Abstract
Charopidae is a highly speciose family of land snails mostly distributed in the main landmasses and archipelagos of the Southern Hemisphere, with a few genera in the Northern Hemisphere. DNA sequencing is making substantial contributions to understanding the phylogeography of Charopidae and the systematic relationships of its subfamilies and related families in the Punctoidea. Here, new data from 28S rRNA are combined with available GenBank sequences to investigate whether Australian Charopidae comprise a single clade, exclusive of species from other regions. Phylogenetic analyses show that this is not the case, with most species from Australia and other regions included in a bootstrap-supported clade that excludes some Australian species. Analyses of concatenated 28S rRNA data and available cytochrome c oxidase subunit I (COI) also contradicted monophyly of Australian species, as did analyses of inferred COI protein sequences. In DNA analyses, the genus Hirasea and Australian taxa with an incomplete secondary ureter were basal within Charopidae. We also examined whether known doubts about the taxonomic status of supra-generic clades in Punctoidea could be resolved by considering autapomorphic COI amino acid changes. Such variation could not generally be applied as an objective criterion for taxonomic categorization but did identify some clades for taxonomic investigation.
1. Introduction
As presently understood, Charopidae F.W. Hutton, 1884 is a highly speciose family of minute to small land snails that is very widely distributed. The family is found in southern Africa [1,2], Australia [3,4,5,6,7,8,9,10,11,12,13], New Zealand [14,15,16] and in many archipelagos of the southern oceans [17,18,19,20,21,22,23,24]. The family is known from Asia [25]. In the Americas, Charopidae is found in South America [26,27,28,29,30,31,32], Central America [33,34] and the U.S.A. [35,36].
Much remains to be learnt of the biodiversity of Charopidae. Many new species continue to be identified and described both from major landmasses such as Africa [37], Australia [38,39,40,41,42,43], continental Asia [44,45] and South America [46,47,48,49,50,51], and from islands including in the Pacific Ocean [52] and the Caribbean Sea [53]. Recent studies of the Australian fauna [39,40,41,42,43,44] provide considerable detail on the utility of protoconch sculpture for defining generic units within the family in the absence of material which is suitable for DNA extraction.
Charopidae belongs to the superfamily Punctoidea which also includes the families Cystopeltidae Cockerell, 1891, Discidae Thiele, 1931 (1866), Endodontidae Pilsbry, 1895, Helicodiscidae Pilsbry, 1927, Oopeltidae Cockerell, 1891, Oreohelicidae Pilsbry, 1939 and Punctidae Morse, 1864. Molecular phylogenetic investigations of Punctoidea [54,55] have revealed that a very substantial amount of work will be required to obtain a robust and resolved phylogeny of the group. Some nominal families have been found not to be supported in these studies. Charopidae, for example, is not monophyletic [54,55]. In particular, clades comprising all Charopidae also include Punctidae. If this is confirmed in further studies the name for the family would be Punctidae, which has priority.
Charopidae is an important component of the land snail fauna of Australia. There are numerous species on the continent, mostly in its east, which is estimated to have more than 700 species, predominantly undescribed ([13], John Stanisic, pers. ob.). Most of the documented species were described on the basis of characteristics of the shell such as size, spire height, ornamentation and umbilicus form. The shells, of which some examples are shown in Figure 1, are small to very small, but often have diagnostic character states such as variation in the height of the spire (Figure 1A,E), the patterns of spiral and radial ribbing on the teleoconch (Figure 1B,F), the patterns (or lack) of spiral and radial ribbing on the protoconch (Figure 1C,G) [56] and the type of umbilicus (Figure 1D,H).

Figure 1.
Scanning electron microscopy images of Charopidae. (A–D) AMS C. 463,085 Elsothera sericatula (Pfeiffer, 1849); (E–H) AMS C.377503 charopid sp. “NN14”. Photographs by Ms Sue Lindsay. Scale bars are 500 µm for all parts except C and G in which they represent 200 µm.
Previous molecular analyses of the phylogeny of Punctoidea have included very few representatives of Australian Charopidae. Consequently, the broader phylogenetic relationships of these taxa are largely unknown. This study was performed to investigate these relationships with the specific aims of resolving (1) whether the Australian species belong to a single clade or to multiple lineages; and (2) whether clades including Australian species are generally basal or derived in the Charopidae and Punctoidea. Such aims could be addressed by combining the published sequences for cytochrome c oxidase subunit I (COI) and the internal transcribed spacer 2 region of the ribosomal genes from Australian species [57] with those of other regions [54,55], but this has not previously been attempted. Data 28S ribosomal RNA sequences are also available from other regions, but these have been reported for only a very few Australian taxa. Additional sequences have been collected from this gene for this study and these are combined with available sequences for analysis. The 28S RNA gene has been widely used in studies of deeper phylogenetic relationships in pulmonate land snails [58]. We selected the D1 expansion of the gene to sequence as this has been found to be sufficiently variable to be informative for gastropods more widely [59,60,61] and because information is also available for this region from other studies of Punctoidea [54,55,58,62].
There are numerous nominal subfamilies of Charopidae [63]. Twelve are currently listed in MolluscaBase [64], and others that are not currently recognized have been proposed [20,54]. At least two subfamilies, Charopinae [20] and Rotadiscinae [20,46] are generally regarded as not monophyletic and this has been confirmed by analysis of DNA sequences [54]. Both of these subfamilies exhibit a considerable range of anatomical diversity. Charopinae, for example, includes a number of Australian taxa such as Setomedea Iredale, 1941, Ngairea Stanisic, 1990 and Hedleyoconcha Pilsbry, 1893 that have an incomplete secondary ureter [9] in contrast to the majority of Charopidae in which this is closed and complete [20]. Molecular data are not available for all subfamilies. However, molecular results [54] indicate that Flammulininae von Martens, 1873 and Ranfurlyinae Schileyko, 2001 are not distinct subfamilies but belong to a strongly supported clade including the type species of Charopinae, Phenacohelicinae and Therasiinae are also unlikely to be distinct [54].
Objective criteria for the definition of taxonomic families and subfamilies are lacking, both in the Punctoidea and more generally. This makes it difficult to develop robust systematics to resolve the nomenclature of Charopidae and its constituent taxa. Diagnostic DNA sequence differences have sometimes been used in the definitions of supra-generic categories [60,65] and programs are available, e.g., [66], to identify diagnostic differences between groups. Less attention has been given to using DNA sequences establish generally applicable criteria for taxonomic definition. Here, we consider whether it might be possible to derive such criteria from protein sequences. We examine whether fixed differences in amino acid sequences in COI, which is quite conservative, can be used to define supra-generic taxa in Punctoidea.
2. Materials and Methods
2.1. 28S rDNA D1 Data Collection
The D1 expansion region of 28S rRNA was amplified by PCR using the D1F [59] and D1R [60] primers. PCR was performed using 0.5 to 1 unit of BIOTAQ DNA polymerase (Bio-Line, London, UK), 0.05 mM dNTPs, 3.5 mM MgCl2, 12.5 pmol of each primer, 1 μL of DNA and purified water to dilute the manufacturer’s buffer to 1X in a total reaction volume of 50 μL.
The cycling profile, following [60] was: 93 °C for 3 min, 50 °C for 1 min, 72 °C for 1 min for one cycle; 93 °C for 30 s, 50 °C for 1 min, 72 °C for 1 min for 32 cycles; and 72 °C for 3 min for the final cycle. Aliquots (3 μL) of PCR products were run on 2% agarose gels and visualized using UV-fluorescence. Single-banded products were treated with ExoSAP-IT (USB Corporation, Cleveland, OH, USA) and sequenced in both directions by Macrogen Inc. (Geumchun-Gu, Korea) using the original primers individually.
National Center for Biotechnology Information (USA) (“NCBI”) GenBank accession numbers for the sequences collected for this study are shown in Table 1.
2.2. Dataset Compilation
NCBI GenBank sequences for Punctoidea (DNA for 28S rRNA and COI, and COI protein sequences) were downloaded. These sequences were reported in [54,55,57,62,67]. COI sequences for most of the specimens for which new 28S rRNA sequences were collected here were available. Alignments were made by CLUSTALX [68] using default parameters.
Specimens lacking data at 30% or more of bases in a sequence were excluded from the DNA datasets. Multiple sequences from the same species were included in the 28S rRNA D1 dataset only where there were differences between them and in the concatenated dataset only where there were differences in at least one of the genes. The 28S rRNA D1 and COI DNA sequences were concatenated for combined analyses using BioEdit [69].
Sequences for the internal transcribed spacer 2 region were also downloaded from NCBI GenBank. However, preliminary analyses of this region showed that there were very few sites remaining in the alignment (<15) after GBlocks analysis [70] (all three relaxed assumptions used) so it was not included in the phylogenetic analyses.
For the 28S rRNA D1 analysis outgroup, we used a range of Panpulmonata species that were sequenced in [71] and did not belong to the stylommatophoran land snails. A similar set of outgroup taxa was used for the combined analysis, although there were some differences as not all outgroup taxa for the 28S rRNA D1 analysis had COI sequences available. Sequences of a number of representative stylommatophoran species available in GenBank were also added as they were used in [54,55] or because of their phylogenetic position as an outgroup with respect to Punctoidea in the Wade et al. [58] topology. These species were Helicodiscus parallelus (Say, 1821) (Helicodiscidae), Allopeas gracile (Hutton, 1834) (Subulinidae), Reticulapex michellae Emberton et al., 2010 and Reticulapex sp. (both Acavidae, Rhytidoidea) and Succinea manaoensis Pilsbry, 1926 (Succinea, Succineoidea) for the 28S rRNA D1 dataset and R. michellae for the combined analysis.

Table 1.
Specimens sequenced, museum registration numbers (those prefaced by “C.” are from the Australian Museum (institutional code: AMS) and those by “MO” from the Queensland Museum (QM), 28S rRNA D1 sequence type and GenBank accession number. Individual specimens from collection lots (with the same registration number) are identified by suffixes beginning with “G”.
Table 1.
Specimens sequenced, museum registration numbers (those prefaced by “C.” are from the Australian Museum (institutional code: AMS) and those by “MO” from the Queensland Museum (QM), 28S rRNA D1 sequence type and GenBank accession number. Individual specimens from collection lots (with the same registration number) are identified by suffixes beginning with “G”.
| Species | Registration Number | 28S rRNA Type | GenBank Accession |
|---|---|---|---|
| Barringtonica montana Shea, Colgan & Stanisic, 2012 | C.472876 | A | OR471375 |
| Barringtonica polblue Shea, Colgan & Stanisic, 2012 | C.309166 | B | OR471374 |
| Cancellocochlea coolongolook Shea, Colgan & Stanisic, 2012 | C.466292 | A | OR471402 |
| Cancellocochlea coolongolook | C.466293 | A | OR471373 |
| Cancellocochlea Shea, Colgan & Stanisic, 2012 cf. coolongolook | MO80164 | A | OR471377 |
| Charopidae NN14 | C.463618 | B | OR471392 |
| Charopidae NN23 | C.463582 | A | OR471384 |
| Comboynea boorganna Shea, Colgan & Stanisic 2012 | C.163287 | A | OR471376 |
| Cumberlandica impressa (Hedley, 1924) | C.457178 | A | OR471381 |
| Cumberlandica impressa | C.462479 | A | OR471379 |
| Cumberlandica impressa | C.462481 | A | OR471372 |
| Cumberlandica impressa | C.462483 | A | OR471380 |
| Cumberlandica impressa | C.463083 | A | OR471389 |
| Cumberlandica impressa | C.463909 | A | OR471383 |
| Decoriropa lirata (Cox, 1864) | C.466867 | C | OR471404 |
| Diphyoropa saturni (Cox, 1864) | C.462488 | A | OR471366 |
| Diphyoropa saturni | C.462489 | A | OR471391 |
| Diphyoropa saturni | C.462495 | A | OR471367 |
| Diphyoropa saturni | C.462525 | A | OR471368 |
| Diphyoropa saturni | C.475921 | A | OR471365 |
| Elsothera sericatula (Pfeiffer, 1849) | C.462523 | A | OR471369 |
| Elsothera sericatula | C.463085 | A | OR471370 |
| Gyrocochlea vinitincta Hedley, 1924 | C.466294 | B | OR471393 |
| Macphersonea canalis (Stanisic, 2010) | C.463585 | D | OR471386 |
| Macrophallikoropa belli (Cox, 1864) | C.465686 | E | OR471403 |
| Macrophallikoropa belli | C.465866 | E | OR471362 |
| Macrophallikoropa Hyman & Stanisic, 2005, sp. | C.466865 | F | OR471401 |
| Nautiliropa omicron (Pfeiifer, 1852) | C.466862_G87 | B | OR471394 |
| Nautiliropa omicron | C.466862_G88 | B | OR471395 |
| Ngairea corticicola (Cox, 1866) | C.466863 | G | OR471397 |
| Planorbacochlea hawkesburyana Stanisic, 2010 | C.462434 | B | OR471382 |
| Planorbacochlea hawkesburyana | C.462496 | B | OR471396 |
| Planorbacochlea hawkesburyana | C.462508 | B | OR471390 |
| Planorbacochlea hawkesburyana | C.462831 | B | OR471405 |
| Planorbacochlea nambucca Shea, Colgan & Stanisic, 2012 | MO44862 | B | OR471378 |
| Planorbacochlea parriwiensis Shea, Colgan & Stanisic, 2012 | C.462478 | A | OR471363 |
| Planorbacochlea parriwiensis | C.462509 | A | OR471364 |
| Planorbacochlea planorbis (Hedley, 1924) | C.309923 | B | OR471371 |
| Planorbacochlea planorbis | C.463729 | B | OR471388 |
| Planorbacochlea Shea, Colgan & Stanisic, 2012 sp. “Port Macquarie” | MO59761 | B | OR471398 |
| Planorbacochlea yessabahensis Shea, Colgan & Stanisic, 2012 | C.168684 | A | OR471387 |
| Planorbacochlea yessabahensis | C.466289 | A | OR471361 |
| Richmondaropa prava (Hedley, 1924) | C.466861 | B | OR471385 |
| Setomedea seticostata (Hedley, 1924) | C.466864_G92 | H | OR471399 |
| Setomedea seticostata | C.466864_G93 | H | OR471400 |
2.3. Data Analysis
2.3.1. Scanning Electron Microscopy
Scanning electron microscopy was performed using an EVO LS15 instrument (Carl Zeiss, Oberkochen, Germany). The specimens were mounted on sticky tabs, gold sputter coated and imaged under high vacuum. The periostracum was not removed from specimens so as to retain as much of the sculpture of the shells as possible.
2.3.2. Genetic Analyses
After final alignment by CLUSTALX [68] and inspection using BioEdit [69], phylogenetic analyses were conducted using maximum likelihood (“ML”) performed on the CIPRES data portal [72] by RAxML [73,74] using the Blackbox and XSEDE interfaces with default assumptions (not using empirical data frequencies, no invariable sites). The 28S rRNA sequences and the three sets of codon positions were treated as separate partitions in the combined analyses. The numbers of required rapid bootstrap replicates were calculated by the majority rules extended (“MRE”) bootstopping criterion [75]. Trees were examined using Figtree v. 1.4.2. (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 31 August 2023) [76]. The MTREV substitution matrix was used for the COI amino acid sequence analysis.
PAUP 4.0a (build 169) [77] was used for listing synapomorphies and determining the consistency index for each character (amino acid site) in the strict consensus of the MP trees of the protein sequence dataset. The MP trees were found using a search with parameters: hsearch, nreps = 100 chucklen = 350, nchuck = 100. The tree was rooted on Discidae. Diagnostic characters were identified as those with ci of 1.00 (i.e., they changed at a single node but were not reversed or paralleled elsewhere in the topology).
MEGA v. 7 [78] was used for determining the number of sites of different types (conserved, parsimony-informative, etc.) in the alignments.
IQ-TREE [79] with 1000 ultrafast bootstrap replicates [80] for ModelFinder [81] and an edge-unlinked partitioned model [82] with 10,000 resampling of estimated log-likelihood replicates [83] was used to compare the unconstrained RAxML tree with the tree constrained not to contradict monophyly of the Australian taxa.
3. Results
The alignments used for the analyses are available in the Supplementary Material. Most sequences of both 28S rDNA D1 and COI were full or nearly full length. The notable exceptions were mostly from the genus Hirasea Pilsbry, 1902. The two 28S rDNA D1 sequences from the genus were among the three shortest sequences for this gene and the four COI sequences from the genus were among the five shortest for this gene.
There were 354 positions in the alignment of the 28S RNA sequences, of which 264 were constant, 24 variable but not parsimony-informative, and 52 parsimony informative.
Eight distinct sequences were found in the newly collected data. Four of these were found only in single specimens but the other four were found in multiple specimens. Two of these occurred in conspecific individuals but the other two (types A and B) were observed in multiple species and genera (Table 1). Types A and B differed by a single base. No species had both types A and B, but the genus Planorbacochlea did have species represented by each type.
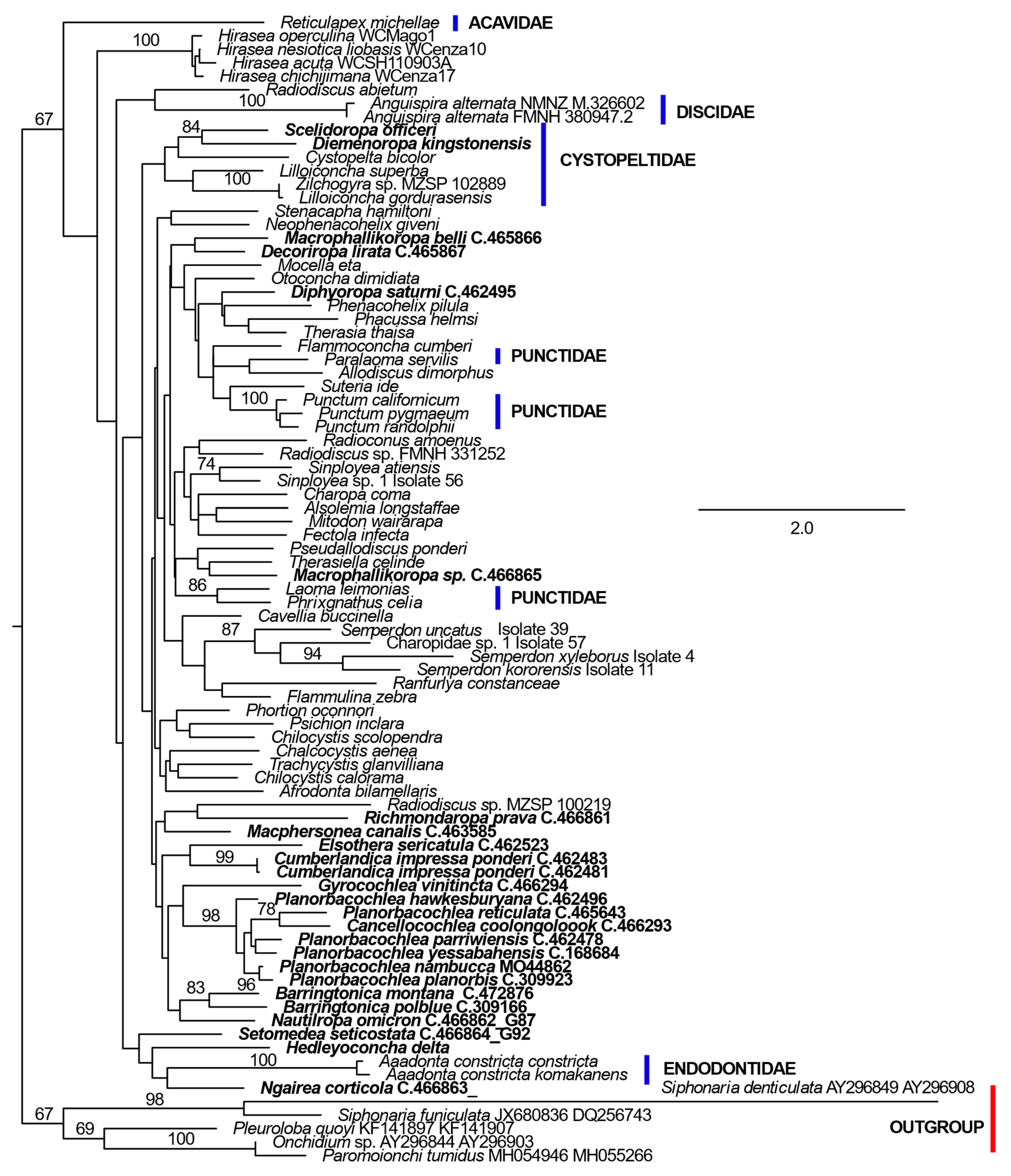
The ln likelihood of the ML tree for the 28S rRNA dataset was −1628.442 (Figure 2). The number of bootstrap replicates determined by the MRE criterion was 750. Excepting the Charopidae and Punctidae, nominal family level taxa were recovered as monophyletic with strong support in this analysis. Charopidae was polyphyletic and Punctidae was separated into two widely divergent clades. One of the punctid clades comprised most members of the family including representatives of the type genus and was monophyletic within a large, bootstrap-supported clade (identified as clade I, Figure 2) also comprising most Charopidae. Clade I also included all Cystopeltidae, which was recovered as a bootstrap-supported derived clade. The second clade of Punctidae which was a lineage in the basal polytomy of Punctoidea was composed of members of the genus Lauria. Gray, 1840 from Europe. The three Australian species with an incomplete secondary ureter [H. delta (Pfeiffer, 1857), N. corticicola and S. seticostata] were included with the Endodontidae in the sister group of Clade I. The remaining nominal Charopidae, the genus Hirasea Pilsbry, 1902, was relatively basal in the topology and resolved, probably as a result of long branch length attraction as the sister group of Succineidae. Most subfamilies in Charopidae (Charopinae, Phenacohelicinae, Rotadiscinae, Trachycystinae) were not monophyletic, in agreement with [54]. The four sequences from Rotadiscinae were widely dispersed in the topology, with none having a member of the subfamily as its sister group. Indeed, one of the sequences, from Radiodiscus abietum (H. B. Baker, 1930) had a species of the family Oreohelicidae as its sister group.
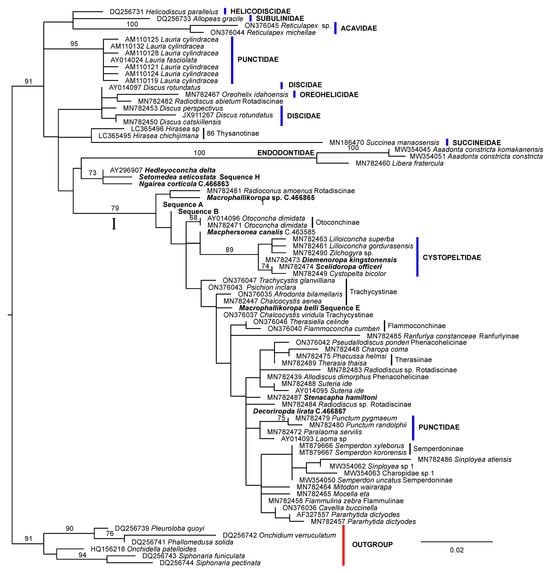
Figure 2.
ML phylogeny of Charopidae and related families based on 28S ribosomal RNA D1 region DNA sequences. Sequences from GenBank are identified by species name and accession number. Multiple sequences are given for species only when they are not identical. Bootstrap percentage support values of 67% or more are shown near branches. The subfamily to which species of Charopidae belong is shown beside the sequence name or thin vertical bars linking multiple taxa. Ingroup sequences lacking family or subfamily names belong to Charopinae. The phylogeny is rooted on the “Outgroup” sequences (defined by the red bar). Australian species are highlighted in bold font. New sequences found in only one individual are identified by Australian Museum registration numbers. Some newly collected sequences were identical in multiple specimens from a species or in multiple species. These are identified by the relevant sequence type detailed in Table 1. The species name is also included where the multiple identical sequences are from conspecific individuals. The scale bar represents the numbers of changes per site.
In the combined analysis, there were 349 positions in the 28S rRNA D1 alignment, of which 243 were constant, 43 variable but not parsimony-informative and 48 parsimony informative. In the COI section of the alignment there were 658 positions of which 281 were constant, 63 variable but not parsimony-informative, and 311 parsimony-informative.
The ln likelihood of the ML tree was −19,949.587 (Figure 3). The number of bootstrap replicates was 100. The ln likelihood of the ML tree constrained not to contradict monophyly of the Australian species was −20,017.180. The number of bootstrap replicates was also 100.
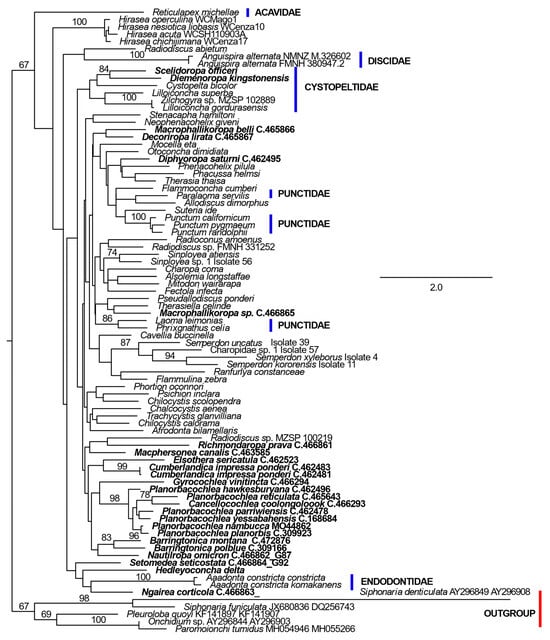
Figure 3.
ML phylogeny of Charopidae and related families based on concatenated COI and 28S ribosomal RNA D1 region DNA sequences. Sequences were obtained from GenBank accessions specified in the articles [54,55,62,65]. Where only one specimen of a species is treated in the original article, these are identified in the topology by species name. For species with multiple representatives in one of the articles, additional identifying information is given. Specimens with new 28S rDNA sequences are identified by Australian Museum registration numbers. Bootstrap percentage support values of 67% or more are shown near branches. The phylogeny is rooted on the “Outgroup” sequences (defined by the red bar). Australian species are highlighted in bold font. The scale bar represents the numbers of changes per site.
All comparisons in the IQ-TREE output gave very similar results, in which the unconstrained tree was significantly better than the tree constrained not to contradict monophyly of the Australian taxa. The constrained tree was excluded from the 95% confidence set in all comparisons. In particular, for the approximately unbiased test of Shimodaira [84], the probability of the unconstrained tree was 0.983 and that of the constrained tree was 0.017.
In the combined ML analysis, the ingroup had bootstrap support of 67%, but the deeper nodes within this had low support. Notably, in contrast to the 28 rRNA D1 analysis (Figure 2), Cystopeltidae was not significantly supported. Punctidae was polyphyletic in this analysis and Endodontidae was embedded in the group of Australian species with an incomplete secondary ureter (H. delta, N. corticicola and S. seticostata). This relative lack of resolution compared to the 28S rRNA D1 analysis suggests that the addition of COI data tended to obscure relationships within Punctoidea, possibly as a result of homoplasious, synonymous changes at first and third codon positions in this coding sequence.
Albeit lacking statistical support, Acavidae was recovered as the sister group of the Punctoidea, with Hirasea forming one branch of the basal dichotomy in this superfamily. The next most basally divergent clades were firstly, the Discidae plus Radiodiscus abietum and then the Endodontidae plus Australian species with an incomplete secondary ureter. Most of the other Australian species were recovered in a single clade including Richmondaropa prava, Macphersonea canalis, Elsothera sericatula, Cumberlandica Shea, Colgan & Stanisic, 2012, Planorbacochlea Shea, Colgan & Stanisic, 2012, Cancellocochlea Shea, Colgan & Stanisic, 2012, Barringtonica Shea, Colgan & Stanisic, 2012 and Nautiliropa omicron. A few species, from the genera Macrophallikoropa Hyman & Stanisic, 2005 and Decoriropa Hyman & Stanisic, 2005, were found in other clades. Notably, M. belli was the sister group of D. lirata, rather than being embedded in the Trachycystinae as in the 28S DNA D1 analysis (Figure 2).
In contrast to supra-generic taxa, most genera with multiple representatives received strong bootstrap support in the combined ML analysis. The exceptions were separation of the two species of Macrophallikoropa and the inclusion of Cancellocochlea coolongolook within Planorbacochlea with strong bootstrap support.
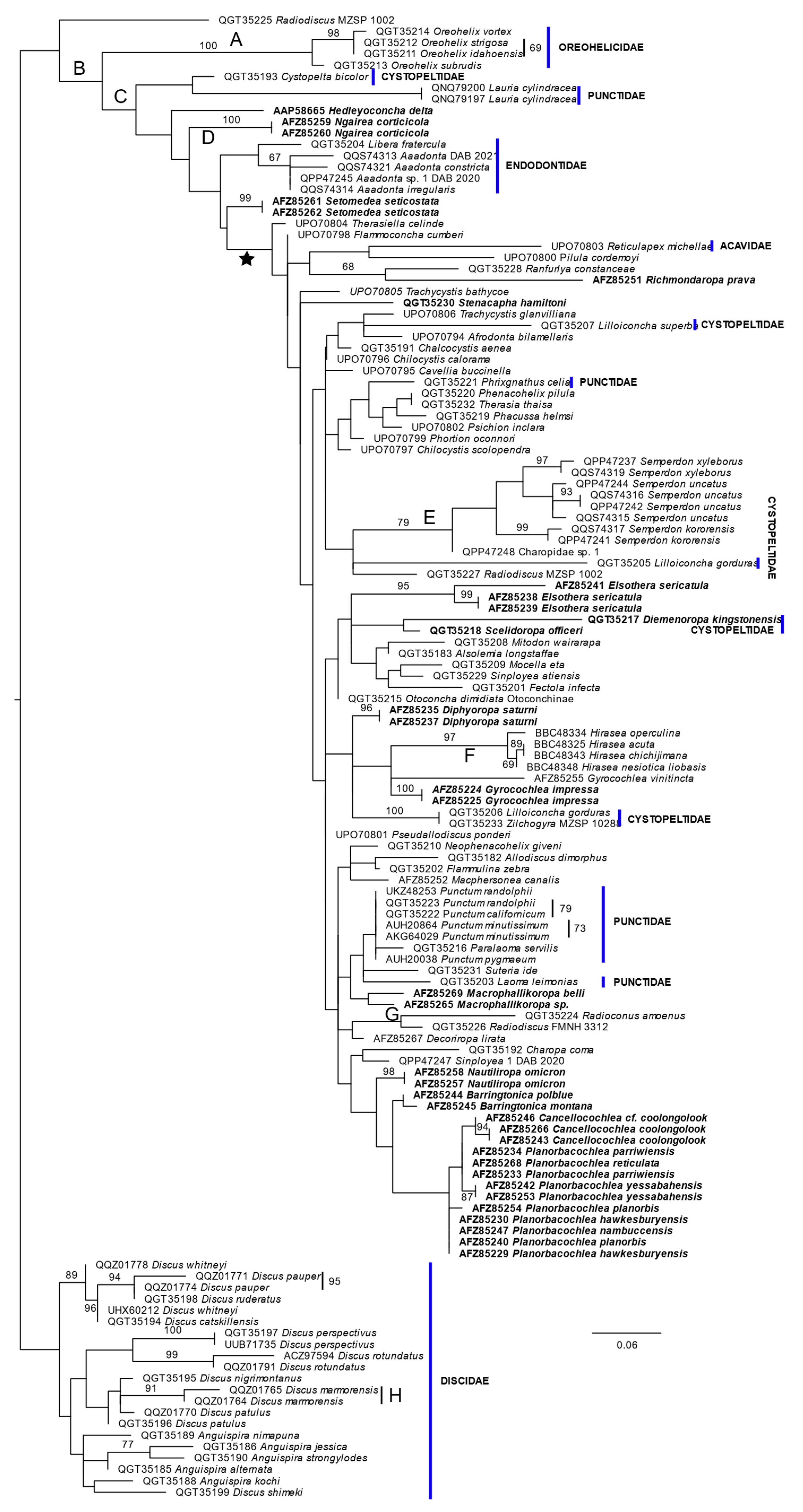
There were 219 positions in the alignment of the COI amino acid sequences, of which 138 were constant, 20 variable but not parsimony-informative and 61 parsimony-informative. One hundred shortest trees, of length 448, were found in the maximum parsimony analysis. The ln likelihood of the ML tree was −3163.058 (Figure 4). The number of bootstrap replicates determined by the MRE criterion was 350.
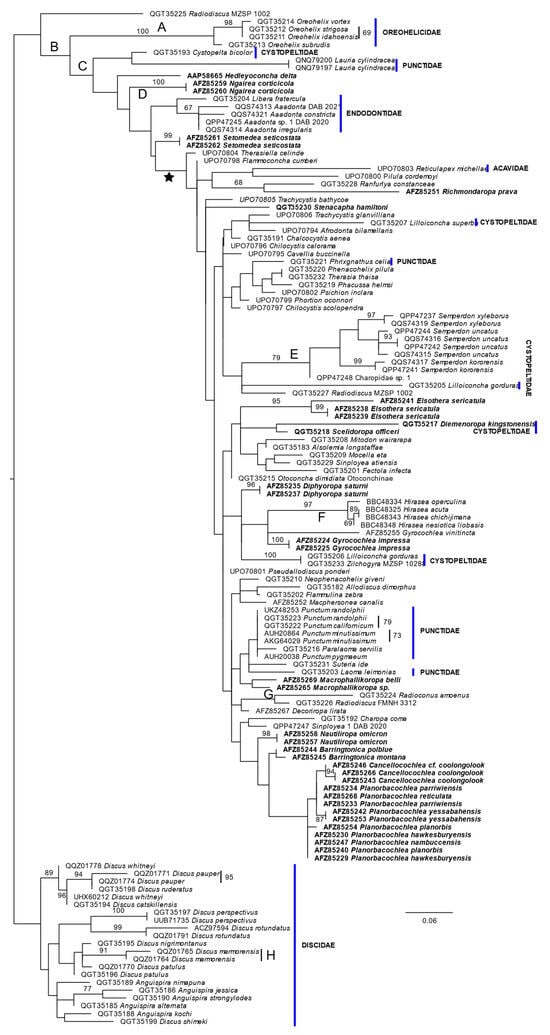
Figure 4.
ML phylogeny of COI amino acid sequences. Sequences are identified by species name and GenBank accession number. The scale bar represents the numbers of changes per site. Bootstrap percentage support values of 67% or more are shown near branches or beside bars to the right of the included sequences. The phylogeny is rooted on the Discidae. The star defines a clade discussed in the text. Letters along branches specify changes that do not occur elsewhere in the MP tree. These changes are from the first amino acid to the second in the following list (using the one letter code). The number indicates the position of the amino acid in the alignment. (A) 7, G-A; (B) 144, S-A; (C) 91, A-S; (D) 171, I-L; (E) 23, L-M; (F) 203, F-I; (G) 4, M-V; and (H) 135, L-M.
When rooted on Discidae, all other taxa in the phylogeny except for a species of Radiodiscus Pilsbry, 1906, were characterized by a single non-reversed amino acid change (B). This large group of taxa is divided into two clades, Oreohelidicae and all others, both of which have a synapomorphic non-reversed change. One branch in the non-Oreohelicidae clade contains a cystopeltid and the two representatives of the punctid genus Lauria. The other branch has a series of bifurcations within the Australian taxa with an incomplete secondary ureter and the Endodontidae. Most Australian taxa were found in the large clade formed by the last of these bifurcations (identified by the star in Figure 4).
In this analysis, Hirasea was recovered in a relatively derived position compared to the DNA analyses, albeit that the genus was characterized by a synapomorphic amino acid change (F; Figure 4). The nominal families Cystopeltidae and Punctidae were each separated into multiple clades that were widely dispersed in the topology.
The Rotadiscinae were found in two clades in this analysis. One, as noted above, was basal in the ingroup. The other, comprising Radioconus amoenus (Thiele, 1927) and Radiodiscus abietum was resolved in a derived position, but defined by a synapomorphic change (G, Figure 4).
4. Discussion
The Australian species of Charopidae were not resolved as a monophyletic clade in any of the analyses, whether of the 28S rRNA D1 dataset, the concatenated COI and 28S rRNA D1 dataset or the inferred COI amino acid sequences dataset. Significantly longer trees are required to recover the Australian species as a monophyletic clade. In all of the analyses, the three sequenced genera with an incomplete secondary ureter (Hedleyoconcha, Setomedea and Ngairea) were relatively basal within Charopidae in the recovered topologies in comparison to all taxa except (in the DNA analyses; Figure 2 and Figure 3) the genus Hirasea from Japan and the Ogasawara Islands. This genus was long considered to belong to Endodontidae. However, Hirasea (s.s.) diplomphalus Pilsbry, 1902 has a complete secondary ureter, unlike Endodontidae [20] and was referred to Helicarionidae by [85]. The genus is presently treated as a member of Charopidae in MolluscaBase [86] but the rationale for this is not provided. The genus cannot be definitely assigned to a family on the basis of the present analyses, a situation that is possibly complicated by the relatively short sequences that were available for inclusion here.
Understanding of the evolution of Punctoidea is certainly in its early stages. Consequently, we do not suggest any nomenclatural changes in this manuscript. However, the results do highlight the necessity for these to be made in the future and they do have implications for the changes that might need to occur. In considering such changes, we give less weight to the concatenated analysis because of its relative lack of resolution compared to the 28S rRNA D1 analysis.
Charopidae is not monophyletic within Punctoidea, reinforcing doubts raised in previous DNA analyses [54,55] as to whether it should be recognized as a family with its presently understood membership. One possibility would be to synonymize Charopidae with Punctidae as that family is presently understood. We prefer not to do this, as the present analyses would also require Endodontidae and Cystopeltidae to be included in the enlarged Punctidae, making it unwieldy.
If Charopidae is not synonymized with Punctidae, the question arises as to whether its name should be restricted to a monophyletic subset of the taxa it presently encompasses, with the excluded taxa being referred to new families or to previously proposed groups not now recognized such as Hedleyoconchidae of which H. delta is the type species [87].
An alternative approach, based on the 28S rRNA D1 analysis and supported by the COI amino acid phylogeny (Figure 4), would be to restrict the current definition of Punctidae (because of the polyphyly caused by the placement of Lauria) to its members belonging to Clade I of the 28S rRNA D1 phylogeny whilst also extending it to comprise the other taxa included in the clade. Punctidae would then include Charopidae (other than Hirasea and the Australian taxa with incomplete secondary ureters) and Cystopeltidae.
The basal position of Hirasea and the Australian taxa with incomplete secondary ureters suggests, in combination with the large geographical distance between their distributions, that considerable investigation would be required to elucidate the phylogeographic history of Charopidae. Even if Hirasea is found not to belong to Charopidae, that history is likely to be complex, as the fossil record suggests that the distribution of the family in the past included regions where it is not now found. Most fossil records are from the Southern Hemisphere. These records are mostly from South America [88,89,90,91,92,93,94,95,96], where the family is first recorded from the Eocene [88,89,90] and has also been found in Oligocene, Miocene and Pliocene deposits. Charopidae is also known from the Miocene and later of New Zealand [97] and from the Miocene in islands in the Pacific Ocean [98]. There is also a fossil record from the Eocene in Germany [99], suggesting that the family had a wider distribution in the past and complicating discussion of its geographic origin.
The range of the Rotadiscinae is particularly problematic as it putatively extends from North and South America to the Fiji archipelago in the Pacific Ocean and possibly Western Australia [20], although the resemblance between material from this state to members of the subfamily is most likely due to morphological convergence (John Stanisic, pers. ob.). The subfamily was not monophyletic in previous molecular studies [54] or in any of the present analyses. It had representatives that were quite basal in the analyses of the concatenated DNA data (Figure 3) and the amino acid sequences (Figure 4), although this representative differed in the two cases. Further investigation is required to establish how many subfamilies the taxon comprises and to determine their phylogeographic relationships.
As noted in the Introduction, there are no available objective criteria for answering questions about which clades should be recognized by naming them as supra-generic taxa. Here, we have investigated the possible use of diagnostic amino acid changes in the definition of such taxa. The approach has some potential but the case of COI in Punctoidea also shows that it has limitations that must be considered in any application.
If possible, it would be best to have criteria for defining supra-generic taxa that were both necessary and sufficient. In the present example, there were unexpectedly high numbers of substitutions between inferred COI amino acid sequences. This suggests that changes in these sequences are not per se sufficient to define families and subfamilies. Conversely, this level of variability suggests that distinct taxa should be diagnosable by differences in the COI sequences, i.e., that the criterion could be considered necessary. COI substitutions that are potentially diagnostic synapomorphies were found here. Some supported taxa that are already recognized (e.g., Oreohelicidae, Semperdoninae) and others suggested that taxonomic recognition at the subfamily, or possibly even family, level might be warranted for some groups that are not currently recognized. These include the synapomorphy of the genus Hirasea which is currently placed in Thysanotinae in MolluscaBase [86] but without a statement of rationale. No sequences of other Thysanotinae are available in GenBank.
There are a number of factors to consider in the potential use of amino acid changes in COI (or other proteins) to define subfamilies of families. One consideration is the set of taxa to be included in the definition. Searches for diagnostic DNA differences frequently emphasise pairwise differences between groups [66]. These are not necessarily synapomorphic for either group when more inclusive taxa are considered. In searching for applicable criteria based on amino acid sequences, we may be more interested in changes that are synapomorphic on a broader level. The effect of taxon inclusion is illustrated by the example of Acavidae in the present analyses. If the sequences from this family were excluded, Cystopeltidae, Endodontidae, Charopidae and Punctidae would be grouped by a diagnostic amino acid difference from Discidae and Oreohelicidae (and one Radiodiscus species). However, blastp searches of Panpulmonata suggest that the difference is a synapomorphy of Discidae and Oreohelicidae and the Radiodiscus species rather than the former group. A second consideration is that taxa could be defined by having non-overlapping sets of amino acids at the same site, rather than having only a single form in all of their included species (“asymmetric characters” sensu [100]. Thirdly, sequencing errors could potentially obscure diagnostic differences between taxa so these should be considered as a possibility when all but a very few species in a group have an amino acid that differs from another group.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15111124/s1; the 28S rDNA D1 alignments, the concatenated DNA alignment and the COI amino acid alignment. All files are in fasta format.
Author Contributions
Both authors are responsible for planning, fieldwork for specimen collection and manuscript writing and preparation. DNA sequencing and analysis were conducted by Colgan. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by an ABRS research (grant 206–39 to D. Colgan and J. Stanisic).
Data Availability Statement
All sequences, including those newly collected here are available in GenBank. The sequence alignment matrices are available as supplementary material.
Acknowledgments
We thank Sue Lindsay for specimen preparation and SEM photography, and Alison Miller, Janet Waterhouse, Amanda Reid (all of the Malacology Section, Australian Museum) and Darryl Potter (Malacology Section, Queensland Museum) for specimen curation. The authors would like to thank Lorelle Stanisic (Honorary Researcher, Queensland Museum) for comments on the manuscript. We thank the NSW National Parks and Wildlife Service to permission to collect in nature reserves and national parks under its authority. We thank the three anonymous reviewers for their very helpful comments on the original version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Emberton, K.C.; Pearce, T.A.; Kasigwa, P.F.; Tattersfield, P.; Habibu, Z. High diversity and regional endemism in land snails of eastern Tanzania. Biodivers. Conserv. 1997, 6, 1123–2236. [Google Scholar] [CrossRef]
- Herbert, D.; Kilburn, D. Field Guide to the Land Snails and Slugs of Eastern South Africa; Natal Museum: Pietermaritzburg, South Africa, 2004. [Google Scholar]
- Hedley, C. Observations on the Charopidae. Proc. Linn. Soc. N.S.W. 1892, 7, 157–169. [Google Scholar]
- Hedley, C. Some notes on Australian land shells. Aust. Zool. 1924, 3, 215–221. [Google Scholar]
- Kershaw, R.C. Studies on Australian Charopidae. Part 2. Some genera. Vic. Nat. 1955, 72, 28–30. [Google Scholar]
- Kershaw, R.C. Studies on Australian Charopidae. Part 3. Planate genera. Vic. Nat. 1956, 72, 137–143. [Google Scholar]
- Stanisic, J. Studies on the Charopidae of tropical and subtropical Australia. 1. Oreokera: A primitive genus from the high mountains of North Queensland (Mollusca: Pulmonata: Charopidae). J. Malacol. Soc. Aust. 1987, 8, 1–21. [Google Scholar]
- Stanisic, J. Systematics and biogeography of eastern Australian Charopidae (Mollusca, Pulmonata) from subtropical rainforests. Mem. Queensl. Mus. 1990, 30, 1–241. [Google Scholar]
- Smith, B.J.; Kershaw, R.C. Description of three Tasmanian charopid genera (Pulmonata: Mollusca) with notes on their type species. Rec. Queen Vic. Mus. Art Gall. 1985, 88, 1–14. [Google Scholar]
- Stanisic, J. Danielleilona gen. nov., from the Wet Tropics, northeastern Queensland (Pulmonata: Charopidae). Mem. Queensl. Mus. 1993, 34, 11–20. [Google Scholar]
- Stanisic, J. Lenwebbia paluma sp. nov., from the Wet Tropics, northeastern Queensland (Pulmonata: Charopidae). Mem. Queensl. Mus. 1993, 34, 21–26. [Google Scholar]
- Hyman, I.T.; Stanisic, J. New charopid landsnails, chiefly from limestone outcrops in eastern New South Wales (Eupulmonata: Charopidae). Mem. Queensl. Mus. 2005, 50, 219–302. [Google Scholar]
- Stanisic, J.; Shea, M.; Potter, D.; Griffiths, O. Australian Land Snails Volume 1: A Field Guide to Eastern Australian Species; Bioculture Press: Senneville, Mauritius, 2010. [Google Scholar]
- Climo, F.M. Classification of New Zealand Arionacea. Nat. Mus. N. Z. Rec. 1981, 2, 9–15. [Google Scholar]
- Barker, G.M. The character of the New Zealand land snail fauna and communities: Some evolutionary and ecological perspectives. Rec. W. A. Mus. 2005, 68, 53–102. [Google Scholar] [CrossRef]
- Marshall, B.A.; Barker, G.M. A revision of the New Zealand landsnails referred to Allodiscus Pilsbry, 1892 and Pseudallodiscus Climo, 1971, with the introduction of three new genera (Mollusca: Gastropoda: Charopidae). Tuhinga 2008, 19, 57–167. [Google Scholar]
- Muratov, I.V.; Abdou, A.; Bouchet, P. Charopid land snails (Gastropoda Pulmonata Charopidae) from Mayotte, Comores: Alive and well. Trop. Zool. 2005, 18, 171–208. [Google Scholar] [CrossRef]
- Iredale, T. The land Mollusca of Lord Howe Island. Aust. Zool. 1944, 10, 299–334. [Google Scholar]
- Iredale, T. The land Mollusca of Norfolk Island. Aust. Zool. 1945, 11, 46–71. [Google Scholar]
- Solem, A. Endodontoid Land Snails from Pacific Islands (Mollusca: Pulmonata: Sigmurethra). Part II. Families Punctidae and Charopidae, Zoogeography; Field Museum: Chicago, IL, USA, 1983. [Google Scholar]
- Pawlowska-Banasiak, E. Andrefrancia Solem, 1960 (Gastropoda: Pulmonata: Charopidae)—A systematic revision. Folia Malacol. 2008, 16, 101–195. [Google Scholar] [CrossRef]
- Slapcinsky, J. A new species of Paryphantopsis (Gastropoda: Pulmonata: Charopidae) from Crater Mountain, Simbu (Chimbu) Province, Papua New Guinea. Nautilus 2009, 123, 53–58. [Google Scholar]
- Brook, F.J. Coastal landsnail fauna of Rarotonga, Cook Islands: Systematics, diversity, biogeography, faunal history, and environmental influences. Tuhinga 2010, 21, 161–252. [Google Scholar]
- Pugh, P.J.A.; Lewis Smith, R.I. Notodiscus (Charopidae) on South Georgia: Some implications of shell size, shell shape, and site isolation in a singular sub-Antarctic land snail. Antarct. Sci. 2011, 23, 442–448. [Google Scholar] [CrossRef]
- Schileyko, A.A. A redescription of Ruthvenia biciliata (L. Pfeiffer, 1855), with revised generic diagnosis for Ruthvenia Gude, 1911 (Gastropoda: Pulmonata: Charopidae). Ann. Nat. Mus. Wien Ser. B 2010, 111, 13–18. [Google Scholar]
- Hausdorf, B. The genus Lillioconcha in Colombia (Gastropoda: Charopidae). J. Nat. Hist. 2005, 39, 2795–2808. [Google Scholar] [CrossRef]
- Cadiz, F.J.; Gallardo, C.S. Morphological and anatomical features of Flammulina festiva Scott, 1970 (Stylommatophora; Charopidae) from southern Chile, with notes on its natural history. Gayana 2008, 72, 1–8. [Google Scholar]
- Miquel, S.E.; Cadiz Lorca, F.J. Araucocharopa gallardoi gen. et sp. n. de Charopidae (Mollusca: Gastropoda: Stylommatophora) del sur de Chile. Rev. Mus. Argent. Cienc. Nat. 2008, 10, 329–339. [Google Scholar] [CrossRef]
- Delhey, V.K.; Piza, J.; Burela, S. Zilchogyra franzi Weyrauch, 1965 (Gastropoda: Charopidae), a minute landsnail rediscovered in Sierra de la Ventana (Southern Pampas, Argentina). Zootaxa 2010, 2450, 61–64. [Google Scholar] [CrossRef]
- Agudo-Padron, A.I. Threatened freshwater and terrestrial molluscs (Mollusca, Gastropoda et Bivalvia) of Santa Catarina State, Southern Brazil: Check list and evaluation of regional threats. Biodivers. J. 2011, 2, 59–66. [Google Scholar]
- Cunha, C.M.; Salvador, R.B.; Simone, L.R.L. The terrestrial microgastropods of Trindade Island, Brazil. Spixiana 2015, 38, 139–143. [Google Scholar]
- Pérez, A.M.; López, A. Listado de la malacofauna continental (Mollusca: Gastropoda) del Pacífico de Nicaragua. Rev. Biol. Trop. 2003, 51 (Suppl. S3), 405–451. [Google Scholar]
- Correa Sandoval, A. Zoogeografía de los gastrópodos terrestres de la región oriental de San Luis Potosí, México. Rev. Biol. Trop. 1999, 47, 493–502. [Google Scholar] [CrossRef]
- Ramirez Perez, M.C.; Hausdorf, B. Low abundance but high land snail diversity in montane rainforest on the western slope of the Andes in Ecuador. J. Mollusc. Stud. 2022, 88, eyab048. [Google Scholar] [CrossRef]
- Branson, B.A. Radiodiscus hubrichti (Pulmonata: Endodontidae) new species from the Olympic Peninsula, Washington. Nautilus 1975, 89, 47–48. [Google Scholar]
- Nekola, J.C. Overview of the North American terrestrial gastropod fauna. Am. Malacol. Bull. 2014, 32, 225–235. [Google Scholar] [CrossRef]
- Herbert, D.G. Revision of the aperturally dentate Charopidae (Gastropoda: Stylommatophora) of southern Africa—Genus Afrodonta s. lat., with description of five new genera, twelve new species and one new subspecies. Eur. J. Taxon. 2020, 629, 1–55. [Google Scholar] [CrossRef]
- Holcroft, L. A revision of Gyrocochlea-grade Charopidae from mid-eastern Queensland and redescription and generic reassignment of three Gyrocochlea-grade species (Eupulmonata: Charopidae). Mem. Queensl. Mus. Nat. 2018, 61, 1–28. [Google Scholar] [CrossRef]
- Holcroft, L. A revision of Charopidae with a finely cancellate protoconch sculpture from mid-eastern Queensland (Eupulmonata: Charopidae). Mem. Queensl. Mus. Nat. 2018, 61, 83–107. [Google Scholar] [CrossRef]
- Holcroft, L.; Stanisic, J. Thirteen new charopid land snails from mid-eastern Queensland rainforests (Gastropoda: Eupulmonata: Charopidae). Mem. Queensl. Mus. Nat. 2018, 61, 155–186. [Google Scholar] [CrossRef]
- Stanisic, J. Two new species of pinwheel snail from contrasting Queensland habitats and a redescription and generic re-assignment of Gyrocochlea myora Stanisic, 2010 (Gastropoda: Eupulmonata: Charopidae). Mem. Queensl. Mus. 2016, 60, 1–12. [Google Scholar] [CrossRef]
- Stanisic, J. New species and genera of pinwheel snails from the brigalow lands of south central Queensland (Gastropoda: Eupulmonata: Charopidae). Mem. Queensl. Mus. Nat. 2020, 62, 157–185. [Google Scholar]
- Stanisic, J. A new genus and three new species of pinwheel snails from Queensland and New South Wales (Gastropoda: Eupulmonata: Charopidae). Mem. Queensl. Mus. Nat. 2022, 64, 1–11. [Google Scholar] [CrossRef]
- Vermeulen, J.J.; Marzuki, M.E. ‘Charopa’ lafargei (Gastropoda, Pulmonata, Charopidae), a new, presumed narrowly endemic species from Peninsular Malaysia. Basteria 2014, 78, 31–34. [Google Scholar]
- Sutcharit, C.; Pholyotha, A. Hidden lineages in the mountains: The genus Glyptaulax Gude, 1914 and Maelamaodiscus gen. nov. (Heterobranchia: Stylommatophora: Charopidae and Ariophantidae) with description of two new species from western Thailand. Trop. Nat. Hist. 2023, 7, 123–138. [Google Scholar]
- Miquel, S.E.; Barker, G.M. New Charopidae from Chilean—Argentine Patagonia (Mollusca: Gastropoda: Stylommatophora). Arch. Mollusk. 2009, 138, 53–61. [Google Scholar] [CrossRef]
- Miquel, S.E.; Araya, J.F. A new Charopidae from Chile and Argentina, Stephacharopa calderaensis n. gen. and n. sp., with remarks on the taxonomy of the genus Stephadiscus Hylton Scott 1981 (Mollusca: Gastropoda Pulmonata). Arch. Molluskenk. 2013, 142, 227–235. [Google Scholar] [CrossRef]
- Araya, J.F.; Miquel, S.E. A new Stephacharopa (Gastropoda: Punctoidea: Charopidae) From Paposo, Northern Chile. J. Conch. 2018, 43, 129–133. [Google Scholar]
- Miquel, S.E.; Araya, J.F. Re-description of genitalia of Stephacharopa and description of a new Peruvian genus (Gastropoda, Stylommatophora, Charopidae). Spixiana 2022, 45, 1–7. [Google Scholar]
- Salvador, R.B.; Cavallari, D.C.; Simone, L.R.L. Taxonomical study on a sample of land snails from Alto Ribeira State Park (Sao Paulo, Brazil), with description of a new species. Arch. Molluskenk. 2016, 145, 59–68. [Google Scholar] [CrossRef]
- Salvador, R.B.; Charles, L.; Simone, L.R.L.; Maestrati, P. Terrestrial gastropods from Pedra Talhada Biological Reserve, Alagoas state, Brazil, with the description of a new species of Radiodiscus (Gastropoda: Charopidae). Arch. Molluskenk. 2018, 147, 101–128. [Google Scholar] [CrossRef]
- Miquel, S.E.; Araya, J.F. New records of terrestrial molluscs of the Juan Fernandez Archipelago (Chile), with the description of a new genus and species of Charopidae (Gastropoda: Stylommatophora). Arch. Molluskenk. 2015, 144, 155–167. [Google Scholar] [CrossRef]
- Rutherford, M.G. A new species of Radiodiscus (Gastropoda: Eupulmonata: Charopidae) from Trinidad and Tobago. Arch. Molluskenk. 2020, 149, 67–74. [Google Scholar] [CrossRef]
- Salvador, R.B.; Brook, F.J.; Shepherd, L.D.; Kennedy, M. Molecular phylogenetic analysis of Punctoidea (Gastropoda, Stylommatophora). Zoosyst. Evol. 2020, 96, 397–410. [Google Scholar] [CrossRef]
- Salvador, R.B. Phylogenetic Position of African punctoid snails (Stylommatophora, Punctoidea, Trachycystinae). Taxonomy 2022, 2, 227–235. [Google Scholar] [CrossRef]
- Holcroft, L. Protoconch sculpture as a taxonomic tool in Australian charopid systematics (Gastropoda: Eupulmonata: Charopidae). Mollusc. Res. 2018, 38, 258–273. [Google Scholar] [CrossRef]
- Shea, M.; Colgan, D.J.; Stanisic, J. Systematics of the landsnail genus Gyrocochlea and relatives (Mollusca: Charopidae). Zootaxa 2012, 3585, 1–109. [Google Scholar] [CrossRef]
- Wade, C.M.; Mordan, P.B.; Naggs, F. Evolutionary relationships among the Pulmonate land snails and slugs (Pulmonata, Stylommatophora). Biol. J. Linn. Soc. 2006, 87, 593–610. [Google Scholar] [CrossRef]
- McArthur, A.G.; Koop, B.F. Partial 28S rDNA sequences and the antiquity of the hydrothermal vent endemic gastropods. Mol. Phylogenet. Evol. 1999, 13, 255–274. [Google Scholar] [CrossRef]
- Colgan, D.J.; Ponder, W.F.; Beacham, E.; Macaranas, J. Molecular phylogenetics of Caenogastropoda (gastropoda: Mollusca). Mol. Phylogenet. Evol. 2007, 42, 717–737. [Google Scholar] [CrossRef]
- Colgan, D.J.; Ponder, W.F.; Beacham, E.; Macaranas, J.M. Molecular phylogenetic studies of Gastropoda based on six gene segments representing coding or non-coding and mitochondrial or nuclear DNA. Mollusc. Res. 2003, 23, 123–148. [Google Scholar] [CrossRef]
- Bullis, D.A.; Rundell, R.J. Molecular phylogenetics and premating isolation in the punctoid land snails of Belau (Republic of Palau, Oceania). Zool. Script. 2021, 50, 555–570. [Google Scholar] [CrossRef]
- Schileyko, A.A. Treatise on recent terrestrial pulmonate molluscs. Part 7. Endodontidae, Thyrophorellidae, Charopidae. Ruthenica 2001, 2, 881–1034. [Google Scholar]
- MolluscaBase. Charopidae F. W. Hutton, 1884. 2023. Available online: https://www.molluscabase.org/aphia.php?p=taxdetails&id=816165 (accessed on 4 September 2023).
- Baker, R.J.; Solari, S.; Cirranello, A.; Simmons, N.B. Higher level classification of phyllostomid bats with a summary of DNA synapomorphies. Acta Chiropt. 2016, 18, 1–38. [Google Scholar] [CrossRef]
- Hütter, T.; Ganser, M.H.; Kocher, M.; Halkic, M.; Agatha, S.; Augsten, N. DeSignate: Detecting signature characters in gene sequence alignments for taxon diagnoses. BMC Bioinform. 2020, 21, 151. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Wada, S.; Mori, H.; Uchida, S.; Saito, T.; Chiba, S. Genetic and morphometric rediscovery of an extinct land snail on oceanic islands. J. Mollusc. Stud. 2018, 84, 148–156. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL X Windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl. Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Holznagel, W.E.; Colgan, D.J.; Lydeard, C. Pulmonate phylogeny based on 28S rRNA gene sequences: A framework for discussing habitat transitions and character transformation. Mol. Phylogenet. Evol. 2010, 57, 1017–1025. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), 14 Nov (2010); Gateway Computing Environments Workshop: New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 312–313. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web-servers. Syst. Biol. 2008, 75, 758–771. [Google Scholar] [CrossRef]
- Pattengale, N.D.; Alipour, M.; Bininda-Emonds, O.R.P.; Moret, B.M.E.; Stamatakis, A. How many bootstrap replicates are necessary? J. Computat. Biol. 2010, 17, 337–354. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Tracer 1.3. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 31 August 2023).
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony; Version 4.0; Laboratory of Molecular Systematics, Smithsonian Institution: Washington, DC, USA, 2003. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [PubMed]
- Bui, Q.M.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar]
- Diep, T.H.; Chernomor, O.; von Haeseler, A.; Bui, Q.M.; Le, S.V. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar]
- Kalyaanamoorthy, S.; Bui, Q.M.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Meth. 2017, 14, 587–589. [Google Scholar]
- Chernomor, O.; von Haeseler, A.; Bui, Q.M. Terrace aware data structure for phylogenomic inference fromsupermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar]
- Kishino, H.; Hasegawa, M. Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data, and the branching order in hominoidea. J. Mol. Evol. 1989, 29, 170–179. [Google Scholar]
- Shimodaira, H. An approximately unbiased test of phylogenetic tree selection. Syst. Biol. 2002, 51, 492–508. [Google Scholar] [CrossRef]
- Ueshima, R.; Kurozumi, T. Anatomical features of Hirasea (ss) diplomphalus and taxonomic position of the genus Hirasea Pilsbry, 1902 (Pulmonata: Sigmurethra). Venus 1988, 47, 261–270. [Google Scholar]
- MolluscaBase. Hirasea Pilsbry, 1902. 2021. Available online: https://molluscabase.org/aphia.php?p=taxdetails&id=996012 (accessed on 4 September 2023).
- Iredale, T. Guide to the land shells of New South Wales, part IV. Aust. Nat. 1942, 11, 33–40. [Google Scholar]
- Bellosi, E.S.; Miquel, S.E.; Kay, R.F.; Madden, R.H. Un calcrete de edad Mustersense de Patagonia central con los primeros Charopidae (Gastropoda) del Eoceno: Significado paleoclimático. Ameghiniana 2002, 39, 465–477. [Google Scholar]
- Miquel, S.E.; Bellosi, E.S. Un nuevo microgastrópodo terrestre (Charopidae) del Eoceno de Patagonia central, Argentina. Ameghiniana 2004, 41, 111–114. [Google Scholar]
- Miquel, S.E.; Bellosi, E.S. Microgasterópodos terrestres (Charopidae) del Eoceno medio de Gran Barranca (Patagonia Central, Argentina). Ameghiniana 2007, 44, 121–131. [Google Scholar]
- Miquel, S.E.; Ramirez, R. First records of actual and fossil Stephadiscus outside Patagonia, and description of a new Amazonian species (Mollusca: Pulmonata: Charopidae). Arch. Molluskenk. 2011, 140, 49–56. [Google Scholar]
- Rodriguez, P.E.; Miquel, S.E.; Tauber, A.A.; Krapovickas, J.M. First record of land gastropods of the family Charopidae in the Early to Middle Miocene from Santa Cruz Province, Southern Patagonia, Argentina (Gastropoda Pulmonata: Stylommatophora: Charopidae). Arch. Molluskenk. 2012, 141, 57–66. [Google Scholar] [CrossRef]
- Salvador, R.B.; Simone, L.R.L. Taxonomic revision of the fossil pulmonate mollusks of Itaborai Basin (Paleocene), Brazil. Pap. Avul. Zool. 2013, 53, 5–46. [Google Scholar] [CrossRef]
- Miquel, S.E.; Rodriguez, P.E. A novel late Early Miocene assemblage of terrestrial gastropods from Santa Cruz (Patagonia, Argentina). J. Paleont. 2015, 89, 748–761. [Google Scholar]
- Turazzini, G.F.; Miquel, S.E. A terrestrial gastropod community from the early Pliocene (Neogene) of Mendoza, Argentina, with description of a new species of Radiodiscus Pilsbry and Ferriss, 1906 (Mollusca: Pulmonata: Charopidae). Ameghiniana 2014, 51, 396–404. [Google Scholar]
- Salvador, R.B.; Cabrera, F.; Martínez, S.; Miquel, S.E.; Simone, L.R.L.; Cunha, C.M. Annotated catalogue of the fossil Hygrophila and Eupulmonata (Mollusca: Gastropoda) from South America (Cretaceous–Neogene). Jb. Geol. Paläont. Abh. 2018, 289, 249–280. [Google Scholar] [CrossRef]
- Marshall, B.A.; Worthy, T.H. Miocene land snails (Mollusca: Gastropoda: Pulmonata) from palaeolake Manuherikia, southern New Zealand. J. Roy. Soc. N. Z. 2017, 47, 294–318. [Google Scholar]
- Ladd, H.S. Fossil land shells from western Pacific atolls. J. Paleontol. 1958, 32, 183–198. [Google Scholar]
- Kadolsky, D. A remarkable non-marine mollusc fauna of early Eocene age from a fissure infill in Karsdorf quarry (Sachsen-Anhalt, Germany). Geol. Saxonica 2020, 65–66, 31–76. [Google Scholar]
- Wägele, J.-W.; Rödding, F. A priori estimation of phylogenetic information conserved in aligned sequences. Mol. Phylogenet Evol. 1998, 9, 358–365. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).