
Assessment of Anthropogenic Impacts on the Genetic Diversity of Phragmites australis in Small-River Habitats

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
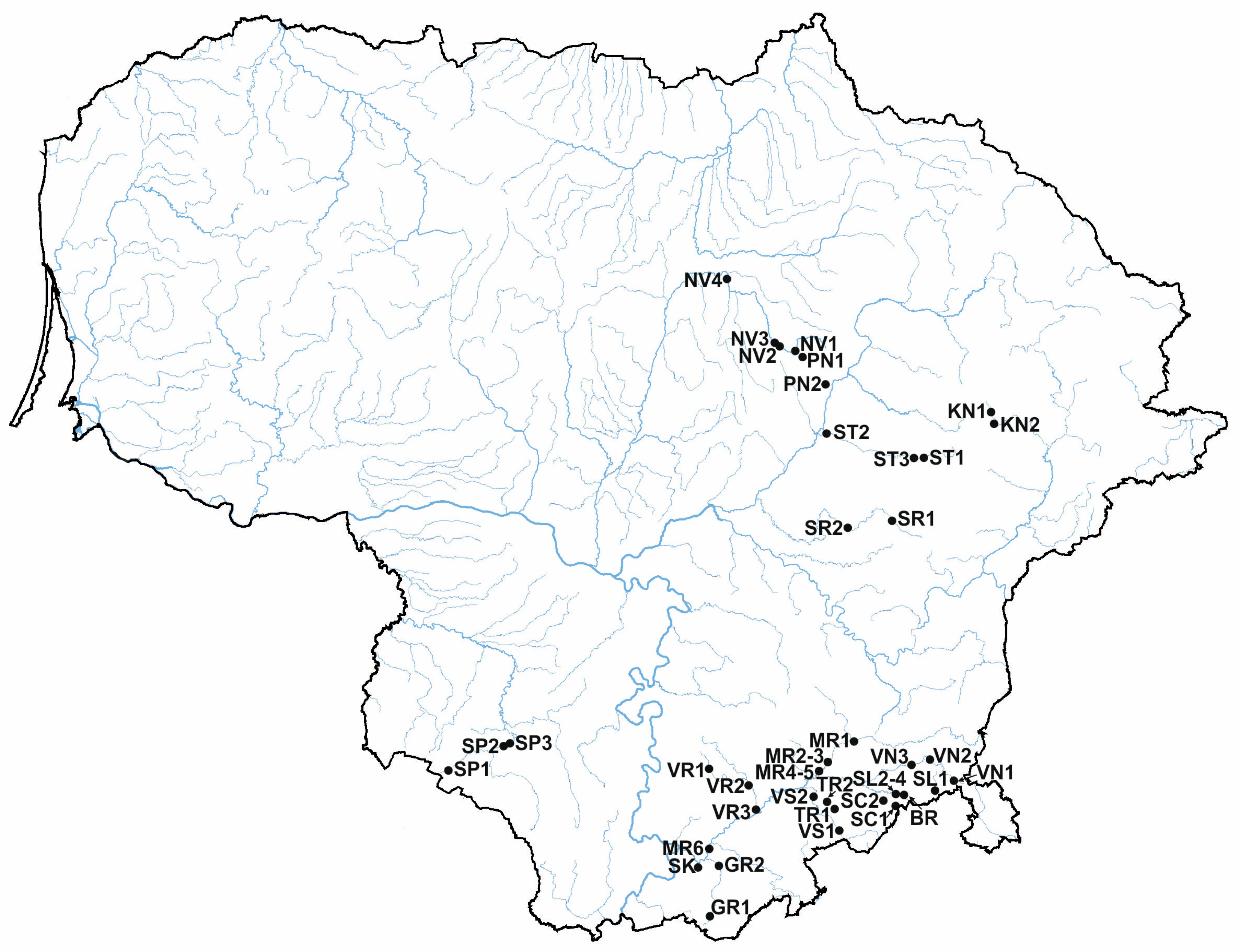
2.1. Sampling Sites and Common Reed Samples
2.2. DNA Extraction
2.3. ISSR Marker Analysis
2.4. Data Analysis
3. Results
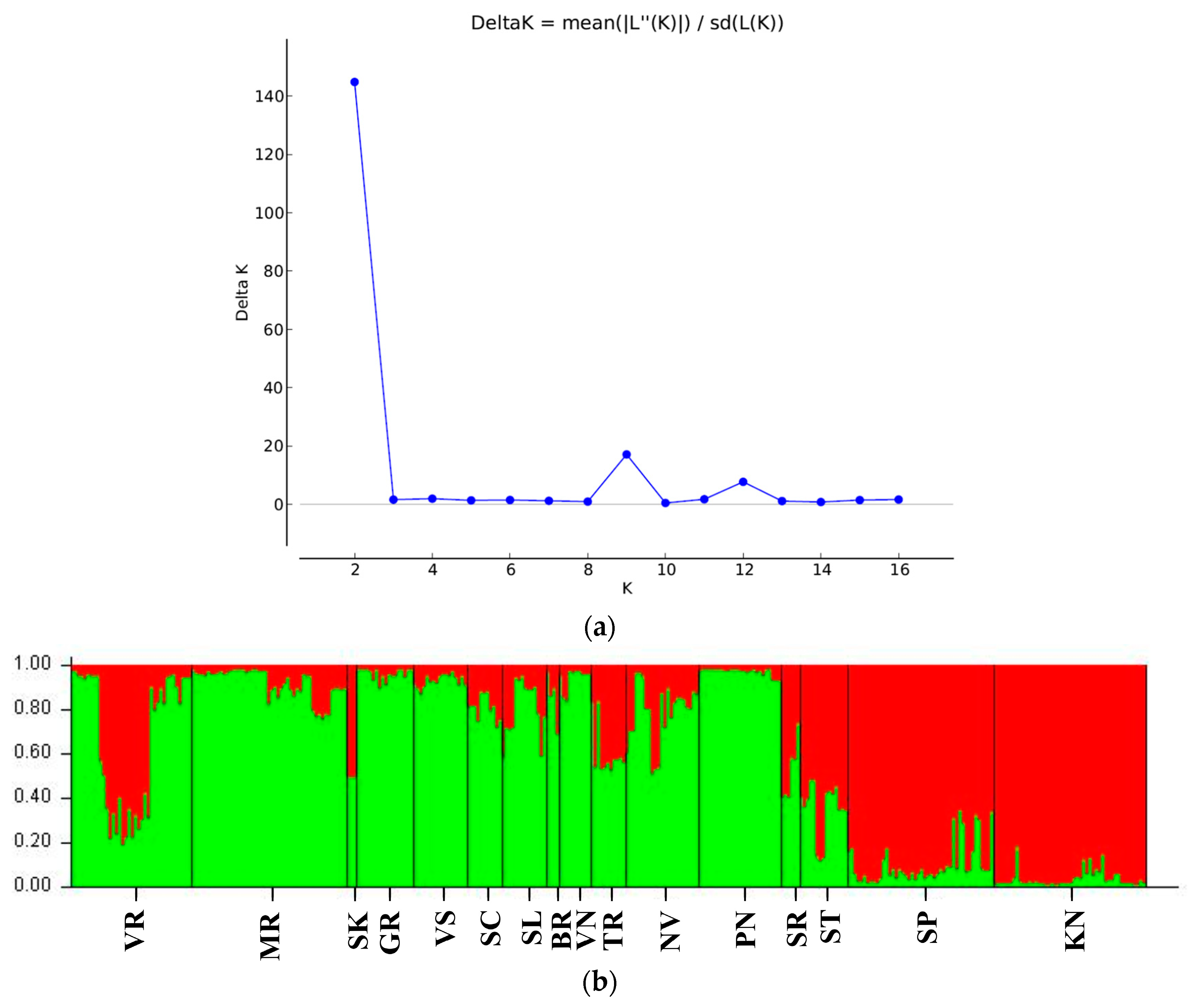
3.1. Genetic Diversity Based on ISSR Markers
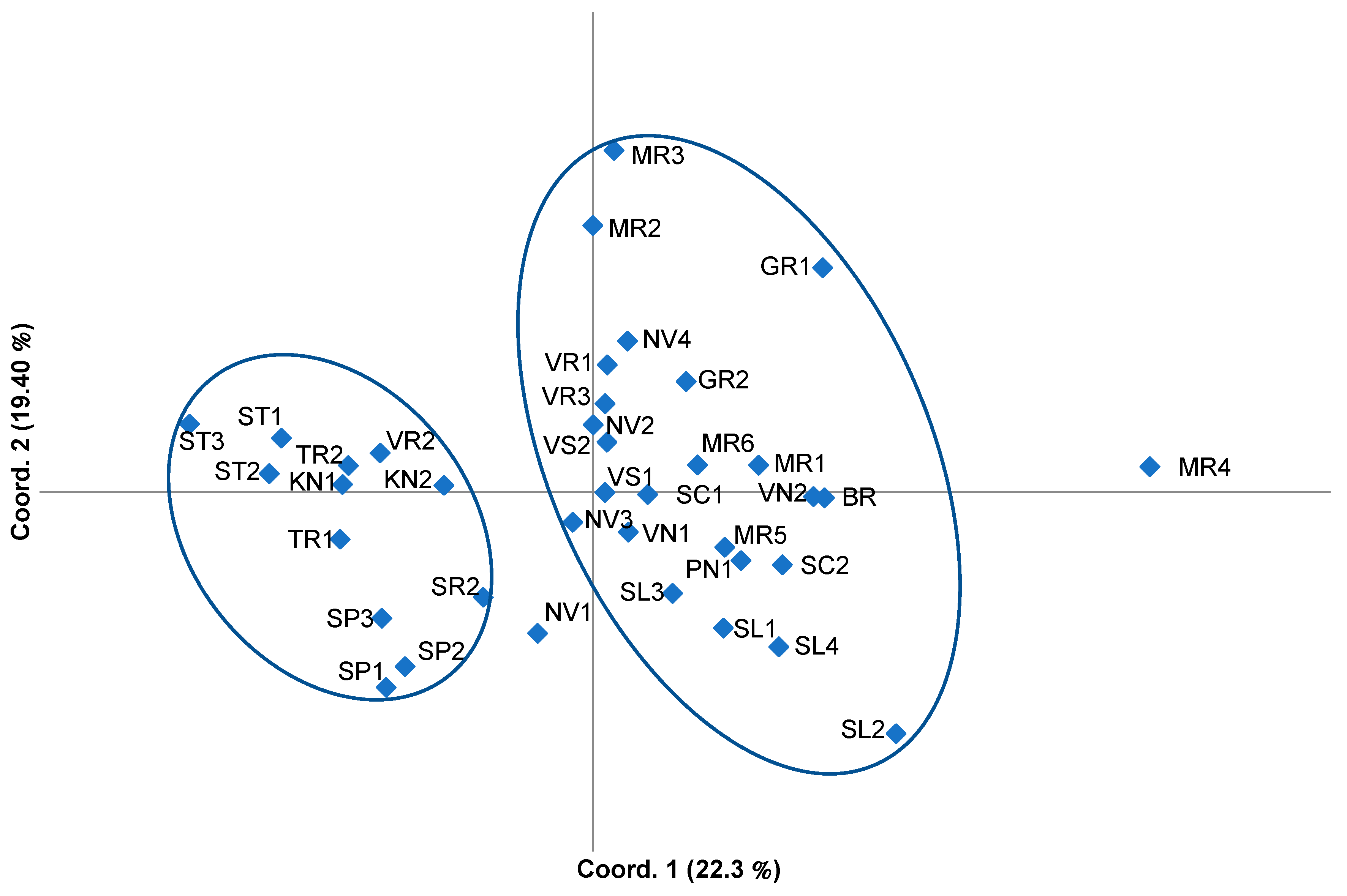
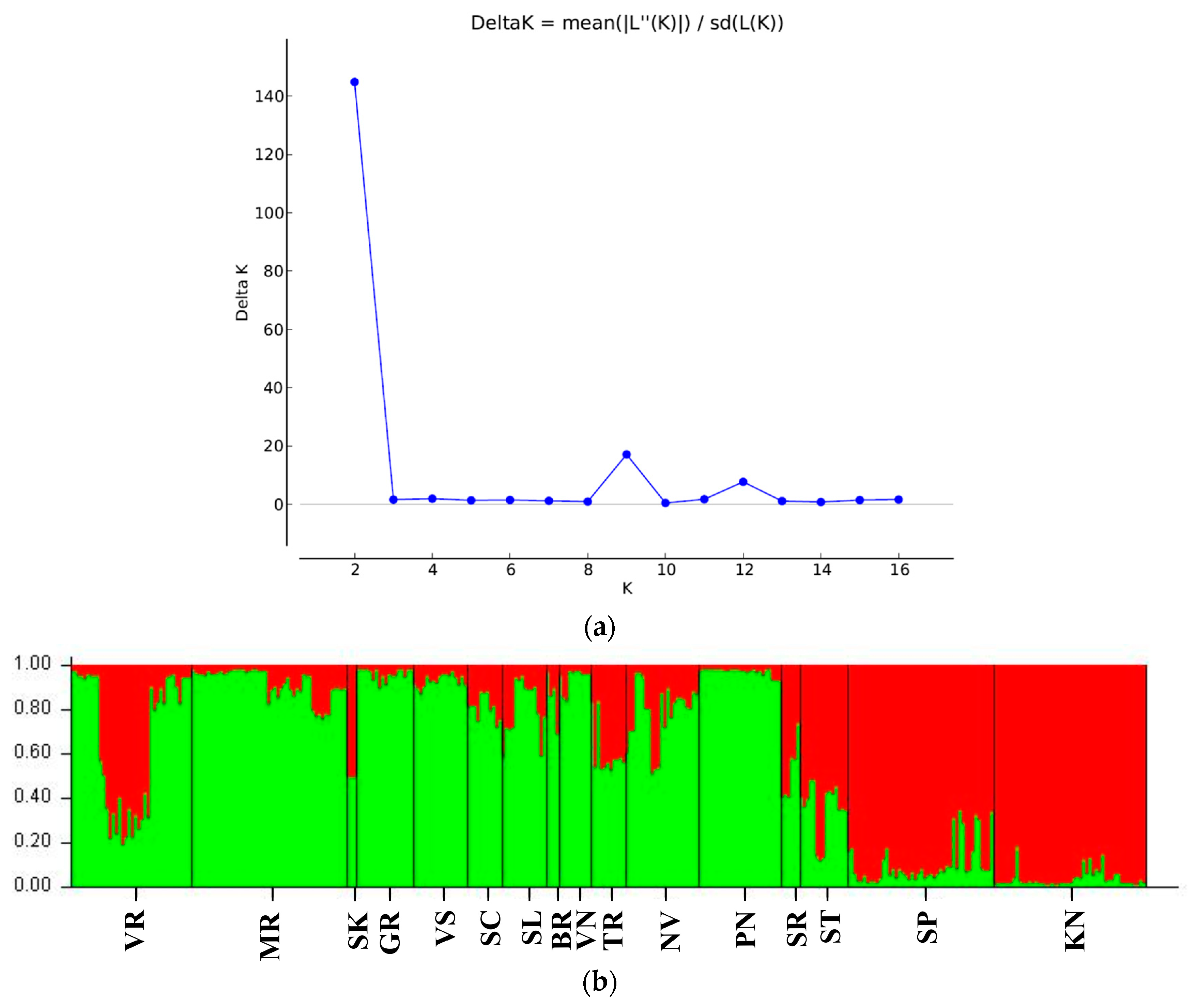
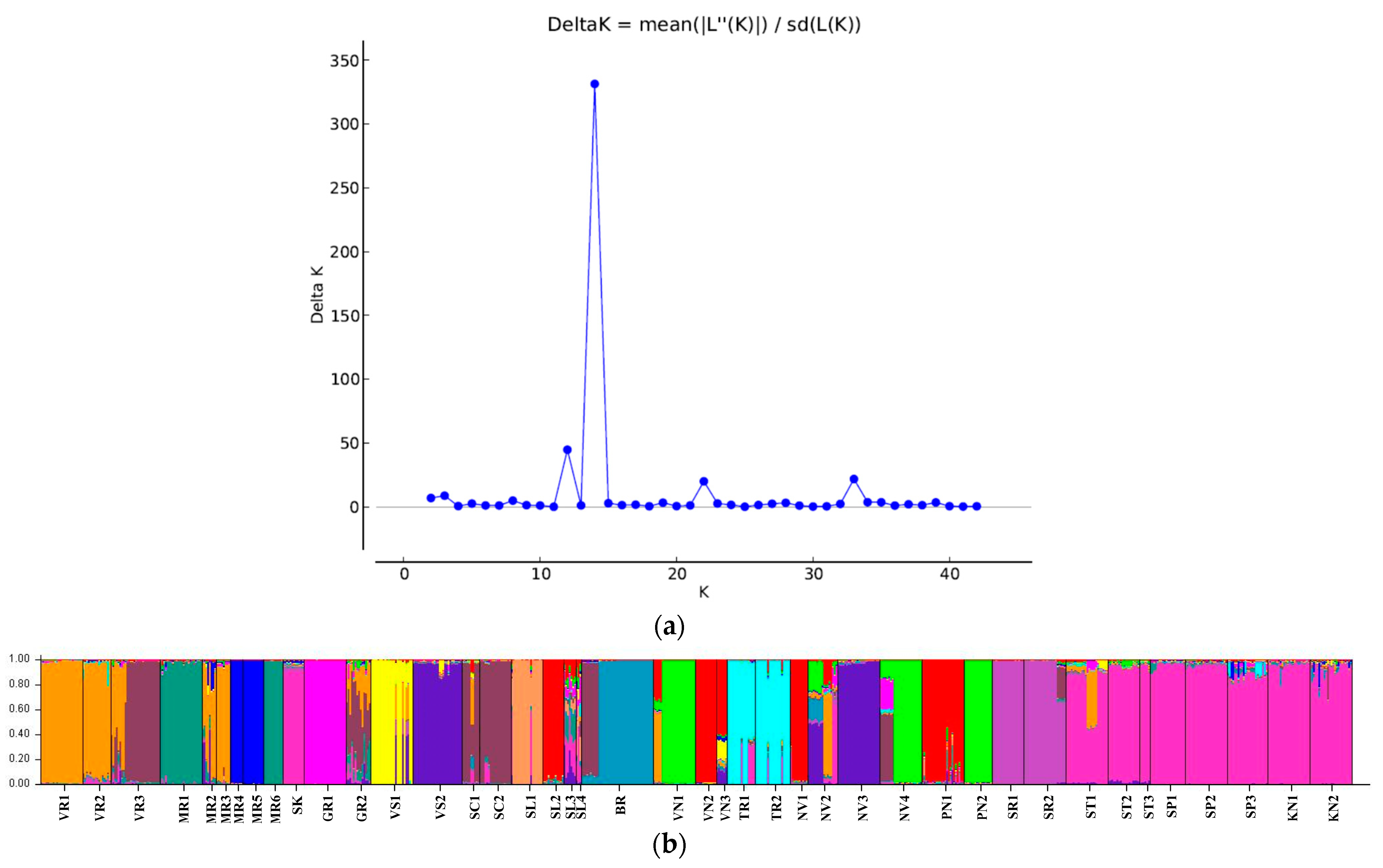
3.2. Genetic Differentiation of Sites and Populations
3.3. Assessment of Environmental and Anthropogenic Impacts on Genetic Diversity
4. Discussion
4.1. Genetic Structure of Populations and Sites
4.2. Eutrophication Impact
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hoban, S.; Archer, F.I.; Bertola, L.D.; Bragg, J.G.; Breed, M.F.; Bruford, M.W.; Coleman, M.A.; Ekblom, R.; Funk, W.C.; Grueber, C.E.; et al. Global genetic diversity status and trends: Towards a suite of Essential Biodiversity Variables (EBVs) for genetic composition. Biol. Rev. 2022, 97, 1511–1538. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Roger, F.; Godhe, A.; Gamfeldt, L. Genetic diversity and ecosystem functioning in the face of multiple stressors. PLoS ONE 2012, 7, e45007. [Google Scholar] [CrossRef] [PubMed]
- Hautier, Y.; Tilman, D.; Isbell, F.; Seabloom, E.W.; Borer, E.T.; Reich, P.B. Anthropogenic environmental changes impact ecosystem stability via biodiversity. Science 2015, 348, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Leimu, R.; Mutikainen, P.; Koricheva, J.; Fisher, M. How general are positive relationship between plant population size, fitness, and genetic variation? J. Ecol. 2006, 94, 942–952. [Google Scholar] [CrossRef]
- Sebbenn, A.M.; Carvalho, A.C.M.; Freitas, M.L.M.; Moraes, S.M.B.; Gaino, A.P.S.C.; daSilva, J.M.; Jolivet, C.; Moraes, M.L.T. Low levels of realized seed and pollen gene flow and strong spatial genetic structure in a small, isolated and fragmented population of the tropical tree Copaifera langsdorffii Desf. Heredity 2011, 106, 134–145. [Google Scholar] [CrossRef]
- Beatty, G.E.; Reid, N.; Provan, J. Retrospective genetic monitoring of the threatened yellow marsh saxifrage (Saxifraga hirculus) reveals genetic erosion but provides valuable insights for conservation strategies. Divers. Distrib. 2014, 20, 529–537. [Google Scholar] [CrossRef]
- Sampson, J.F.; Hankinson, M.; McArthur, S.; Tapper, S.; Langley, M.; Gibson, N.; Yates, C.; Byrne, M. Long-term ‘islands’ in the landscape: Low gene flow, effective population size and genetic divergence in the shrub Hakea oldfieldii (Proteaceae). Bot. J. Linn. Soc. 2015, 179, 319–334. [Google Scholar] [CrossRef]
- Banks, S.C.; Cary, G.J.; Smith, A.L.; Davies, I.D.; Driscoll, D.A.; Gill, A.M.; Lindenmayer, D.B.; Peakall, R. How does ecological disturbance influence genetic diversity? Trends Ecol. Evol. 2013, 28, 670–679. [Google Scholar] [CrossRef]
- Fuller, M.R.; Doyle, M.W.; Strayer, D.L. Causes and consequences of habitat fragmentation in river networks. Ann. N. Y. Acad. Sci. 2015, 1355, 31–51. [Google Scholar] [CrossRef]
- Kiviat, E. Ecosystem services of Phragmites in North America with emphasis on habitat functions. AoB Plants 2013, 5, plt008. [Google Scholar] [CrossRef]
- Engloner, A.I.; Szegő, D. Genetic diversity of riverine reed stands indicating the water regime of the habitat. Ecol. Indic. 2016, 61, 846–849. [Google Scholar] [CrossRef]
- Kirk, H.; Paul, J.; Straka, J.; Freeland, J.R. Long distance dispersal and genetic diversity are implicated in the invasive spread of the common reed Phragmites australis (Poaceae) in Eastern North America. Am. J. Bot. 2011, 98, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Lambertini, C.; Sorrell, B.K.; Riis, T.; Olesen, B.; Brix, H. Exploring the borders of European Phragmites within a cosmopolitan genus. AoB Plants 2012, 2012, pls020. [Google Scholar] [CrossRef] [PubMed]
- Eller, F.; Skálová, H.; Caplan, J.S.; Bhattarai, G.P.; Burger, M.K.; Cronin, J.T.; Guo, W.-Y.; Guo, X.E.; Hazelton, L.G.; Kettenring, K.M.; et al. Cosmopolitan species as models for ecophysiological responses to global change: The common reed Phragmites australis. Front. Plant Sci. 2017, 8, 1833. [Google Scholar] [CrossRef] [PubMed]
- Saltonstall, K. Cryptic invasion by a non-native genotype of Phragmites australis into North America. Proc. Natl. Acad. Sci. USA 2002, 99, 2445–2449. [Google Scholar] [CrossRef] [PubMed]
- Kettenring, K.M.; McCormick, M.K.; Baron, H.M.; Whigham, D.F. Mechanisms of Phragmites australis invasion: Feedbacks among genetic diversity, nutrients, and sexual reproduction. J. Appl. Ecol. 2011, 48, 1305–1313. [Google Scholar] [CrossRef]
- Guo, W.Y.; Lambertini, C.; Pyšek, P.; Meyerson, L.A.; Brix, H. Living in two worlds: Evolutionary mechanisms act differently in the native and introduced ranges of an invasive plant. Ecol. Evol. 2018, 8, 2440–2452. [Google Scholar] [CrossRef] [PubMed]
- Cronk, J.K.; Fennessy, M.S. Wetland Plants: Biology and Ecology; CRC Press/Lewis Publishers: Boca Raton, FL, USA, 2001; pp. 147–188. [Google Scholar]
- McCormick, M.K.; Kettenring, K.M.; Baron, H.M.; Whigham, D.F. Extent and reproductive mechanisms of Phragmites australis spread in brackish wetlands in Chesapeake Bay, Maryland (USA). Wetlands 2010, 30, 67–74. [Google Scholar] [CrossRef]
- Ishii, J.; Kadono, Y. Factors influencing seed production of Phragmites australis. Aquat. Bot. 2002, 72, 129–141. [Google Scholar] [CrossRef]
- Brochet, A.L.; Guillemain, M.; Fritz, H.; Gauthier-Clerc, M.; Green, A.J. The role of migratory ducks in the long-distance dispersal of native plants and the spread of exotic plants in Europe. Ecography 2009, 32, 919–928. [Google Scholar] [CrossRef]
- Fér, T.; Hroudová, Z. Genetic diversity and dispersal of Phragmites australis in a small river system. Aquat. Bot. 2009, 90, 165–171. [Google Scholar] [CrossRef]
- Stabile, J.; Lipus, D.; Maceda, L.; Maltz, M.; Roy, N.; Wirgin, I. Microsatellite DNA analysis of spatial and temporal population structuring of Phragmites australis along the Hudson River Estuary. Biol. Invasions 2016, 18, 2517–2529. [Google Scholar] [CrossRef]
- Kuprina, K.; Seeber, E.; Schnittler, M.; Landeau, R.; Lambertini, C.; Bog, M. Genetic diversity of common reed in the Southern Baltic Sea region—Is there an influence of disturbance? Aquat. Bot. 2022, 177, 103471. [Google Scholar] [CrossRef]
- Kettenring, K.M.; McCormick, M.K.; Baron, H.M.; Whigham, D.F. Phragmites australis (common reed) invasion in the Rhode River subestuary of the Chesapeake Bay: Disentangling the effects of foliar nutrients, genetic diversity, patch size, and seed viability. Estuaries Coast. 2010, 33, 118–126. [Google Scholar] [CrossRef]
- Belzile, F.; Labbé, J.; LeBlanc, M.C.; Lavoie, C. Seeds contribute strongly to the spread of the invasive genotype of the common reed (Phragmites australis). Biol. Invasions 2010, 12, 2243–2250. [Google Scholar] [CrossRef]
- Pyšek, P.; Skálová, H.; Čuda, J.; Guo, W.-Y.; Suda, J.; Doležal, J.; Kauzál, O.; Lambertini, C.; Lučanová, M.; Mandáková, T.; et al. Small genome separates native and invasive populations in an ecologically important cosmopolitan grass. Ecology 2018, 99, 79–90. [Google Scholar] [CrossRef]
- Oh, D.-H.; Kowalski, K.P.; Quach, Q.N.; Wijesinghege, C.; Tanford, P.; Dassanayake, M.; Clay, K. Novel genome characteristics contribute to the invasiveness of Phragmites australis (common reed). Mol. Ecol. 2022, 31, 1142–1159. [Google Scholar] [CrossRef]
- Tao, F.; Fan, C.; Liu, Y.; Sivakumar, S.; Kowalski, K.P.; Golenberg, E.M. Optimization and application of non-native Phragmites australis transcriptome assemblies. PLoS ONE 2023, 18, e0280354. [Google Scholar] [CrossRef]
- Coppi, A.; Lastrucci, L.; Cappelletti, D.; Cerri, M.; Ferranti, F.; Ferri, V.; Foggi, B.; Gigante, D.; Venanzoni, R.; Viciani, D.; et al. AFLP approach reveals variability in Phragmites australis: Implications for its dieback and evidence for genotoxic effects. Front. Plant Sci. 2018, 9, 386. [Google Scholar] [CrossRef]
- Erős, T.; Kuehne, L.; Dolezsai, A.; Sommerwerk, N.; Wolter, C. A systematic review of assessment and conservation management in large floodplain rivers—Actions postponed. Ecol. Indic. 2019, 98, 453–461. [Google Scholar] [CrossRef]
- Krokaitė, E.; Shakeneva, D.; Juškaitytė, E.; Rekašius, T.; Nemaniūtė-Gužienė, J.; Butkuvienė, J.; Patamsytė, J.; Rančelienė, V.; Vyšniauskienė, R.; Duchovskienė, L.; et al. Nitrogen concentration of the aquatic plant species in relation to land cover type and other variables of the environment. Zemdirb. Agric. 2019, 106, 203–212. [Google Scholar] [CrossRef]
- Liu, L.; Wang, J.; Ma, X.; Li, M.; Guo, X.; Yin, M.; Cai, Y.; Yu, X.; Du, N.; Wang, R.; et al. Impacts of the Yellow River and Qingtongxia dams on genetic diversity of Phragmites australis in Ningxia Plain, China. Aquat. Bot. 2021, 169, 103341. [Google Scholar] [CrossRef]
- Naugžemys, D.; Lambertini, C.; Patamsytė, J.; Butkuvienė, J.; Khasdan, V.; Žvingila, D. Genetic diversity patterns in Phragmites australis populations in straightened and in natural river sites in Lithuania. Hydrobiologia 2021, 848, 3317–3330. [Google Scholar] [CrossRef]
- Lambertini, C.; Gustafsson, M.H.G.; Frydenberg, J.; Speranza, M.; Brix, H. Genetic diversity patterns in Phragmites australis at the population, regional and continental scales. Aquat. Bot. 2008, 88, 160–170. [Google Scholar] [CrossRef]
- McCormick, M.K.; Kettenring, K.M.; Baron, H.M.; Whigham, D.F. Spread of invasive Phragmites australis in estuaries with differing degrees of development: Genetic patterns, Allee effects and interpretation. J. Ecol. 2010, 98, 1369–1378. [Google Scholar] [CrossRef]
- Anderson, N.O.; Jocienė, L.; Krokaitė, E.; Rekašius, T.; Paulauskas, A.; Kupčinskienė, E. Genetic diversity of Phalaris arundinacea populations in relation to river regulation in the Merkys basin, Lithuania. River Res. Appl. 2018, 34, 300–309. [Google Scholar] [CrossRef]
- Koppitz, H.; Kühl, H. To the importance of genetic diversity of Phragmites australis in the development of reed stands. Wetl. Ecol. Manag. 2000, 8, 403–414. [Google Scholar] [CrossRef]
- Engloner, A.I.; Major, Á. Clonal diversity of Phragmites australis propagating along water depth gradient. Aquat. Bot. 2011, 94, 172–176. [Google Scholar] [CrossRef]
- Hazelton, E.L.G.; McCormick, M.K.; Sievers, M.; Kettenring, K.M.; Whigham, D.F. Stand age is associated with clonal diversity, but not vigor, community structure, or insect herbivory in Chesapeake Bay Phragmites australis. Wetlands 2015, 35, 877–888. [Google Scholar] [CrossRef]
- Paul, J.; Kirk, H.; Freeland, J.R. Genetic diversity and differentiation of fragmented reedbeds (Phragmites australis) in the United Kingdom. Hydrobiologia 2011, 665, 107–115. [Google Scholar] [CrossRef]
- Vymazal, J.; Březinová, T. Accumulation of heavy metals in aboveground biomass of Phragmites australis in horizontal flow constructed wetlands for wastewater treatment: A review. Chem. Eng. J. 2016, 290, 232–242. [Google Scholar] [CrossRef]
- Uddin, M.N.; Robinson, R.W. Can nutrient enrichment influence the invasion of Phragmites australis? Sci. Total. Environ. 2018, 613–614, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Prica, M.; Andrejić, G.; Šinžar-Sekulić, J.; Rakić, T.; Dželetović, Ž. Bioaccumulation of heavy metals in common reed (Phragmites australis) growing spontaneously on highly contaminated mine tailing ponds in Serbia and potential use of this species in phytoremediation. Bot. Serbica 2019, 43, 85–95. [Google Scholar] [CrossRef]
- Ahmad, S.S.; Reshi, Z.A.; Shah, M.A.; Rashid, I.; Ara, R.; Andrabi, S.M.A. Phytoremediation potential of Phragmites australis in Hokersar wetland—A Ramsar Site of Kashmir Himalaya. Int. J. Phytoremediation 2014, 16, 1183–1191. [Google Scholar] [CrossRef]
- Rodrigo, M.A.; Valentin, A.; Claros, J.; Moreno, L.; Segura, M.; Lassalle, M.; Vera, P. Assessing the effect of emergent vegetation in a surface-flow constructed wetland on eutrophication reversion and biodiversity enhancement. Ecol. Eng. 2018, 113, 74–87. [Google Scholar] [CrossRef]
- Carricondo, J.M.; Oliver-Villanueva, J.V.; Turégano, J.V.; González, J.A.; Mengual, J. Use of Phragmites australis for controlling phosphorus contamination in anthropogenic wetland ecosystems. Environ. Technol. 2021, 42, 3055–3064. [Google Scholar] [CrossRef]
- Bonanno, G.; Lo Giudice, R. Heavy metal bioaccumulation by the organs of Phragmites australis (common reed) and their potential use as contamination indicators. Ecol. Indic. 2010, 10, 639–645. [Google Scholar] [CrossRef]
- Chandra, R.; Yadav, S. Phytoremediation of CD, CR, CU, MN, FE, NI, PB and ZN from aqueous solution using Phragmites cummunis, Typha angustifolia and Cyperus esculentus. Int. J. Phytoremediation 2011, 13, 580–591. [Google Scholar] [CrossRef]
- Labra, M.; Di Fabio, T.; Grassi, F.; Regondi, S.M.G.; Bracale, M.; Vannini, C.; Agradi, E. AFLP analysis as biomarker of exposure to organic and inorganic genotoxic substances in plants. Chemosphere 2003, 52, 1183–1188. [Google Scholar] [CrossRef]
- Piqueras, J.; Klimes, L.; Redbo-Torstensson, P. Modelling the morphological response to nutrient availability in the clonal plant Trientalis europaea L. Plant Ecol. 1999, 141, 117–127. [Google Scholar] [CrossRef]
- Bai, W.M.; Sun, X.Q.; Wang, Z.W.; Li, L.H. Nitrogen addition and rhizome severing modify clonal growth and reproductive modes of Leymus chinensis population. Plant Ecol. 2009, 205, 13–21. [Google Scholar] [CrossRef]
- EPA (Environmental Protection Agency). Agriculture and Lithuanian Waters. Impact of Agricultural Activity on the Status of Lithuanian Rivers and Pollution Loads in the Baltic Sea. Vilnius. 41 (in Lithuanian). 2018. Available online: https://aaa.lrv.lt/lt/veiklos-sritys/vanduo/upes-ezerai-ir-tvenkiniai/valstybinis-upiu-ezeru-ir-tvenkiniu-monitoringas/upiu-monitoringo-rezultatai (accessed on 28 August 2023).
- Lambertini, C.; Guo, W.Y.; Ye, S.; Eller, F.; Guo, X.; Li, X.Z.; Sorrell, B.K.; Speranza, M.; Brix, H. Phylogenetic diversity shapes salt tolerance in Phragmites australis estuarine populations in East China. Sci. Rep. 2020, 10, 17645. [Google Scholar] [CrossRef] [PubMed]
- Tanhuanpää, P.; Erkkilä, M.; Kalendar, R.; Schulman, A.H.; Manninen, O. Assessment of genetic diversity in Nordic timothy (Phleum pratense L.). Hereditas 2016, 153, 5. [Google Scholar] [CrossRef] [PubMed]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR) anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Gentili, R.; Fenu, G.; Mattana, E.; Citterio, S.; De Mattia, F.; Bacchetta, G. Conservation genetics of two island endemic Ribes spp. (Grossulariaceae) of Sardinia: Survival or extinction? Plant Biol. 2015, 17, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.S.; Mendonça, D.; Bettencourt, S.X.; Borba, A.R.; Melo, C.; Baptista, C.; da Câmara Machado, A. Genetic diversity of an Azorean endemic and endangered plant species inferred from inter-simple sequence repeat markers. AoB Plants 2014, 6, plu034. [Google Scholar] [CrossRef]
- Cavanzón-Medrano, L.E.; Machkour-M’Rabet, S.; Chablé-Iuit, L.R.; Pozo, C.; Hénaut, Y.; Legal, L. Effect of climatic conditions and land cover on genetic structure and diversity of Eunica tatila (Lepidoptera) in the Yucatan Peninsula, Mexico. Diversity 2018, 10, 79. [Google Scholar] [CrossRef]
- Krokaitė, E.; Janulionienė, R.; Jocienė, L.; Rekašius, T.; Rajackaitė, G.; Paulauskas, A.; Marozas, V.; Kupčinskienė, E. Relating invasibility and invasiveness: Case study of Impatiens parviflora. Front. Ecol. Evol. 2022, 10, 845947. [Google Scholar] [CrossRef]
- Naugžemys, D.; Patamsytė, J.; Žilinskaitė, S.; Hoshino, Y.; Skridaila, A.; Žvingila, D. Genetic structure of native blue honeysuckle populations in the Western and Eastern Eurasian Ranges. Plants 2022, 11, 1480. [Google Scholar] [CrossRef]
- Lal, A.; Pant, M.; Pant, G.; Palni, L.M.S.; Kumar, A.; Kumar, G. ISSR marker assisted genetic diversity assessment in natural populations of two endemic orchids Aerides multiflora and Rhynchostylis retusa from Uttarakhand, India. S. Afr. J. Bot. 2023, 157, 151–158. [Google Scholar] [CrossRef]
- Saltonstall, K. Microsatellite variation within and among North American lineages of Phragmites australis. Mol. Ecol. 2003, 12, 1689–1702. [Google Scholar] [CrossRef] [PubMed]
- Patamsytė, J.; Naugžemys, D.; Čėsnienė, T.; Kleizaitė, V.; Demina, O.N.; Mikhailova, S.I.; Agafonov, V.A.; Žvingila, D. Evaluation and comparison of the genetic structure of Bunias orientalis populations in their native range and two non-native ranges. Plant Ecol. 2018, 219, 101–114. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Dorken, M.E.; Eckert, C.G. Severely reduced sexual reproduction in northern populations of a clonal plant, Decodon verticillatus (Lythraceae). J. Ecol. 2001, 89, 339–350. [Google Scholar] [CrossRef]
- Vekemans, X.; Beauwens, T.; Lemaire, M.; Roldán-Ruiz, I. Data from amplified fragment length polymorphism (AFLP) markers show indication of size homoplasy and of a relationship between degree of homoplasy and fragment size. Mol. Ecol. 2002, 11, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Yang, R.; Boyle, T. POPGENE: Microsoft Windows-Based Freeware for Population Genetic Analysis: Release 1.31; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Evanno, G.; Regnault, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Koppitz, H. Analysis of genetic diversity among selected populations of Phragmites australis world-wide. Aquat. Bot. 1999, 64, 209–221. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Honnay, O.; Looy, K.V.; Breyne, P. Spatiotemporal structure of genetic variation of a spreading plant metapopulation on dynamic riverbanks along the Meuse River. Heredity 2006, 96, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.N.; Mangel, M. Modeling investments in seed, clonal offspring, and translocation in a clonal plant. Ecology 1999, 80, 1202–1220. [Google Scholar] [CrossRef]
- Weppler, T.; Stocklin, J. Variation of sexual and clonal reproduction in the alpine Geum reptans in contrasting altitudes and successional stages. Basic Appl. Ecol. 2005, 6, 305–306. [Google Scholar] [CrossRef]
- Jampeetong, A.; Brix, H.; Kantawanichkul, S. Effects of inorganic nitrogen forms on growth, morphology, nitrogen uptake capacity and nutrient allocation of four tropical aquatic macrophytes (Salvinia cucullata, Ipomoea aquatica, Cyperus involucratus and Vetiveria zizanioides). Aquat. Bot. 2012, 97, 10–16. [Google Scholar] [CrossRef]
- Tho, B.T.; Lambertini, C.; Eller, F.; Brix, H.; Sorrell, B.K. Ammonium and nitrate are both suitable inorganic nitrogen forms for the highly productive wetland grass Arundo donax, a candidate species for wetland paludiculture. Ecol. Eng. 2017, 105, 379–386. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | River Site | Code | Samples | Coordinates | L | M | H | ||
|---|---|---|---|---|---|---|---|---|---|
| N | G | Latitude | Longitude | ||||||
| 1. | Varėnė 1 | VR1 | 24 | 9 | 54.391611 | 24.407747 | A | + | − |
| 2. | Varėnė 2 | VR2 | 16 | 16 | 54.329047 | 24.511211 | N | − | + |
| 3. | Varėnė 3 | VR3 | 28 | 13 | 54.250614 | 24.554161 | U | − | − |
| 4. | Merkys 1 | MR1 | 24 | 24 | 54.436978 | 24.982144 | A | + | + |
| 5. | Merkys 2 | MR2 | 8 | 7 | 54.409117 | 24.910669 | A | + | + |
| 6. | Merkys 3 | MR3 | 8 | 4 | 54.388197 | 24.893189 | N | − | + |
| 7. | Merkys 4 | MR4 | 7 | 3 | 54.337967 | 24.822267 | N | − | + |
| 8. | Merkys 5 | MR5 | 12 | 6 | 54.336086 | 24.808022 | N | − | + |
| 9. | Merkys 6 | MR6 | 11 | 5 | 54.118106 | 24.302706 | N | − | + |
| 10. | Skroblus | SK | 12 | 1 | 54.105942 | 24.279528 | N | − | + |
| 11. | Grūda 1 | GR1 | 24 | 4 | 54.022956 | 24.333617 | A | + | + |
| 12. | Grūda 2 | GR2 | 14 | 14 | 54.110042 | 24.352975 | N | − | + |
| 13. | Verseka 1 | VS1 | 24 | 4 | 54.180353 | 24.949422 | A | + | + |
| 14. | Verseka 2 | VS2 | 28 | 13 | 54.311475 | 24.815764 | A | − | + |
| 15. | Šalčykščia 1 | SC1 | 10 | 3 | 54.255969 | 25.214581 | N | − | − |
| 16. | Šalčykščia 2 | SC2 | 18 | 8 | 54.266883 | 25.178094 | N | + | − |
| 17. | Šalčia 1 | SL1 | 18 | 2 | 54.319219 | 25.403611 | U | + | + |
| 18. | Šalčia 2 | SL2 | 12 | 3 | 54.291917 | 25.209867 | N | − | + |
| 19. | Šalčia 3 | SL3 | 7 | 4 | 54.299950 | 25.202411 | N | − | + |
| 20. | Šalčia 4 | SL4 | 3 | 3 | 54.304858 | 25.141147 | N | − | + |
| 21. | Beržė | BR | 41 | 4 | 54.298883 | 25.204058 | N | + | + |
| 22. | Visinčia 1 | VN1 | 24 | 3 | 54.323150 | 25.507589 | A | + | − |
| 23. | Visinčia 2 | VN2 | 12 | 2 | 54.386056 | 25.374642 | A | − | + |
| 24. | Visinčia 3 | VN3 | 6 | 1 | 54.370264 | 25.271892 | N | − | − |
| 25. | Taurupis 1 | TR1 | 16 | 7 | 54.284014 | 24.850161 | A | + | − |
| 26. | Taurupis 2 | TR2 | 20 | 4 | 54.300647 | 24.838781 | A | + | − |
| 27. | Nevėžis 1 | NV1 | 10 | 3 | 55.511533 | 24.768736 | A | + | + |
| 28. | Nevėžis 2 | NV2 | 17 | 8 | 55.533814 | 24.682608 | A | + | + |
| 29. | Nevėžis 3 | NV3 | 24 | 8 | 55.527264 | 24.698569 | A | + | + |
| 30. | Nevėžis 4 | NV4 | 24 | 2 | 55.700283 | 24.433556 | U | − | + |
| 31. | Pienia 1 | PN1 | 24 | 23 | 55.511019 | 24.771975 | A | + | + |
| 32. | Pienia 2 | PN2 | 16 | 1 | 55.434881 | 24.928181 | U | − | + |
| 33. | Širvinta 1 | SR1 | 18 | 1 | 55.062150 | 25.198025 | A | + | + |
| 34. | Širvinta 2 | SR2 | 24 | 3 | 55.028725 | 25.008336 | N | − | + |
| 35. | Siesartis 1 | ST1 | 24 | 8 | 55.227114 | 25.270822 | A | + | − |
| 36. | Siesartis 2 | ST2 | 18 | 4 | 55.291158 | 24.893558 | N | − | + |
| 37. | Siesartis 3 | ST3 | 6 | 3 | 55.226875 | 25.248489 | A | + | − |
| 38. | Šešupė 1 | SP1 | 20 | 11 | 54.356864 | 23.063047 | A | + | + |
| 39 | Šešupė 2 | SP2 | 24 | 18 | 54.409133 | 23.225194 | U | − | + |
| 40. | Šešupė 3 | SP3 | 23 | 17 | 54.417664 | 23.250158 | N | − | + |
| 41. | Kiauna 1 | KN1 | 24 | 24 | 55.306550 | 25.88635 | N | − | + |
| 42. | Kiauna 2 | KN2 | 24 | 24 | 55.294283 | 25.898083 | N | − | + |
| Site | PL | P, % | R | Na | Ne | I | He |
|---|---|---|---|---|---|---|---|
| VR1 | 81 | 44.51 | 0.348 | 1.445 | 1.309 | 0.256 | 0.174 |
| VR2 | 71 | 39.01 | 1.000 | 1.390 | 1.241 | 0.210 | 0.141 |
| VR3 | 95 | 52.20 | 0.444 | 1.522 | 1.331 | 0.284 | 0.191 |
| MR1 | 58 | 31.87 | 1.000 | 1.319 | 1.204 | 0.171 | 0.116 |
| MR2 | 55 | 30.22 | 0.857 | 1.302 | 1.208 | 0.171 | 0.117 |
| MR3 | 31 | 17.03 | 0.429 | 1.170 | 1.109 | 0.092 | 0.062 |
| MR4 | 3 | 1.65 | 0.333 | 1.017 | 1.013 | 0.010 | 0.007 |
| MR5 | 6 | 3.30 | 0.455 | 1.033 | 1.020 | 0.018 | 0.012 |
| MR6 | 4 | 2.20 | 0.400 | 1.022 | 1.018 | 0.014 | 0.010 |
| SK | 0 | 0 | 0.000 | 1 | 1.000 | 0.000 | 0.000 |
| GR1 | 3 | 1.65 | 0.130 | 1.017 | 1.009 | 0.008 | 0.005 |
| GR2 | 67 | 36.81 | 1.000 | 1.368 | 1.262 | 0.215 | 0.148 |
| VS1 | 57 | 31.32 | 0.130 | 1.313 | 1.221 | 0.189 | 0.130 |
| VS2 | 76 | 41.76 | 0.444 | 1.418 | 1.285 | 0.235 | 0.160 |
| SC1 | 48 | 26.37 | 0.222 | 1.263 | 1.182 | 0.153 | 0.104 |
| SC2 | 68 | 37.36 | 0.412 | 1.374 | 1.266 | 0.217 | 0.149 |
| SL1 | 51 | 28.02 | 0.059 | 1.280 | 1.184 | 0.153 | 0.103 |
| SL2 | 6 | 3.30 | 0.182 | 1.033 | 1.017 | 0.017 | 0.011 |
| SL3 | 33 | 18.13 | 0.500 | 1.181 | 1.128 | 0.110 | 0.075 |
| SL4 | 35 | 19.23 | 1.000 | 1.192 | 1.140 | 0.114 | 0.078 |
| BR | 59 | 32.42 | 0.075 | 1.324 | 1.213 | 0.180 | 0.122 |
| VN1 | 50 | 27.47 | 0.087 | 1.275 | 1.198 | 0.162 | 0.111 |
| VN2 | 2 | 1.10 | 0.091 | 1.011 | 1.008 | 0.007 | 0.005 |
| VN3 | 0 | 0 | 0.000 | 0.000 | 1.000 | 0.000 | 0.000 |
| TR1 | 42 | 23.08 | 0.400 | 1.231 | 1.154 | 0.129 | 0.088 |
| TR2 | 38 | 20.88 | 0.053 | 1.209 | 1.148 | 0.126 | 0.086 |
| NV1 | 21 | 11.54 | 0.222 | 1.115 | 1.081 | 0.067 | 0.046 |
| NV2 | 76 | 41.76 | 0.438 | 1.418 | 1.304 | 0.248 | 0.171 |
| NV3 | 61 | 33.52 | 0.304 | 1.335 | 1.225 | 0.193 | 0.131 |
| NV4 | 43 | 23.63 | 0.043 | 1.236 | 1.167 | 0.143 | 0.098 |
| PN1 | 60 | 32.97 | 0.957 | 1.330 | 1.206 | 0.177 | 0.119 |
| PN2 | 0 | 0 | 0.000 | 0.000 | 1.000 | 0.000 | 0.000 |
| SR1 | 0 | 0 | 0.000 | 0.000 | 1.000 | 0.000 | 0.000 |
| SR2 | 40 | 21.98 | 0.087 | 1.220 | 1.155 | 0.128 | 0.088 |
| ST1 | 57 | 31.32 | 0.304 | 1.313 | 1.210 | 0.178 | 0.121 |
| ST2 | 33 | 18.13 | 0.176 | 1.181 | 1.126 | 0.107 | 0.073 |
| ST3 | 25 | 13.74 | 0.200 | 1.137 | 1.105 | 0.083 | 0.058 |
| SP1 | 54 | 29.67 | 0.526 | 1.297 | 1.201 | 0.168 | 0.115 |
| SP2 | 80 | 43.96 | 0.739 | 1.440 | 1.276 | 0.236 | 0.160 |
| SP3 | 80 | 43.96 | 0.727 | 1.440 | 1.291 | 0.243 | 0.165 |
| KN1 | 79 | 43.41 | 1.000 | 1.434 | 1.261 | 0.228 | 0.153 |
| KN2 | 93 | 51.10 | 1.000 | 1.511 | 1.302 | 0.268 | 0.178 |
| Average 1 | 43.83 | 24.09 | 0.399 | 1.169 | 1.161 | 0.136 | 0.092 |
| SE 1 | 4.48 | 2.46 | 0.052 | 0.055 | 0.016 | 0.014 | 0.009 |
| Average 2 | 48.447 | 26.62 | 0.441 | 1.266 | 1.178 | 0.150 | 0.102 |
| SE 2 | 4.300 | 2.36 | 0.053 | 0.024 | 0.015 | 0.013 | 0.009 |
| Population | PL | P, % | Samples | R | Na | Ne | I | He | |
|---|---|---|---|---|---|---|---|---|---|
| Total | G | ||||||||
| VR | 147 | 80.77 | 68 | 38 | 0.552 | 1.808 | 1.494 | 0.430 | 0.288 |
| MR | 132 | 72.53 | 70 | 49 | 0.696 | 1.725 | 1.458 | 0.389 | 0.263 |
| SK | 0 | 0 | 12 | 1 | 0.000 | 0.000 | 1.000 | 0.000 | 0.000 |
| GR | 90 | 49.45 | 38 | 18 | 0.459 | 1.495 | 1.358 | 0.289 | 0.199 |
| VS | 113 | 62.09 | 52 | 17 | 0.314 | 1.621 | 1.409 | 0.340 | 0.231 |
| SC | 110 | 60.44 | 28 | 11 | 0.370 | 1.604 | 1.412 | 0.337 | 0.230 |
| SL | 119 | 65.38 | 40 | 13 | 0.308 | 1.654 | 1.457 | 0.372 | 0.254 |
| BR | 59 | 32.42 | 41 | 4 | 0.075 | 1.324 | 1.214 | 0.180 | 0.122 |
| VN | 94 | 51.65 | 42 | 6 | 0.122 | 1.517 | 1.312 | 0.277 | 0.185 |
| TR | 78 | 42.86 | 36 | 9 | 0.229 | 1.429 | 1.275 | 0.235 | 0.158 |
| NV | 136 | 74.73 | 75 | 22 | 0.284 | 1.747 | 1.500 | 0.419 | 0.285 |
| PN | 83 | 45.60 | 40 | 24 | 0.590 | 1.456 | 1.283 | 0.241 | 0.162 |
| SR | 62 | 34.07 | 42 | 4 | 0.073 | 1.341 | 1.241 | 0.199 | 0.136 |
| ST | 113 | 62.09 | 48 | 14 | 0.277 | 1.621 | 1.420 | 0.349 | 0.238 |
| SP | 129 | 70.88 | 67 | 46 | 0.682 | 1.709 | 1.460 | 0.389 | 0.263 |
| KN | 119 | 65.38 | 48 | 48 | 1.000 | 1.654 | 1.393 | 0.342 | 0.229 |
| Average | 99.00 | 54.40 | 46.69 | 20.25 | 0.377 | 1.482 | 1.355 | 0.299 | 0.203 |
| SE | 9.25 | 5.08 | 4.16 | 4.09 | 0.068 | 0.105 | 0.033 | 0.028 | 0.019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patamsytė, J.; Lambertini, C.; Butkuvienė, J.; Naugžemys, D.; Žvingila, D. Assessment of Anthropogenic Impacts on the Genetic Diversity of Phragmites australis in Small-River Habitats. Diversity 2023, 15, 1116. https://doi.org/10.3390/d15111116
Patamsytė J, Lambertini C, Butkuvienė J, Naugžemys D, Žvingila D. Assessment of Anthropogenic Impacts on the Genetic Diversity of Phragmites australis in Small-River Habitats. Diversity. 2023; 15(11):1116. https://doi.org/10.3390/d15111116
Chicago/Turabian StylePatamsytė, Jolanta, Carla Lambertini, Jurgita Butkuvienė, Donatas Naugžemys, and Donatas Žvingila. 2023. "Assessment of Anthropogenic Impacts on the Genetic Diversity of Phragmites australis in Small-River Habitats" Diversity 15, no. 11: 1116. https://doi.org/10.3390/d15111116
APA StylePatamsytė, J., Lambertini, C., Butkuvienė, J., Naugžemys, D., & Žvingila, D. (2023). Assessment of Anthropogenic Impacts on the Genetic Diversity of Phragmites australis in Small-River Habitats. Diversity, 15(11), 1116. https://doi.org/10.3390/d15111116