files in WAVE format with a Sony PCM–D50 Linear PCM Recorder with built-in stereo microphones (Sony Deutschland GmbH, Cologne, Germany). For the characterisation of the advertisement call, I analysed a total of 37 calls from six different males. Stereo recordings were converted to mono at a sampling rate of 44.1 kHz and 16 bits resolution using Adobe Audition 1.5. Spectrograms and waveforms were obtained applying Blackman–Harris Fast Fourier transformation with a FFT window width of 1024 points. Temporal data were obtained from the waveforms and frequency information was obtained from the spectrograms and power spectra. Values are given as mean ± standard deviation with range in parentheses. Definitions of acoustic parameters follow [27].
3. Results
Morphological, bioacoustic, and phylogenetic comparisons revealed that the small Arthroleptis species from Nyungwe cannot be assigned to any of the described species, nor any of the taxa considered synonyms. Therefore, it is herein described as a new species.
- Arthroleptis nyungwensis sp. nov.
- Nyungwe Squeaker
- urn:lsid:zoobank.org:act:E8F25561-AD7C-46D8-9107-73F6ED10799C
- Arthroleptis sp.—Dehling & Sinsch 2023: 13 [17].
Holotype: ZFMK 104075, adult male, from a small stream near Rukuzi (2.4635 S, 29.2293 E; 2200 m), Nyungwe National Park, Western Province, Rwanda, collected on 17 September 2014 by J. M. Dehling.
Paratypes: ZFMK 104076, adult male, from the type locality, collected in March 2013 by J.M. Dehling; ZFMK 104077, adult male, from Cyamudongo Forest (2.546 S, 28.989 E, about 2000 m), Western Province, Rwanda, collected 21 October 2018 by J.M. Dehling; SMNS 15743 and 15744, two subadult males, from the edge of Cyamudongo Forest (2.5378 S, 29.9942 E; 1830 m), Western Province, Rwanda, collected 14 June 2014 by J.M. Dehling.
Referred material: JMD 599, adult male, from the Kamiranzovu Waterfall Trail, appr. 1800 m a.s.l., Nyungwe National Park, Western Province, Rwanda, collected 25 September 2010 by J. M. Dehling.
Diagnosis: The species is assigned to the genus Arthroleptis for exhibiting the following morphological characteristics [28]: pedal webbing absent, median dorsal skin raphe present, third finger elongate in adult males, dermal spines present on third finger. The new species differs from all other members of the genus by the combination of the following characteristics: small size (SVL of adult males 16.0–16.5 mm); dorsal surfaces of head, trunk, and limbs and lateral surfaces of trunk finely shagreened; tympanum well visible externally; median lingual process absent; legs long, tibiotarsal joint reaching to eye; tibiofibular length equal to foot length; tip of third finger and tips of toes slightly enlarged; inner metatarsal tubercle very small and rounded, outer metatarsal tubercle absent; ventral colour reddish to orange with two slightly concave longitudinal yellow stripes and white dots; advertisement call consisting of single note, lasting 17.4 ± 6.4 [11–32] ms and dominant frequency at 5861 ± 188 [5531–6029] Hz; sequence of the 16S rRNA gene differing from available homologous sequences of other species by uncorrected p-distance of at least 4.6%.
Description of the holotype: Measurements of the holotype are provided in Table 1. Very small (SVL 16.5 mm) adult male; body slender, widest at temporal region, slightly tapering to groin (Figure 1A,B); limbs slender; head large (HL/SVL 0.37, HW/SVL 0.33), longer than wide (HW/HL 0.89); snout moderately long (SL/HL 0.41), wider than long (SL/EE 0.79); rostral tip rounded in both dorsal view and lateral view; canthus rostralis moderately distinct, straight between eye and nostril; loreal region oblique; nostrils rounded, directed laterally, situated about halfway between eye and tip of snout (EN/NS 0.97), separated from each other by distance much larger than distance between eye and nostril (NN/EN 1.68); eye directed anterolaterally, hardly protruding, projecting just beyond margins of head in dorsal view, relatively large (ED/HL 0.35), its diameter shorter than snout (ED/SL 0.84); dorsal surface of eye lids slightly above dorsal surface of head in lateral view; interorbital distance much wider than upper eyelid (IO/EW 1.61) and smaller than internarial distance (IO/NN 0.89); tympanum well visible externally, its annulus well defined; upper jaw with dentition; teeth on premaxilla larger than teeth on maxilla; choanae small, rounded, located far anterolaterally at anterolateral margin of roof of mouth, completely covered by maxillary bone, not visible in ventral view; vomer ridges and teeth absent; tongue long and moderately broad, posteriorly expanded, hardly bilobed, free distally for about two-thirds its length; median lingual process absent.

Table 1.
Morphometrics of the adult male type specimens of Arthroleptis nyungwensis sp. nov. (in mm). For abbreviations, see Materials and Methods.

Figure 1.
(A) Dorsolateral view and (B) ventral view of the holotype (ZFMK 104075) of Arthroleptis nyungwensis sp. nov. in life. (C) Dorsolateral view and (D) ventral view of an adult male (JMD 599) of Arthroleptis nyungwensis sp. nov. in life. (E) Dorsolateral view and (F) ventral view of one of the subadult paratypes (SMNS 15743) of Arthroleptis nyungwensis sp. nov. in life.
Dorsal surfaces of head, trunk, and limbs and lateral surfaces of trunk finely shagreened (Figure 1A); median skin raphe barely visible on dorsal side in preservative, only discernible on head surface; supratympanic region smooth with no dermal fold; ventral side of head smooth; vocal sac present; chest and anterior part of abdomen smooth, posterior part of abdomen weakly areolate; ventral side of limbs smooth (Figure 1B); short transverse fold above vent.
Forelimbs slender; hand small, but with very long third finger (HND/SVL 0.42); few keratinous spines along lateral sides of third finger; tips of fingers not enlarged into small disc, except slightly in Finger III; relative length of fingers: I < IV < II < III; subarticular tubercles well developed, numbering one on Fingers I and II, three on Finger III and two on Finger IV, proximal tubercles on Fingers III and IV and tubercles on Fingers I and II singular, rounded; distal tubercles on Fingers III and IV indistinctly bipartite; fingers without webbing; thenar tubercle small, low and oval; inner palmar tubercle large, low, and rounded, outer palmar tubercle absent.
Hind limbs slender, moderately long; heel reaching to level of eye when legs adpressed forwardly to body; crus moderately long (TFL/SVL 0.42), slightly shorter than thigh (TFL/THL 0.94); foot length about equal to crus length (FOL/TFL 1.03); relative length of toes: I < II < VI < III < IV; toe tips slightly enlarged into small disks; subarticular tubercles singular, numbering one on Toes I and II, two on Toes III and V, and three on Toe IV; toes without webbing; inner metatarsal tubercle small, rounded and moderately prominent, about one-third length of metatarsus of Toe I; outer metatarsal tubercle indiscernible.
Colouration in life. Dorsal basic colouration light brown. Slightly darker brown pattern on head and trunk hardly discernible, consisting of dark transversal interorbital bar and faint broad longitudinal band along spine (Figure 1A). Top of head anterior to interorbital bar lighter brown. Dark brown stripe in straight line along ventral part of canthus rostralis from tip of snout to anterior edge of eye, continued downwardly curved from posterior edge of eye almost to arm insertion, just touching the posterodorsal edge of tympanum (Figure 1A). Lateral sides of head and trunk lighter, greyish to yellowish brown; dorsal side of upper arms and hands yellowish brown. Sides of head and forearm loosely, flanks more or less regularly and densely scattered with small white dots (Figure 1A). Base colour of ventral side yellowish brown with tiny dark brown speckles, most densely on ventral side of head; ventral side of head along the jaw and ventral parts of forelimbs darker reddish brown; ventral parts of leg and lateral sides of thigh dark red (Figure 1B); abdomen yellowish brown with two bright yellow longitudinal, slightly concave stripes, faintly merging with each other at posterior end (Figure 1B); small white dots on ventral side, few on forearm and thigh, densely on lateral sides of abdomen, around cloaca and on tarsus; irregularly shaped yellow flecks on abdomen (Figure 1B).
Colouration in preservative. Dorsal basic colouration faded to greyish brown. Darker greyish brown dorsal pattern more conspicuous than in life. Ventral colour faded to uniformly yellowish creme-coloured. Dark brown speckling more conspicuous than in life. Yellow colour pattern indiscernible.
Variation. The adult male paratypes match the male holotype in general appearance, proportions, colouration, and colour pattern. Mensural variation within the species is shown in Table 1. The dorsal colour pattern was more conspicuous in life in one of the adult males (JMD 599) and in the subadults (Figure 1C,E). In the subadult specimens, the sides of the head and the flanks were dark grey in life and there was a black stripe running in a straight line from the nostril to the anterior edge of the eye and slightly ventrally curved from the posterior edge of the eye above the tympanum to the level of the arm insertion; the venter was light yellow in the centre, darkening to dark yellow then orange on the edges, without the conspicuous yellow stripes (Figure 1E,F).
Bioacoustics. Series of advertisement calls of the holotype, two paratypes, another collected and two uncollected males were recorded at different ambient temperatures between 12 °C and 20 °C. Males called from the leaf litter on the forest floor during both night and day. The most-often-emitted vocalisation was a single note (Figure 2A), and I regard this as the advertisement call. The call was repeated in series at irregular intervals of 2.5–8.8 s. In most recordings, the note appeared unpulsed or diffusely pulsatile, but in a few instances, two separate pulses were visible (Figure 2A). The note had a total duration of 17.4 ± 6.4 [11–32] ms (N = 5 males). Individual pulses, when traceable, were separated by an interval of 1 ms duration and had an individual duration of 7 ms (first pulse) and 5 ms (second pulse (Figure 2A). There was no measurable frequency modulation in shorter notes of less than 20 ms and dominant frequency was at 5861 ± 188 [5531–6029] Hz. A fundamental was present at about 2950 Hz, and harmonics were present at 9000, 12,000, 15,000, and 18,000 Hz (Figure 2A). Prevalent bandwidth was 5400–6300 Hz. In a few longer notes of more than 30 ms, the dominant frequency increased from 5404 ± 30 [5383–5426] Hz in the first third of the note to 5598 Hz in the last third of the note.
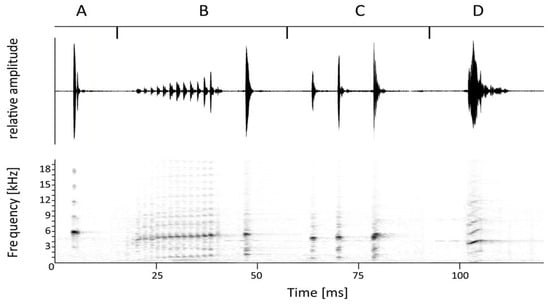
Figure 2.
Waveform (top) and corresponding sound spectrogram of (A) an advertisement call of Arthroleptis nyungwensis sp. nov., (B) an aggressive call, consisting of a thirteen-notes group followed by the main note, (C) a variant of the aggressive call, and (D) a possible female courtship call. See text for details.
One of the paratypes (ZFMK 104077) was collected while duetting with another uncollected male. The two males emitted a second call type (Figure 2B,C). It was impossible to differentiate between the two males in the recording, and therefore the following description is based on all recorded calls. The second call type likewise contained a prominent last note lasting 21.4 ± 10.1 [11–39] ms with the dominant frequency increasing from 5175 ± 197 [4866–5469] Hz in the first third of the note to 5480 ± 130 [5167–5727] Hz in the last third of the note (Figure 2B,C). However, the main note was always initiated by a note group that had a much lower amplitude and was separated from the main note by an interval of 90.1 ± 13.6 [71–113] ms. The composition of the note group varied considerably. In about half of the recorded calls, it consisted of an uninterrupted series of 1–13 notes (Figure 2B), in the other half, two or three subgroups, each consisting of 1–3 notes, were separated from each other by intervals of 37.3 ± 22.1 [19–86] ms. As calls with both interrupted and uninterrupted pulse groups were recorded in quick succession, these differences were not attributable to the two different calling individuals. Individual notes within the (sub)groups were repeated at a rate of 58.6 ± 4.9 [47.6–62.5] per second, were weakly pulsatile, and had a duration of 8.8 ± 3.3 [4–14] ms. Amplitude increased from the first to the last note of the group (Figure 2B,C), going along with an increase in dominant frequency from 4658 ± 323 [4177–5124] Hz in the first note to 4944 ± 210 [4608–5254] Hz in the last note (Figure 2B,C). Whereas the interrupted and uninterrupted note groups sounded like a trill, in one case, two widely separated initial notes had a relatively high amplitude, giving the impression that the main note was rapidly repeated (Figure 2C). Total duration of the second call type was 209.1 ± 40.2 [139–285] ms.
On the same recording, a third call type was identified. It had been recorded a total of three times and was a brief whistle that consisted of a single pulsatile note, lasting 40, 44 and 74 ms. The dominant frequency was markedly modulated, increasing from 3760 ± 108 [3660–3875] Hz in the first third to 4205 ± 25 [4177–4220] Hz in the last third (Figure 2D).
Habitat and natural history: Calling males were observed in moist leaf litter of closed-canopy montane forest between 1800 and 2200 m a.s.l. Subadults were found at the forest edge in Cyamudongo Forest. The species is active at both night and day. The reproduction is unknown, but the species is suspected to deposit eggs in moist leaf litter and to undergo direct development without a tadpole stage.
Etymology: The species is named after Nyungwe Forest in southern Rwanda.
Distribution and Conservation: Despite intensive surveys in Nyungwe Forest and elsewhere in Rwanda [17,29], the species is currently known from only a few localities in the western part of Nyungwe Forest and in Cyamudongo Forest (Figure 3). I propose the species to be classified as “Near Threatened” according to the criteria established by the International Union for Conservation of Nature (IUCN) [7] in both the global Red List and the Rwandan national Red List [17].
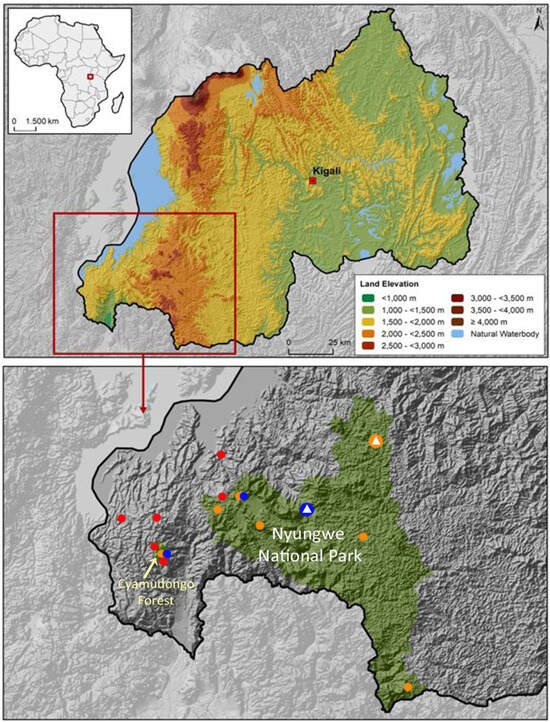
Figure 3.
Map of Rwanda with major elevation zones and natural water bodies (top) and detail of map showing Nyungwe National Park including Cyamudongo Forest (bottom) in southwestern Rwanda; forested areas are displayed in green. The distribution of the three species of Arthroleptis in Nyungwe National Park and surroundings is indicated by coloured dots at localities where specimens were collected or observed: Arthroleptis adolfifriederici (orange), A. nyungwensis sp. nov. (blue), A. schubotzi (red). Type localities are marked with a white triangle in the centre of the coloured dot. Map courtesy of S. Seidel.
Phylogenetic relationships: The phylogenetic tree resulting from the ML analysis was well supported by nonparametric bootstrap values (Figure 4). All samples of Arthroleptis nyungwensis formed a clade that was weakly supported as being sister to a clade containing two deeply divergent clades assigned to A. schubotzi, one from Rwanda (A. schubotzi A), the other from Bwindi, Uganda (A. schubotzi B). The three clades are all from montane forests in the central and northern Albertine Rift. They were sister to a clade containing A. xenodactyloides and A. xenochirus (Figure 4), two species that are distributed from Angola to southern DRC, southern Tanzania and northwards to Kenya. Another sequence assigned to Arthroleptis xenodactyloides was resolved as sister to all aforementioned clades, suggesting that the identification of the source specimens of these three latter sequences need to be revised. The third Rwandan species, A. adolfifriederici, is only distantly related to A. nyungwensis and A. schubotzi and appears to be most closely related to A. tanneri from the Usambara Mountains in Tanzania (Figure 4). The samples of A. nyungwensis differed from each other by 0.00–0.54% and from samples of all other species by at least 4.6% in uncorrected p-distance. Samples of A. schubotzi A from Rwanda differed from the sample of A. schubotzi B from Uganda by 5.6%.
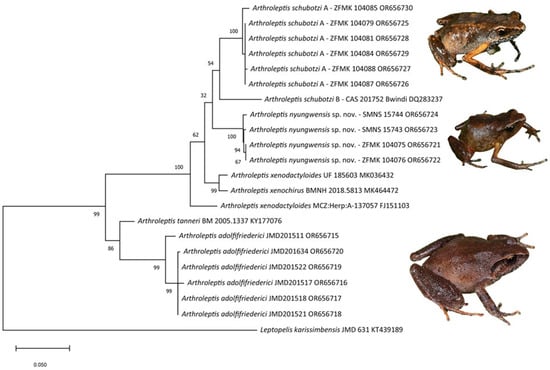
Figure 4.
Phylogram inferred from mitochondrial nucleotide sequence data of 16S rRNA (563 BP). Numbers below branches are non-parametric bootstrap support values from ML (1000 replicates). Sequences are listed with species name, voucher number, and GenBank accession number, respectively. Specimens of the three species occurring in Rwanda are depicted on the right at the level of their corresponding samples.
Morphological comparison: The very small size (SVL of adult males 16.0–16.5 mm) distinguishes A. nyungwensis sp. nov. from the other species in the wider region that are larger, i.e., A. adolfifriederici: males 27.6–32.0 mm; A. francei Loveridge, 1953: male 32 mm; A. phrynoides (Laurent, 1976): male 20.4 mm; A. reichei Nieden, 1911: males to 30 mm; A. schubotzi: males 19.9–21.2 mm (including Schoutedenella discodactyla Laurent, 1954 with males 17.5–19 mm); A. spinalis Boulenger, 1919: male 21.4 mm; A. stenodactylus Pfeffer, 1893: males to 33 mm; A. xenochirus Boulenger, 1905 (including Schoutedenella globosa de Witte, 1921 as well as A. lameeri de Witte, 1921 and Schoutedenella muta de Witte, 1933; see also [5]): males with enlarged third finger 16.5–24 mm; A. xenodactylus Boulenger, 1909: males to 26 mm; A. xenodactyloides Hewitt, 1933: males to 25 mm. The very small inner metatarsal tubercle distinguishes the new species from all remaining species with large, elongate, prominent inner metatarsal tubercle that is about two-thirds the length of the first metatarsus, i.e., A. loveridgei de Witte, 1933 (also outer palmar tubercle present, ventral side of thigh coarsely granular, discs of toes strongly enlarged vs. outer palmar tubercle absent, thigh smooth, discs of toes slightly enlarged); A. sylvaticus (Laurent, 1954) (also discs of toes strongly enlarged vs. slightly enlarged); and A. xenochirus Boulenger, 1905 (also venter uniformly off-white, throat grey in males vs. venter reddish to orange with longitudinal yellow stripes and white dots, throat yellowish brown with tiny dark brown speckles). Arthroleptis fichika Blackburn, 2009 and A. kidogo Blackburn, 2009 are known only from females that are much smaller (SVL 13.5–14.2 mm and 13.9–14.1 mm, respectively) than the new species, and differ in having a dappled ventral colour pattern [7].
Morphological differentiation between the similarly sized species of Arthroleptis from the Albertine Rift is hampered by the shortage of external diagnostic characters, especially in preserved specimens, in combination with the briefness of the original descriptions that specify only a handful of characteristics, some of which are not diagnostic. In addition, the whereabouts and the identity of the types of several species described by Laurent [3,16] are currently vague. The life colouration can be a useful character to easily distinguish between species if it is thoroughly documented. The distributional range of Arthroleptis nyungwensis sp. nov. overlaps with that of A. schubotzi in western Rwanda, although the two species are usually found in different habitats. The Rwandan specimens match the types of A. schubotzi and the types of its junior synonym Schoutedenella kivuensis de Witte, 1941, very well. Arthroleptis schubotzi (males 19.9–21.2 mm, male syntype of S. kivuensis 19.8 mm; females 21.1–21.6 mm; female holotype currently 19.2 mm, previously measured 21 mm [13] and 20 mm [20], female syntype of S. kivuensis 20.6 mm) is significantly larger than A. nyungwensis sp. nov. (males 16.0–16.5 mm), has relatively shorter legs (TFL/SVL 0.36–0.41 vs. 0.42–0.46 in A. nyungwensis), a relatively larger tympanum (TD/EYE in adult males >0.60 vs. <0.48), a relatively shorter head (HL/SVL 0.31–0.33 vs. 0.35–0.37), and its dorsum is granular, covered with small, low tubercles, especially in males (Figure 5; vs. dorsum finely shagreened). It is most easily distinguished from A. nyungwensis sp. nov. (venter reddish to orange with longitudinal yellow stripes and white dots, Figure 1) by its conspicuous ventral pattern consisting of an off-white background with dark reticulation, with the ventral side of the head having a higher proportion of dark pattern than the breast and abdomen in females and the gular region of males being black with few whitish flecks, and bright red thighs (Figure 5). This colour pattern was also described in the female holotype of A. schubotzi [20] and described and depicted for the types of Schoutedenella kivuensis [15]. There are a number of similarly sized, but poorly defined taxa from eastern DRC that are only known from the type series [3,16] and whose taxonomic status is doubtful. A very similar ventral colour pattern to A. schubotzi is present in the types of Arthroleptis mossoensis (Laurent, 1954), Schoutedenella discodactyla (currently considered a junior synonym of A. schubotzi) and A. pyrrhoscelis Laurent, 1952, and these taxa are readily distinguished from A. nyungwensis sp. nov.: Arthroleptis mossoensis was described from Murugaragara, Mosso, Territoire de Rutana, Burundi, at an altitude of 1200 m, based on a single female (SVL 22.4 mm) that remains the only specimen of the species ever collected. It further differs from A. nyungwensis sp. nov. in its short hindlimbs (TFL/SVL 0.36 vs. 0.42–0.46), the tibio-tarsal articulation only reaching the shoulder (vs. reaching the eye). Arthroleptis pyrrhoscelis from the Kabobo Plateau in eastern DRC is similar in size (male types 15.1–16.2 mm) to A. nyungwensis sp. nov., but has shorter hindlimbs, the tibiotarsal joint reaching only to tympanum (vs. to eye). The posterior part of the abdomen and the ventral side of the thighs are coarsely granular (vs. weakly areolate), an outer palmar tubercle and several prominent metacarpal tubercles are present (vs. absent), and the inner metatarsal tubercle is large, elongate and prominent (vs. small and rounded). Schoutedenella discodactyla from Lutunguru in the North Kivu Province of DRC (currently referred to A. schubotzi) differs by its markedly dilated discs of fingers and toes (vs. tip of third finger only and tips of toes slightly enlarged). Arthroleptis vercammeni (Laurent, 1954) from Mwana in eastern DRC is a very small species (SVL of males 13–15 mm, of females 15–17 mm [3]) and thus even smaller than A. nyungwensis sp. nov. (males 16.0–16.5 mm), the throat is blackish in males (vs. yellowish brown with tiny dark brown speckles), the tibia is noticeably longer than the foot (TFL/FOT 1.10–1.25 vs. 0.97–1.03 in A. nyungwensis), and the tips of fingers and toes are clearly dilated [3] (vs. tip of third finger only and tips of toes slightly enlarged). Arthroleptis hematogaster (Laurent, 1954) is a poorly known species from the South Kivu Province in eastern DRC. The species is similar in size (only known male 16.5 mm), but is reported to have a uniform ventral colouration varying from orange-red to blood-red [3] (vs. yellowish brown) and lacks the distinct two longitudinal yellow ventral stripes and the white dots on venter, arms and thighs that are present in A. nyungwensis sp. nov., the foot is relatively shorter with FOL/SVL 0.38–0.41 (vs. 0.43–0.45), and finger tips and toe tips are strongly enlarged (vs. slightly enlarged).

Figure 5.
Arthroleptis schubotzi from Cyamudongo Forest, Rwanda, in life. (A) Dorsal view of adult female (ZFMK 104089) and (B) adult male (ZFMK 104090). (C) Corresponding ventral view of adult female and (D) adult male, showing the distinct ventral colour pattern.
Bioacoustic comparison: The advertisement calls of only about half the species of Arthroleptis are known [1]. The advertisement call of Arthroleptis nyungwensis sp. nov. (single note, 17.4 ± 6.4 [11–32] ms duration, dominant frequency at 5861 ± 188 [5531–6029] Hz; Figure 2) differs from all described calls of its congeners. The call of one of the two other species of Arthroleptis recorded from Rwanda, A. adolfifriederici, is similar in consisting of a single, brief (52–65 ms) note, but has a much lower dominant frequency of about 2900 Hz [17]. The known advertisement calls of the phylogenetically closely related species differ markedly from the call of A. nyungwensis sp. nov., i.e., A. xenochirus: high-pitched trill, seven notes, repeated at a rate of 15/s, dominant frequency at 4600 Hz [22]; A. xenodactylus: brief whistle, duration 50 ms, repeated two or three times per second, dominant frequency at 6400–7000 Hz [22]; A. xenodactyloides: single note, 88–95 ms, repeated at an interval of 340–390 ms, composed of three pulses, each lasting 16–24 ms, dominant frequency at 6300–6500 ms [30]; brief, cricket-like chirp, consisting of three brief clicks, dominant frequency at 5500 Hz [22]; call duration 0.05–0.1 s, repeated every 0.3–0.7 s, dominant frequency 5.2–6.4 kHz, 2–4 pulses [31].
Based on observations of males of its junior synonym Schoutedenella kivuensis, the call of A. schubotzi has been described onomatopoeically as “cri-cri, cri-cri” and characterized as “extremely piercing and very characteristic cry, absolutely similar to that of S. globosa” [=A. xenochirus], “quite comparable to the ‘song’ of our cricket, but much louder” [15]. This was subsequently cited as a “harsh series of double chirps” [1,22]. The call description agrees with the results of the analysis of call recordings of A. schubotzi from Rwanda [17]: call duration 246–429 ms, 5–6 notes, each lasting 23–29 ms and composed of three pulses, internote interval 34–40 ms, dominant frequency 4331–4450 Hz. The advertisement call of males assigned to A. schubotzi from Kibale National Park, Uganda, differs from the advertisement call of Rwandan specimens in consisting of only either two or three notes, the first two being separated from each other by an interval of 80 [59–98] ms, the second and third by an interval of 94 [84–113] ms; in having a slightly higher dominant frequency of 4661 [4462–4892] Hz; and in a briefer call duration of 158 [92–273] ms [32]. The advertisement calls of both populations assigned to A. schubotzi differ from the advertisement call of A. nyungwensis.
4. Discussion
The mountains of the Alberine Rift harbour the type localities for many small Arthroleptis species [3,8,13,14,15,16]. Several of these species are known only from their types, and in some cases their taxonomic status is unclear. Examination of the available type material and a survey of the available literature suggest that the group is in urgent need of a thorough revision. For example, Arthroleptis mossoensis and A. pyrrhoscelis match A. schubotzi in many characteristics such as size, relative leg length, rough dorsal skin, and the conspicuous ventral colour pattern, and were described from roughly the same geographic region. The status of these taxa currently recognized as distinct species need to be reassessed. On the other hand, the ML analysis of phylogenetic relationships revealed two deeply divergent clades between populations currently assigned to A. schubotzi, one from Rwanda (“A. schubotzi A”), the other from Bwindi, Uganda (“A. schubotzi B”). There are also marked advertisement-call differences between populations of A. schubotzi from Rwanda [17] and from Kibale, Uganda [32]. These differences indicate that two or more unrecognized species could be admixed under the nominal taxon A. schubotzi. These taxonomic issues will be addressed in an upcoming study.
Arthroleptis nyungwensis is usually found in closed-canopy montane forest and rarely at the forest edge, whereas A. schubotzi is abundant at forest edges and in anthropogenically disturbed habitats. Therefore, both species usually occur in parapatry to each other in western Rwanda. Calling males of both species were, however, found syntopically at one locality in Cyamudongo Forest, in a transition between secondary forest and a buffer zone. The advertisement call of A. nyungwensis differs more markedly from the advertisement call of A. schubotzi from Rwanda than from the allopatric population in Kibale, Uganda, assigned to A. schubotzi [32]. The second call type recorded from one of the paratypes of A. nyungwensis is very similar to the vocalisation of A. schubotzi from Kibale National Park, Uganda, which was identified as an aggressive call (sensu [33]) and consisted of either one or two elongated pulse trains, followed by a single high-pitched metallic note similar to those of the advertisement call [32]. Given the circumstances at the time of the recording and its similarity to the aggressive call of “A. schubotzi”, the second call type of A. nyungwensis appears to be an aggressive call directed towards competing males during male-male interactions. The third call type of A. nyungwensis differs markedly from the advertisement and the aggressive call of males. Considering its characteristics and the fact that the dominant frequency was much lower than in calls emitted by males, the most plausible explanation is that it was the courtship call of a female [33] that had also been present at the scene but had been overlooked. This, however, needs to be confirmed by a direct observation of a calling female.
Several new species of amphibians and reptiles have recently been discovered or recorded for the first time from Nyungwe Forest and its surroundings [17,34,35,36,37,38]. The discovery of yet another new species from Nyungwe Forest and Cyamudongo Forest indicates that the herpetofaunal diversity of these forests is still not fully assessed. Future field surveys will probably yield additional records, especially in the vast unexplored areas that still remain in Nyungwe Forest. The discovery of another amphibian species that is locally endemic to Nyungwe National Park underlines the importance of the national park for the protection of the area’s biodiversity.
Funding
Visits of the RMCA were partly funded by the Synthesys Project (http://www.synthesys.info), which is financed by European Community Research Infrastructure Action under the FP6 “Structuring the European Research Area” programme (BE-TAF-2107, BE-TAF-4004).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are available upon request from the corresponding author.
Acknowledgments
The Rwanda Development Board (RDB; Antoine Mudakikwa and Richard Muvunyi) kindly issued research, collecting and export permits. Siegmar Seidel (Rwanda Centre, University of Koblenz), Ulrich Sinsch and Eberhard Fischer (Department of Biology, University of Koblenz), Bonny Dumbo (Cyamudongo Project, University of Koblenz), and Harald Hinkel (World Bank, Kigali) provided various help during field work in Rwanda. Garin Cael and Danny Meirte (RMCA), Alexander Kupfer (SMNS), Wolfgang Böhme and Morris Flecks (ZFMK), and Mark-Oliver Rödel and Frank Tillack (ZMB) provided working space and let me examine the material under their care. Axel Hochkirch, Michael Veith and Stefan Lötters (Department of Biogeography, University of Trier) kindly permitted the use of their laboratory facilities for the molecular analysis of tissue samples. Siegmar Seidel also kindly provided the map of Rwanda and Nyungwe National Park.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Table A1.
Specimens of Arthroleptis species examined for this study, their taxonomic status, voucher number, present-day country of origin and collection locality.
Table A1.
Specimens of Arthroleptis species examined for this study, their taxonomic status, voucher number, present-day country of origin and collection locality.
| Species | Status | Voucher # | Country of Origin | Locality |
|---|---|---|---|---|
| A. adolfifriederici | syntype | ZMB 21787 | Rwanda | Rugegewald [=northeastern Nyungwe Forest] |
| A. adolfifriederici | topotype | 201511 | Rwanda | Rugegewald [=northeastern Nyungwe Forest] |
| A. adolfifriederici | - | JMD 597, 202103 | Rwanda | Nyungwe Forest |
| A. adolfifriederici | - | JMD 201517–23 | Rwanda | Gishwati Forest |
| A. adolfifriederici | - | JMD 201634 | Rwanda | Cyamudongo Forest |
| A. hematogaster | possible types | RMCA “Caja 40, Lote 60” (2 specs.) | DRC | May ya Moto |
| A. loveridgei | syntypes | RMCA 2624–25 | DRC | Arebi |
| A. nyungwensis | holotype | ZFMK 104075 | Rwanda | Rukuzi (2.4635 S, 29.2293 E; 2200 m), Nyungwe National Park |
| A. nyungwensis | paratype | ZFMK 104076 | Rwanda | Rukuzi (2.4635 S, 29.2293 E; 2200 m), Nyungwe National Park |
| A. nyungwensis | paratype | ZFMK 104077 | Rwanda | Cyamudongo Forest (2.546 S, 28.989 E, about 2000 m) |
| A. nyungwensis | paratype | SMNS 15743–44 | Rwanda | Cyamudongo Forest (2.5378 S, 29.9942 E; 1830 m) |
| A. phrynoides | holotype | RMCA 75-43-B-23 | DRC | Lomami, Terr. de Lomela, Sankuru |
| A. pyrrhoscelis | possible types | RMCA “Caja 9, Lote 98” (5 females, 2 males) | DRC | Haute Lubitshako, Terr. de Fizi (Kabobo) |
| A. pyrrhoscelis | possible types | RMCA “Caja 39, Lote 64” (many specs.) | DRC | Haute Lubitshako, Terr. de Fizi (Kabobo) |
| A. schubotzi | holotype | ZMB 21774 | Burundi | Usumbura [=Bujumbura] |
| A. schubotzi | - | ZMB 22860 | DRC | Insel Kwidschwi |
| A. schubotzi | - | ZFMK 104078–104092, JMD 201909–10 | Rwanda | Cyamudongo Forest |
| A. schubotzi | - | ZFMK 104081,-84,-85 | Rwanda | Buhinga |
| A. schubotzi | syntypes of Schoutedenella kivuensis | RMCA 48.783–784 | DRC | N’Zulu (Lac Kivu)—Passe de Sake (alt. 1500 m.) |
| A. schubotzi | possible types of Schoutedenella discodactyla | RMCA “Caja 31, Lote 81” (2 specs.) | DRC | Lutunguru |
| A. spinalis | holotype | RMCA 455 | DRC | Plaine St. Louis (Baudounville) |
| A. spinalis | holotype of A. boulengeri | RMCA 534 | DRC | Plaine St. Louis (Baudounville) |
| A. vercammeni | - | RMCA “Caja 4, Lote 3” | DRC | Mwana |
| A. xenochirus | syntypes of Arthroleptis globosa | RMCA 586, B 585/A–F (7 specs.) | DRC | Lofoi |
| A. xenochirus | syntypes of Arthroleptis lameeri | RMCA 180–180D | DRC | Lofoi |
| A. xenochirus | syntypes of Schoutedenella muta | RMCA 16219– 16248 | DRC | Kando près Tenke |
Table A2.
Samples of species used for molecular genetic analyses, their geographic origins, voucher specimens, GenBank accession numbers, and original sources.
Table A2.
Samples of species used for molecular genetic analyses, their geographic origins, voucher specimens, GenBank accession numbers, and original sources.
| Species | Location | Voucher | GenBank# | Original Source |
|---|---|---|---|---|
| Arthroleptis adolfifriederici | Nyungwe Forest, Rwanda | JMD 201511 | OR656715 | this study |
| A. adolfifriederici | Gishwati Forest, Rwanda | JMD 201517 | OR656716 | this study |
| JMD 201518 | OR656717 | |||
| JMD 201521 | OR656718 | |||
| JMD 201522 | OR656719 | |||
| A. adolfifriederici | Cyamudongo Forest, Rwanda | JMD 201634 | OR656720 | this study |
| A. nyungwensis | Rukuzi, Nyungwe National Park, Western Province, Rwanda | ZFMK 104075 | OR656721 | this study |
| ZFMK 104076 | OR656722 | |||
| A. nyungwensis | Cyamudongo Forest (2.5378 S, 29.9942 E; 1830 m), Western Province, Rwanda | SMNS 15743 | OR656723 | this study |
| SMNS 15744 | OR656724 | |||
| A. schubotzi | Cyamudongo Forest, Rwanda | ZFMK 104079 | OR656725 | this study |
| ZFMK 104087 | OR656726 | |||
| ZFMK 104088 | OR656727 | |||
| A. schubotzi | Buhinga, Rwanda | ZFMK 104081 | OR656728 | this study |
| ZFMK 104084 | OR656729 | |||
| ZFMK 104085 | OR656730 | |||
| A. schubotzi | Bwindi Impenetrable National Park, Uganda | CAS 201752 | DQ283237 | [12] |
| A. xenochirus | Lukwakwa, Zambia | BMNH 2018.5813 | MK464472 | [39] |
| A. xenodactyloides | Angola | UF185602 | MK036432 | [40] |
| A. xenodactyloides | Misuku Mountains, Mughese Forest Reserve, Malawi | MCZ:Herp:A-137057 | FJ151103 | [6] |
| Leptopelis karissimbensis | Rwasenkoko swamp, Nyungwe National Park, Rwanda | JMD 631 | KT439189 | [41] |
References
- Channing, A.; Rödel, M.-O. Field Guide to the Frogs & Other Amphibians of Africa; Penguin Random House: Cape Town, South Africa, 2019. [Google Scholar]
- Frost, D.R. Amphibian Species of the World: An Online Reference. Version 6.2. Electronic Database; American Museum of Natural History: New York, NY, USA, 2023; Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 20 September 2023).
- Laurent, R.F. Remarques sur le genre Schoutedenella Witte. Ann. Musée R. Congo Belg. Sci. Zool. 1954, 1, 34–40. [Google Scholar]
- Laurent, R.F. Amphibiens. In Exploration du Parc National des Virunga; Deuxième Series; Fondation pour Favoriser les Recherches Scientifique en Afrique: Bruxelles, Belgium, 1972; Volume 22, pp. 1–125. [Google Scholar]
- Schmidt, K.; Inger, R. Amphibians exclusive of the genera Afrixalus and Hyperolius. In Exploration du Parc National de l’Upemba; Institut des Parcs Nationaux du Congo Belge: Bruxelles, Belgium, 1959; Volume 56, pp. 1–294. [Google Scholar]
- Blackburn, D.C. Biogeography and evolution of body size and life history of African frogs: Phylogeny of squeakers (Arthroleptis) and long-fingered frogs (Cardioglossa) estimated from mitochondrial data. Mol. Phylogenetics Evol. 2008, 49, 806–826. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, D.C. Description and phylogenetic relationships of two new species of miniature Arthroleptis (Anura: Arthroleptidae) from the Eastern Arc Mountains of Tanzania. Breviora 2009, 517, 1–17. [Google Scholar] [CrossRef]
- de Witte, G.-F. Description de batraciens nouveaux du Congo Belge. Rev. Zool. Afr. 1921, 9, 1–27. [Google Scholar]
- Laurent, R.F. The natural classification of the Arthroleptinae (Amphibia, Hyperoliidae). Rev. Zool. Bot. Afr. 1973, 87, 666–678. [Google Scholar]
- Loveridge, A. Check list of the reptiles and amphibians of East Africa (Uganda; Kenya; Tanganyika; Zanzibar). Bull. Mus. Comp. Zool. Harv. Coll. 1957, 117, 151–362. [Google Scholar]
- Poynton, J.C.; Broadley, D.G. Amphibia Zambesiaca 1. Scolecomorphidae, Pipidae, Microhylidae, Hemisidae, Arthroleptidae. Ann. Natal Mus. 1985, 26, 503–553. [Google Scholar]
- Frost, D.R.; Grant, T.; Faivovich, J.; Bain, R.H.; Haas, A.; Haddad, C.F.B.; de Sá, R.O.; Channing, A.; Wilkinson, M.; Donnellan, S.C.; et al. The amphibian tree of life. Bull. Am. Mus. Nat. Hist. 2006, 297, 1–370. [Google Scholar] [CrossRef]
- Nieden, F. Neue ostafrikanische Frösche aus dem Kgl. Zool. Museum in Berlin. Sitzungsberichte Der Ges. Naturforschender Freunde Zu Berl. 1911, 1910, 436–441. [Google Scholar]
- Boulenger, G.A. Descriptions d’un ophidien et d’un batracien nouveaux du Congo. Rev. Zool. Afr. 1919, 7, 186–187. [Google Scholar]
- de Witte, G.F. Batraciens et reptiles. In Exploration du Parc National Albert, Mission G. F. de Witte (1933–1935), Fascicle 33; Fondation pour Favoriser les Recherches Scientifiques en Afrique: Bruxelles, Belgium, 1941; 261p. [Google Scholar]
- Laurent, R. Reptiles et batraciens nouveaux du massif du mont Kabobo et du plateau des Marungu. Rev. De Zool. Et De Bot. Afr. 1952, 46, 18–34. [Google Scholar]
- Dehling, J.M.; Sinsch, U. Amphibians of Rwanda: Diversity, community features, and conservation status. Diversity 2023, 15, 512. [Google Scholar] [CrossRef]
- Nieden, F. Neues Verzeichnis der Kriechtiere (außer den Schlangen) von Deutsch-Ostafrika. II. Teil: Amphibia. Mitteilungen Aus Dem Zool. Mus. Berl. 1915, 7, 345–390. [Google Scholar]
- Hinkel, H.; Fischer, E. Checklist of amphibians and reptiles of Nyungwe Forest, Rwanda. Tauraco Res. Rep. 1990, 3, 135–138. [Google Scholar]
- Nieden, F. Amphibia. In Wissenschaftliche Ergebnisse der Deutschen Zentral-Afrika-Expedition 1907–1908 unter Führung Adolf Friedrichs, Herzogs zu Mecklenburg; Band, I.V., Zoologie, I.I., Schubotz, H., Eds.; Klinkhardt & Biermann: Leipzig, Germany, 1912; Volume IV, pp. 165–196. [Google Scholar]
- de Witte, G.-F. Batraciens nouveaux du Congo Belge. Rev. Zool. Bot. Afr. 1933, 24, 97–103. [Google Scholar] [CrossRef]
- Channing, A.; Howell, K.M. Amphibians of East Africa; Edition Chimaira: Frankfurt, Germany, 2006. [Google Scholar]
- Palumbi, S. Nucleic acids II: The polymerase chain reaction. In Molecular Systematics; Hillis, D.M., Moritz, C., Mable, B.K., Eds.; Sinauer Associates: Sunderland, MA, USA, 1996; pp. 205–247. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and high-performance computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Köhler, J.; Jansen, M.; Rodríguez, A.; Kok, P.J.R.; Toledo, L.F.; Emmrich, M.; Glaw, F.; Haddad, C.F.B.; Rödel, M.-O.; Vences, M. The use of bioacoustics in anuran taxonomy: Theory, terminology, methods and recommendations for best practice. Zootaxa 2017, 4251, 1–124. [Google Scholar] [CrossRef]
- Zimkus, B.M.; Blackburn, D.C. Distinguishing features of the sub-Saharan frog genera Arthroleptis and Phrynobatrachus: A short guide for field and museum researchers. Breviora 2008, 513, 1–12. [Google Scholar] [CrossRef]
- Dehling, D.M.; Dehling, J.M. Elevated alpha diversity in disturbed sites obscures regional decline and homogenization of amphibian taxonomic, functional and phylogenetic diversity. Sci. Rep. 2023, 13, 1710. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, V. Amphibians of Malawi. In An Analysis of Their Richness and Community Diversity in a Changing Landscape; Edition Chimaira: Frankfurt am Main, Germany, 2011. [Google Scholar]
- Pickersgill, M. Frog Search. Results of Expeditions to Southern and Eastern Africa; Edition Chimaira: Frankfurt am Main, Germany, 2007. [Google Scholar]
- Maiditsch, I.; Liedtke, H.C.; Ng’wava, J.M.; Hödl, W. Advertisement and close-range encounter call of Arthroleptis schubotzi Nieden, 1911, with notes on phonotaxis and sexual dimorphism in the third manual digit (Anura: Arthroleptidae). Herpetozoa 2011, 24, 23–31. [Google Scholar]
- Wells, K.D. The Ecology and Behavior of Amphibians; The University of Chicago Press: Chicago, IL, USA; London, UK, 2007. [Google Scholar]
- Dehling, J.M. An African glass frog: A new Hyperolius species (Anura: Hyperoliidae) from Nyungwe National Park, southern Rwanda. Zootaxa 2012, 3391, 52–64. [Google Scholar] [CrossRef]
- Channing, A.; Hillers, A.; Lötters, S.; Rödel, M.O.; Schick, S.; Conradie, W.; Rödder, D.; Mercurio, V.; Wagner, P.; Dehling, J.M.; et al. Taxonomy of the super-cryptic Hyperolius nasutus group of long reed frogs of Africa (Anura: Hyperoliidae), with descriptions of six new species. Zootaxa 2013, 3620, 301–350. [Google Scholar] [CrossRef]
- Hughes, D.F.; Tolley, K.A.; Behangana, M.; Lukwago, W.; Menegon, M.; Dehling, J.M.; Stipala, J.; Tilbury, C.R.; Khan, A.M.; Kusamba, C.; et al. Cryptic diversity in Rhampholeon boulengeri (Sauria: Chamaeleonidae), a pygmy chameleon from the Albertine Rift biodiversity hotspot. Mol. Phylogenetics Evol. 2018, 122, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Dehling, J.M.; Sinsch, U. Partitioning of morphospace in larval and adult reed frogs (Anura: Hyperoliidae: Hyperolius) of the Central African Albertine Rift. Zool. Anz. 2019, 280, 65–77. [Google Scholar] [CrossRef]
- Greenbaum, E.; Portik, D.M.; Allen, K.E.; Vaughan, E.R.; Badjedjea, G.; Barej, M.F.; Behangana, M.; Conkey, N.; Dumbo, B.; Gonwouo, L.N.; et al. Systematics of the Central African spiny reed frog Afrixalus laevis (Anura: Hyperoliidae), with the description of two new species from the Albertine Rift. Zootaxa 2022, 5174, 201–232. [Google Scholar] [CrossRef]
- Bittencourt-Silva, G.B. A herpetological survey of western Zambia. Amphib. Reptile Conserv. 2019, 13, 1–28. [Google Scholar]
- Hayes, D.T.; Ceriaco, L.M.P.; Jongsma, G.F.M.; Bandeira, S.; Hedley, R.W.; Stanley, E.L.; Bauer, A.M.; Blackburn, D.C. Amphibian Surveys at Two Important Type Localities in Malanje Province, Angola. Available online: https://www.ncbi.nlm.nih.gov/nuccore/MK036432 (accessed on 20 September 2023).
- Lehr, E.; Dehling, J.M.; Greenbaum, E.; Sinsch, U. Embryogenesis and tadpole description of Hyperolius castaneus Ahl, 1931 and H. jackie Dehling, 2012 (Anura, Hyperoliidae) from montane bog pools. ZooKeys 2015, 546, 125–152. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).