
Genotypic and Phenotypic Diversity of Endemic Golden Camellias Collected from China

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Part
2.1.1. Test Materials
2.1.2. DNA Extraction, Enzyme Digestion and Library Construction for Sequencing
2.1.3. SNP Calling
2.1.4. Genetics Analysis
2.2. Chemical Profiling
2.2.1. Total Phenolic Content
2.2.2. Total Flavonoid Content
2.2.3. Total Anthocyanin Content
2.2.4. Ellagic Acid Content
2.2.5. Data Analysis
3. Results
3.1. SNP Calling Based on the Reference Genome
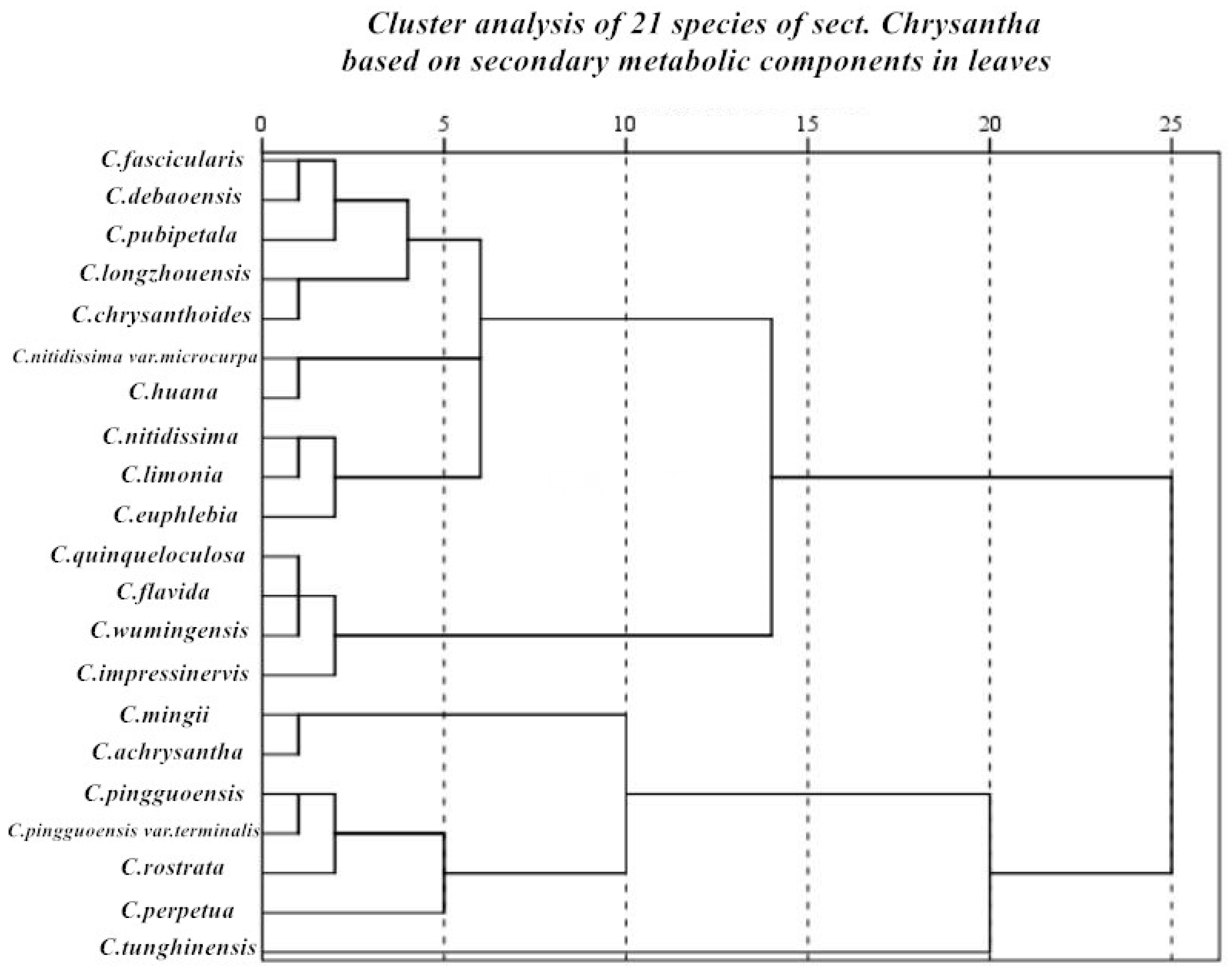
3.2. Genetic Relationship and Phylogenetic Analysis
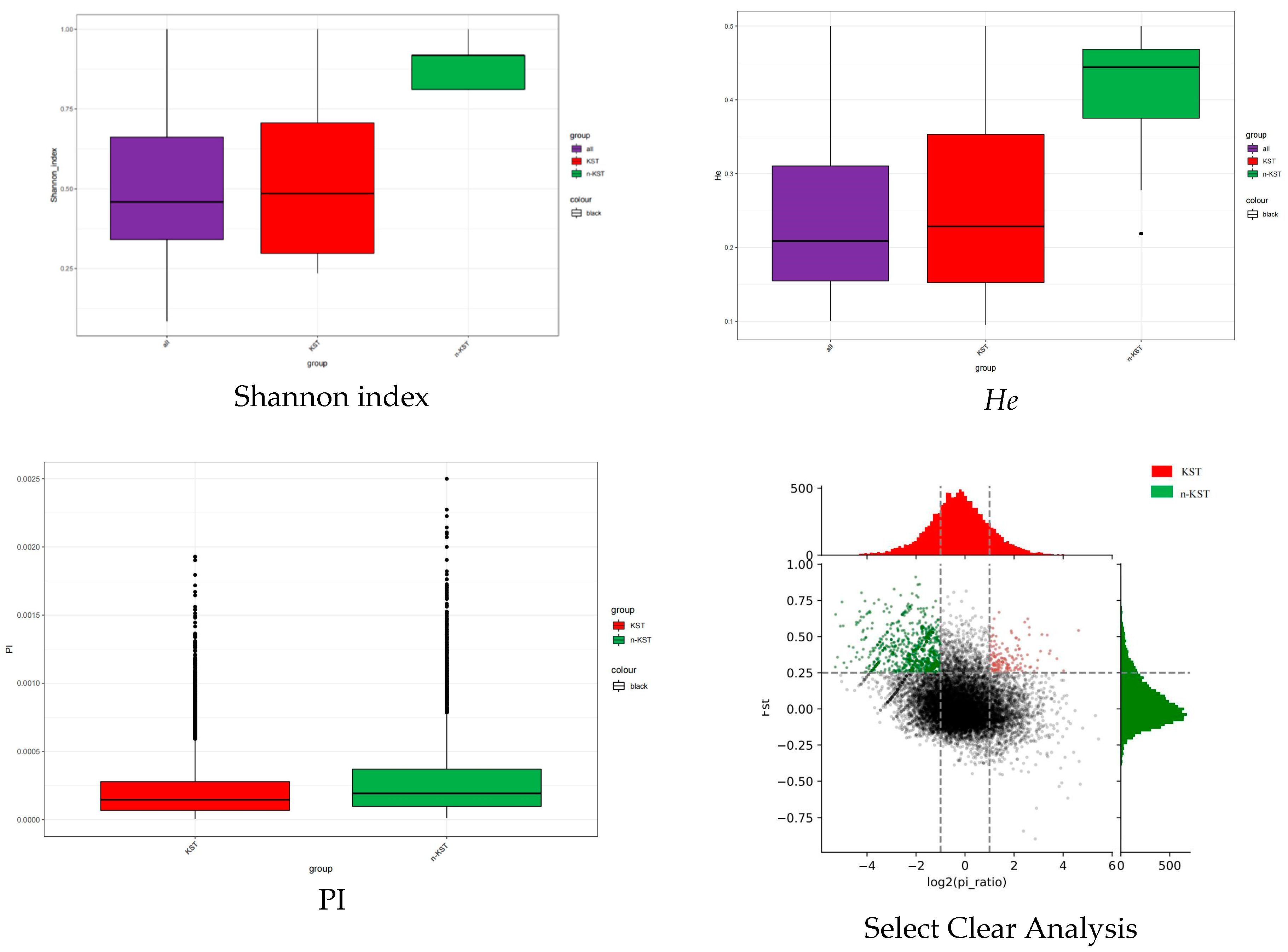
3.3. Comparison of the Genetic Diversity between Karst and Non-Karst
3.4. Chemical Activity and Cluster Analysis of Plant Leaves of Camellia Scet. Chrysantha
3.4.1. Chemical Profile Content Comparative Analysis
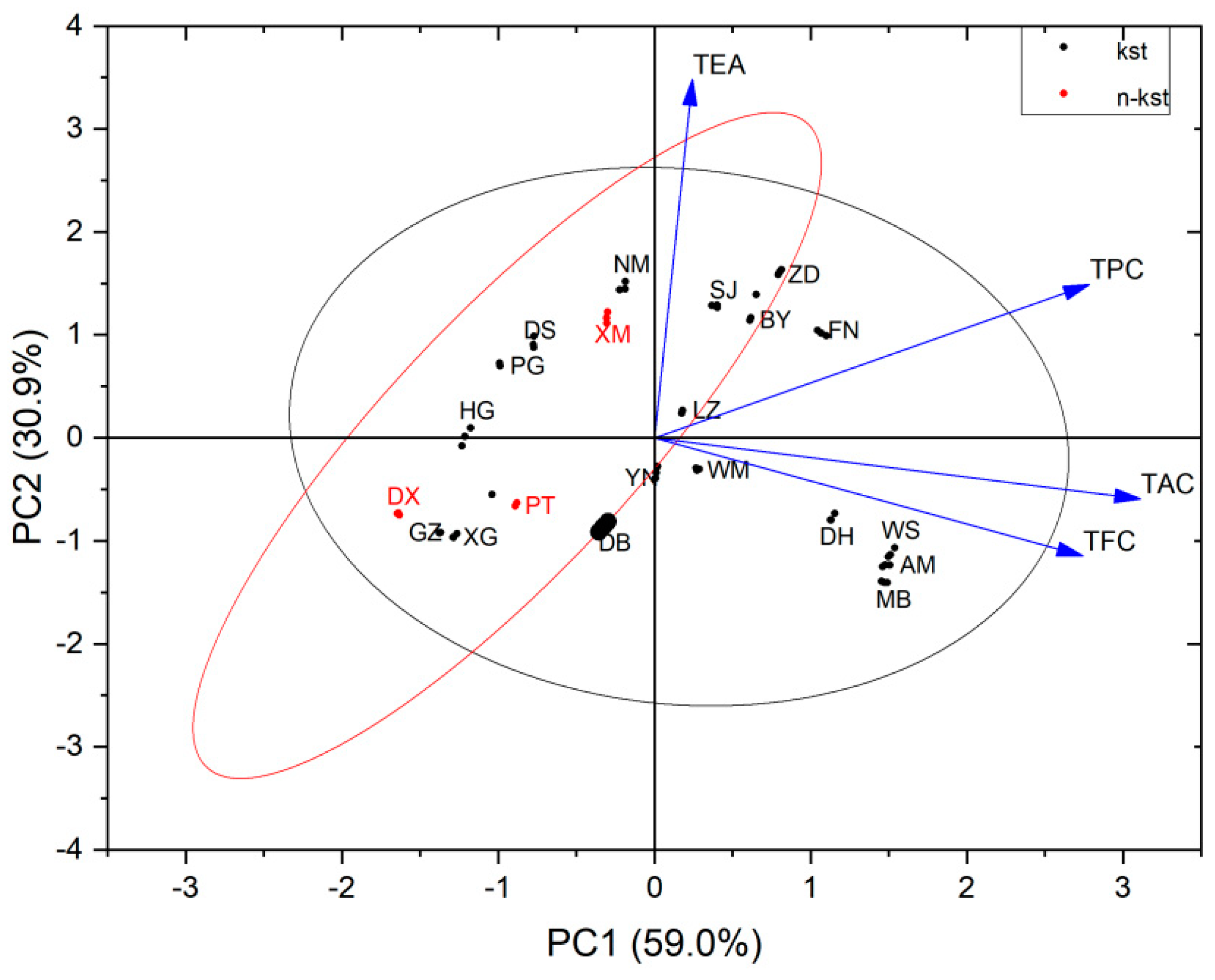
3.4.2. Principal Component Cluster Analysis
4. Discussion
4.1. Relationship between Plant Species in Camellia Sect. Chrysantha
4.2. Effects of the Different Environmental Pressures on the Evolutionary Selection of Camellia Sect. Chrysantha
4.3. Principal Component Cluster Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, X.; Jiang, S.Y.; Jiang, Y.S.; Tang, H.; Cao, H.L. Research progress of rare and endangered plant Camellia nitidissima. Fujian For. Sci. Technol. 2006, 33, 169–174. [Google Scholar]
- Gao, H. Construction of Golden Camellia Fingerprint Map and Screening of Excellent Germplasm. Ph.D. Thesis, Minzu University of China, Beijing, China, 2023. [Google Scholar]
- Shi, Y.T. Research on the Conservation Genetics of Camellia debaoensis and Camellia mingii Based on Simplified Genome Sequencing Technology. Master’s Thesis, Guangxi Normal University, Guilin, China, 2023. [Google Scholar]
- Huang, X.N.; Qi, Y.; Ye, P.M.; Jiang, C.J.; Li, G.E.; Li, Z.H. Research on the Breeding of Two New Golden Camellia Hybrid Varieties. Anhui Agric. Sci. 2021, 16, 54–56+66. [Google Scholar]
- Huang, L.D. Registration of new camellia varieties (Part 1). Chin. Flower Gard. 2010, 16, 44–47. [Google Scholar]
- Hakoda, N. Pedigree and breeding status of yellow camellia. Rep. Hortic. Cult. Inst. Huiquan Women’s Coll. Hortic. Cult. 2006, 3, 43–69. [Google Scholar]
- Nishikawa, H.; Tanabe, R.; Takahashi, K. Deep mountain landscape and golden tea growing environment in Kye Hung County, North Central Vietnam. Landsc. Tech. Rep. Collect. 2023, 2, 42–43. [Google Scholar]
- Tang, J.M.; Shi, Y.C.; Liao, Y.Q.; Fan, J.S.; Wei, J.Q.; Wen, X.Y.; Wei, X. Analysis of nutritional components of Camellia nitidissima. Guihaia 2017, 37, 1176–1181. [Google Scholar]
- Chen, Y.X.; Lv, S.J.; Wei, J.B. Research progress on the chemical composition and pharmacological effects of Camellia nitidissima. Guangxi Tradit. Chin. Med. 2013, 1, 4–6. [Google Scholar]
- Niu, G.J.; Xin, J.H.; Zhu, S.; Wang, W.H.; Chen, Q.Y.; Liang, Y.C. Determination of active ingredients and antioxidant activity of Camellia nitidissima. J. For. Environ. 2015, 35, 165–168. [Google Scholar]
- Cao, F.; Fan, L.L. Research progress of Golden Camellia. China Pharm. Ind. 2013, 22, 95–96. [Google Scholar]
- Deng, G.Y.; Yang, Z.D.; Lu, T.L. Overview of research on yellow Camellia in China. Guangxi Agric. Biosci. 2000, 19, 126–130. [Google Scholar]
- lden Flower Tea series of tea snacks sell well at home and abroad. Mod. Mark. (Bus. Ed.) 2015, 8, 26.
- Zhu, X.L.; Tang, J.M.; Qin, H.Z.; Bai, K.D.; Chen, Z.Y.; Zou, R.; Chai, S.F. Contrasting Adaptation Mechanisms of Golden Camellia Species to Different Soil Habitats Revealed by Nutrient Characteristics. Agronomy 2022, 12, 1511. [Google Scholar] [CrossRef]
- Ye, C.X.; Xu, Z.R. Research on golden camellia Formation. J. Sun Yat-Sen Univ. Nat. Sci. Ed. 1992, 31, 68–77. [Google Scholar]
- Tang, S.Q.; Shi, S.H.; Chen, Y.Q.; Qu, L.H.; Zhang, H.D. RAPD analysis and taxonomic significance of Golden Camellia and related species. J. Sun Yat-Sen Univ. (Nat. Sci. Ed.) 1998, 4, 29–33. [Google Scholar]
- Tang, S.Q.; Du, L.F.; Wang, Y. AFLP analysis of the golden camellia Camellia genus. Wuhan Bot. Res. 2004, 22, 44–48. [Google Scholar]
- Xiao, Z.; Li, J.Y.; Li, Z.H.; Huang, L.D.; Fan, A.Q.; Jiang, C.J. Using ISSR analysis of genetic relationships among Camellia species. For. Sci. Res. 2014, 1, 71–76. [Google Scholar]
- Chen, H.L. Study on the Genetic Diversity and Genetic Structure of Camellia chrysanthoides and Its Related Species. Master’s Thesis, Guangxi Normal University, Guilin, China, 2019. [Google Scholar]
- Lu, X.L.; Chen, H.L.; Wei, S.J.; Tang, S.Q. Chloroplast and nuclear DNA analyses provide insight into the phylogeography and conservation genetics of Camellia nitidissima (Theaceae) in southern Guangxi, China. Tree Genet. Genomes 2022, 16, 8. [Google Scholar] [CrossRef]
- Wei, S.J.; Liufu, Y.Q.; Zheng, H.W.; Chen, H.L.; Lai, Y.C.; Liu, Y.; Tang, S.Q. Using phylogenomics to untangle the taxonomic incongruence of yellow-flowered Camellia species (Theaceae) in China. J. Syst. Evol. 2022, 61, 748–763. [Google Scholar] [CrossRef]
- Zhang, Y.Z. Distribution Patterns of Secondary Metabolites in the Tree of Life of Seed Plants. Ph.D. Thesis, Yunnan University, Kunming, China, 2020. [Google Scholar]
- Le, V.A.; Mai Nga, T.X.; Nhi Nguyen, T.; Nguyen, K.O. Genotypic and Phenotypic Diversity of Endemic Golden Camellias Collected from the North of Vietnam. Chem. Biodivers. 2023, 20, e202200843. [Google Scholar]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- Zhu, X.L.; Zou, R.; Qin, H.Z.; Chai, S.F.; Tang, J.M.; Li, Y.Y.; Wei, X. Genome-wide diversity evaluation and core germplasm extraction in ex situ conservation: A case of golden Camellia tunghinensis. Evol. Appl. 2023, 16, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.M.; Sul, J.H.; Service, S.K.; Zaitlen, N.A.; Kong, S.Y.; Freimer, N.B.; Sabatti, C.; Eskin, E. Variance component model to account for sample structure in genome-wide association studies. Nat. Genet. 2010, 42, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Zhang, W.P.; Zhang, Z.Y.; Wang, Y.P.; Jiang, W. Study on extraction process of polyphenols in Camellia nitidissima. Chin. Cosmet. 2003, 2, 114–120. [Google Scholar]
- Piyawan, P.; Pensri, T.; Chutharat, T.; Kaewta, D.; Víctor, C. Development of a Digital Microscope Spectrophotometric System for Determination of the Antioxidant Activity and Total Phenolic Content in Teas. Anal. Lett. 2021, 54, 2727–2735. [Google Scholar]
- Zhang, P.; Yu, B.; Zou, C.P.; Huang, L.; Zhao, C.Y.; Sun, Y.B. Research progress on biological functions and extraction and separation methods of flavonoids in Camellia sinensis. Guangdong Agric. Sci. 2019, 46, 13–20. [Google Scholar]
- Wang, L.L.; Lin, Q.X.; Song, Z.S.; Chen, L. Spectrophotometric determination of total flavonoid content in tea. J. Tea 2021, 25, 740–750. [Google Scholar]
- Wu, Y.Y.; Hou, D.Y.; Hui, R.H. Determination of proanthocyanidin content in Tie Guanyin tea and tea stem. J. Anshan Norm. Univ. 2012, 2, 30–33. [Google Scholar]
- Ortega Vidal, J.; Ruiz Martos, L.S.S.; Altarejos, J. Proantho cyanidins in Pruning Wood Extracts of Four European Plum (Prunus domestica L.) Cultivars and Their hLDHA Inhibitory Activity. Chem. Biodivers. 2023, 20, e202200931. [Google Scholar] [CrossRef]
- Deng, W.J. Study on the conversion law of free ellagic acid and the determination method of raspberry content. Pharm. Today 2023, 6, 431–434. [Google Scholar]
- Wang, J.L.; Zhao, L.Y.; Wu, W.L.; Li, W.L.; Zhang, C.H. Change law of ellagic acid in blackberry fruit and its correlation with quality index. South. Chin. Fruit Trees 2023, 3, 154–159. [Google Scholar]
- Wang, A.; Huang, X.L.; Liang, X.J.; Wei, X.J.; Li, K.X.; Ma, J.L. Comparison of leaf active ingredient content of 11 species of Camellia sinensis group. Econ. For. Res. 2018, 1, 110–114. [Google Scholar]
- Li, X.M.; Li, C.N.; Lu, J.S.; Huang, Z.W.; Cui, X.Q.; Zhang, Z.B.; Bu, C.Y. Analysis and evaluation of flower active components of six yellow Camellia group plants. Food Res. Dev. 2020, 21, 33–37. [Google Scholar]
- Zhu, C.H.; Tang, J.M.; Jiang, H.L.; Zou, R.; Qin, H.Z.; Chai, S.F.; Wei, X. Evaluation of nutritional value of Guangxi Yuliang Tiankeng characteristic plant stem Huashan pomelo. J. Guangxi Acad. Sci. 2020, 1, 109–116. [Google Scholar]
- Hardy, O.J.; Vekemans, X. SPAGeDi: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Molecular ecology notes. Mol. Ecol. Notes 2002, 2, 618–620. [Google Scholar] [CrossRef]
- Wei, X.; Haung, X.X.; Jiang, Y.S.; Tang, H.; Qi, X.X.; Chen, Z.Y. Comparison of the antioxidant activity of three plant extracts from the Golden Camellia group. Chin. J. Tradit. Chin. Med. 2011, 36, 639–641. [Google Scholar]
- Jiang, L.N.; Li, J.Y.; Fan, Z.Q.; Tong, R.; Mo, R.H.; Li, Z.; Jiang, C.J. Analysis of polyphenol components in flowers of Golden Camellia plants. For. Sci. Res. 2020, 33, 117–126. [Google Scholar]
- Zhang, H.L.; Yu, Q.T.; Wu, Q.X.; Qin, X.M. Research on online screening of antioxidant substances and in vivo antioxidant activity of Yellow Camellia flavonoids based on the action of iron ions combined with high-performance liquid chromatography. Nat. Prod. Res. Dev. 2022, 32, 719–726. [Google Scholar] [CrossRef]
- Yan, D.M.; Yao, A.D. Research on the antioxidant properties of Camellia nitidissima polyphenols. J. Henan Univ. Technol. Nat. Sci. Ed. 2009, 30, 42–45. [Google Scholar]
- Duan, X.X.; Tang, X.L.; Su, J.J.; Wang, Y.; Li, Y.; Yang, C.; Fu, J.Y. Study on the inhibitory effect of Camellia nitidissima on liver cancer induced by diethylnitrosamine in rats. J. Med. Res. 2006, 35, 14–16. [Google Scholar]
- Nong CLChen, Y.X.; He, X.K.; Wei, J.B. Experimental study on the in vitro anti-tumor activity of total flavonoids of Camellia nitidissima. Chin. J. Cancer Prev. Treat. 2013, 4, 324–327. [Google Scholar]
- Wang, Z.L.; Guo, Y.J.; Zhu, Y.Y.; Chen, L.; Wu, T.; Liu, D.B.; Du, H.Z. Study on the mechanism of the effective part of Camellia nitidissima inhibiting epidermal growth factor receptor (EGFR) against non-small cell lung cancer. China J. Tradit. Chin. Med. 2021, 46, 5362–5371. [Google Scholar]
- Tian, X.C.; Qin, X.M.; Lin, H.J.; Yang, J.Z.; Wei, L. Study on the physical and chemical properties of Camellia nitidissima polysaccharides. J. Chin. Food Sci. 2011, 8, 47–52. [Google Scholar]
- Wei, L.; Qin, X.M.; Lin, H.J.; Ning, E.C.; Yang, H. Study on the hypolipidemic function of Camellia nitidissima polysaccharide. Food Sci. Technol. 2008, 33, 247–249. [Google Scholar]
- Zhang, H.L. Effect and Mechanism of Golden Camellia Flavonoids on Food-Borne Obesity. Ph.D. Thesis, Guangdong Ocean University, Zhanjiang, China, 2020. [Google Scholar]
- Tsoi, B.; Gao, C.; Yan, S.Y.; Du, Q.H.; Yu, H.; Li, P.; Shen, J.G. Camellia nitidissima Chi extract promotes adult hippocampal neurogenesis and attenuates chronic corticosterone-induced depressive behaviours through regulating Akt/GSK3β/CREB signaling pathway. J. Funct. Foods 2022, 95, 105119. [Google Scholar] [CrossRef]
- An, L.; Zhang, W.; Ma, G.W.; Wang, Y.S. Neuroprotective effects of Camellia nitidissima Chi leaf extract in hydrogen peroxide-treated human neuroblastoma cells and its molecule mechanisms. Food Sci. Nutr. 2022, 8, 4782–4793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.W.; Zhao, L.; Tan, W. Advances in the application of plant proanthocyanidins. Chin. Fruits Veg. 2023, 7, 24–28+41. [Google Scholar]
- Liu, H.; Sun, J.; Fei, X.W.; Yu, H.L.; Cui, Y.F.; Song, L.L. Research progress on physiological efficacy and analysis of proanthocyanidins. Food Res. Dev. 2023, 12, 211–217. [Google Scholar]
- Zeng, W.; Gu, Z.L. Research progress on the pharmacological effects of ellagic acid. J. Chang. Inst. Technol. 2023, 2, 72–77. [Google Scholar]
- Yu, W.D.; Liu, M.; Song, P. Research progress on biological activity and analysis of ellagic acid. Bioprocessing 2023, 21, 83–90. [Google Scholar]
- Xia, M.L.; Yin, Y.X.; Xiao, Y.T.; Zheng, S.Z.; Tan, B.; Wang, L.; Chen, J.S. Research progress on the physiological functions of ellagic acid and its application in animal production. Feed. Res. 2023, 11, 149–154. [Google Scholar]
- Zhang, Q.; Folk, R.A.; Mo, Z.Q.; Ye, H.; Zhang, Z.Y.; Peng, H.; Yu, X.Q. Phylotranscriptomic analyses reveal deep gene tree discordance in Camellia (Theaceae). Mol. Phylogenetics Evol. 2023, 188, 107912. [Google Scholar] [CrossRef] [PubMed]
- Endashaw, T.; Gurja, B.; Abdulfatai, T.; Jianlin, H.; Olivier, H. Whole Genome Resequencing Reveals Genetic Diversity and Selection Signatures of Ethiopian Indigenous Cattle Adapted to Local Environments. Diversity 2023, 15, 540. [Google Scholar]
- Li, J.B.; Hashimoto, F.; Shimizu, K. Chemical taxonomy of red flowered wild Camellia species based on floral anthocyanins. Phytochemistry 2013, 85, 99–106. [Google Scholar] [CrossRef]
- Li, J.-H.; Nesumi, A.; Shimizu, K.; Sakata, Y.; Liang, M.-Z.; He, Q.-Y.; Zhou, H.-J.; Hashimoto, F. Chemosystematics of tea trees based on tea leaf polyphenols as phenetic markers. Phytochemistry 2010, 71, 1342–1349. [Google Scholar] [CrossRef]
- Min, T.L.; Zhang, W.J. Taxonomic issues of the Ancient Tea Group and golden camellia Group. Yunnan Bot. Res. 1993, 1, 1–15. [Google Scholar]
- Wu, Q. Study on the Phylogeny of Camellia and the Variation of Its Characteristic Secondary Metabolites. Ph.D. Thesis, Anhui Agricultural University, Hefei, China, 2023. [Google Scholar]
- Zhong, C.S.; Lin, Y.L.; Liu, S.C.; Chen, Y.; Lai, Z.X. Research progress on secondary metabolites of Camellia nitidissima. Hortic. Seedl. 2014, 7, 58–62. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nm. | Scientific Name | Code | Place of Origin | Habitat In Situ |
|---|---|---|---|---|
| 1 | Camellia achrysantha | ZD | Guangxi | Karst |
| 2 | Camellia chrysanthoides | BY | Guangxi | Karst |
| 3 | Camellia debaoensis | DB | Guangxi | Karst |
| 4 | Camellia euphlebia | XM | Guangxi | Non-Karst |
| 5 | Camellia fascicularis | YN | Yunnan | Karst |
| 6 | Camellia flavida | DH | Guangxi | Karst |
| 7 | Camellia huana | GZ | Guangxi/Guizhou | Karst |
| 8 | Camellia impressinervis | AM | Guangxi | Karst |
| 9 | Camellia libelofifilamenta | LR | Guangxi/Guizhou | Karst |
| 10 | Camellia limonia | NM | Guangxi | Karst |
| 11 | Camellia longgangensis | NG | Guangxi | Karst |
| 12 | Camellia longzhouensis | LZ | Guangxi | Karst |
| 13 | Camellia micrantha | XH | Guangxi | Karst |
| 14 | Camellia mingii | FN | Yunnan/Guangxi | Karst |
| 15 | Camellia multipetala | DD | Guangxi | Karst |
| 16 | Camellia multipetala var. patens | ZM | Guangxi | Karst |
| 17 | Camellia nitidissima | PT | Guangxi | Non-Karst |
| 18 | Camellia nitidissima var. microcurpa | XG | Guangxi | Karst |
| 19 | Camellia parvipetala | XB | Guangxi | Non-Karst |
| 20 | Camellia perpetua | SJ | Guangxi | Karst |
| 21 | Camellia pingguoensis | PG | Guangxi | Karst |
| 22 | Camellia pingguoensis var. terminalis | DS | Guangxi | Karst |
| 23 | Camellia ptilosperma | MZ | Guangxi | Karst |
| 24 | Camellia pubipetala | MB | Guangxi | Karst |
| 25 | Camellia quinqueloculosa | WS | Guangxi | Karst |
| 26 | Camellia rostrata | HG | Guangxi | Karst |
| 27 | Camellia tianeensis | TE | Guangxi | Karst |
| 28 | Camellia tunghinensis | DX | Guangxi | Non-Karst |
| 29 | Camellia wumingensis | WM | Guangxi | Karst |
| 30 | Camellia xiashiensis | XS | Guangxi | Karst |
| Species | TPCg/100 g | TEAg/100 g | TACg/100 g | TFCg/100 g |
|---|---|---|---|---|
| C. mingii | 4.563 ± 0.049 m | 0.048 ± 0.001 g | 3.950 ± 0.006 n | 4.064 ± 0.003 i |
| C. fascicularis | 1.603 ± 0.009 d | 0.036 ± 0.001 ef | 1.743 ± 0.005 i | 6.970 ± 0.012 o |
| C. wumingensis | 2.310 ± 0.015 g | 0.024 ± 0.0003 cd | 3.450 ± 0.006 m | 4.661 ± 0.002 l |
| C. tunghinensis | 0.600 ± 0.012 a | 0.002 ± 0.0003 a | 0.004 ± 0.001 a | 0.368 ± 0.001 a |
| C. rostrata | 1.217 ± 0.045 b | 0.028 ± 0.001 de | 0.479 ± 0.001 c | 1.083 ± 0.003 b |
| C. quinqueloculosa | 3.220 ± 0.047 k | 0.001 ± 0.0003 a | 5.720 ± 0.012 q | 8.133 ± 0.044 s |
| C. pubipetala | 2.807 ± 0.003 i | 0.002 ± 0.001 ab | 4.460 ± 0.058 o | 10.718 ± 0.010 t |
| C. pingguoensis | 1.403 ± 0.004 c | 0.039 ± 0.017 fg | 0.790 ± 0.005 d | 1.530 ± 0.003 c |
| C. pingguoensis var. terminalis | 1.803 ± 0.003 e | 0.061 ± 0.001 h | 0.937 ± 0.002 e | 1.874 ± 0.010 d |
| C. perpetua | 3.253 ± 0.029 k | 0.071 ± 0.0003 h | 2.430 ± 0.005 k | 4.437 ± 0.033 j |
| C. nitidissima var. microcurpa | 0.617 ± 0.017 a | 0.002 ± 0.0003 ab | 0.213 ± 0.009 b | 2.594 ± 0.004 f |
| C. nitidissima | 1.410 ± 0.006 c | 0.006 ± 0.0003 ab | 0.801 ± 0.006 d | 2.614 ± 0.002 f |
| C. longzhouensis | 2.405 ± 0.003 h | 0.045 ± 0.0003 fg | 2.282 ± 0.002 j | 5.352 ± 0.002 m |
| C. limonia | 2.070 ± 0.035 f | 0.090 ± 0.001 i | 1.315 ± 0.003 g | 4.573 ± 0.001 k |
| C. impressinervis | 2.810 ± 0.006 i | 0.002 ± 0.0003 a | 6.560 ± 0.058 s | 7.590 ± 0.006 q |
| C. huana | 0.600 ± 0.006 a | 0.002 ± 0.0011 ab | 0.004 ± 0.0003 a | 2.2763 ± 0.003 e |
| C. flavida | 3.016 ± 0.017 j | 0.011 ± 0.001 ab | 4.943 ± 0.009 p | 7.013 ± 0.003 p |
| C. euphlebia | 2.403 ± 0.004 h | 0.070 ± 0.001 h | 1.044 ± 0.001 f | 3.587 ± 0.007 h |
| C. debaoensis | 1.403 ± 0.004 c | 0.014 ± 0.001 bc | 1.467 ± 0.005 h | 5.838 ± 0.002 n |
| C. chrysanthoides | 2.805 ± 0.006 i | 0.083 ± 0.003 i | 3.276 ± 0.018 l | 5.840 ± 0.026 n |
| C. achrysantha | 4.203 ± 0.004 l | 0.072 ± 0.001 h | 3.500 ± 0.012 m | 3.497 ± 0.012 g |
| Principal Component Number | Eigenvalue | Percentage of Variance (%) | Cumulative (%) |
|---|---|---|---|
| 1 | 2.359 | 58.984 | 58.983 |
| 2 | 1.235 | 30.880 | 89.864 |
| 3 | 0.335 | 8.364 | 98.228 |
| 4 | 0.071 | 1.772 | 100 |
| Active Ingredient Index | Factor Load Value | Component Score Coefficient Matrix | ||
|---|---|---|---|---|
| F1 | F2 | F1 | F2 | |
| TPC | 0.903 | 0.336 | 0.903 | 0.336 |
| TEA | 0.182 | 0.925 | 0.182 | 0.925 |
| TAC | 0.958 | −0.120 | 0.958 | −0.120 |
| TFC | 0.701 | −0.510 | 0.701 | −0.510 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, J.; Zou, R.; Yang, Y.; Jiang, Y.; Wang, F.; Chai, S.; Wei, X. Genotypic and Phenotypic Diversity of Endemic Golden Camellias Collected from China. Diversity 2023, 15, 1134. https://doi.org/10.3390/d15111134
Tang J, Zou R, Yang Y, Jiang Y, Wang F, Chai S, Wei X. Genotypic and Phenotypic Diversity of Endemic Golden Camellias Collected from China. Diversity. 2023; 15(11):1134. https://doi.org/10.3390/d15111134
Chicago/Turabian StyleTang, Jianmin, Rong Zou, Yishan Yang, Yunsheng Jiang, Faming Wang, Shengfeng Chai, and Xiao Wei. 2023. "Genotypic and Phenotypic Diversity of Endemic Golden Camellias Collected from China" Diversity 15, no. 11: 1134. https://doi.org/10.3390/d15111134