

Still Many to Be Named: An Integrative Analysis of the Genus Dendronotus (Gastropoda: Nudibranchia) in the North Pacific Revealed Seven New Species

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
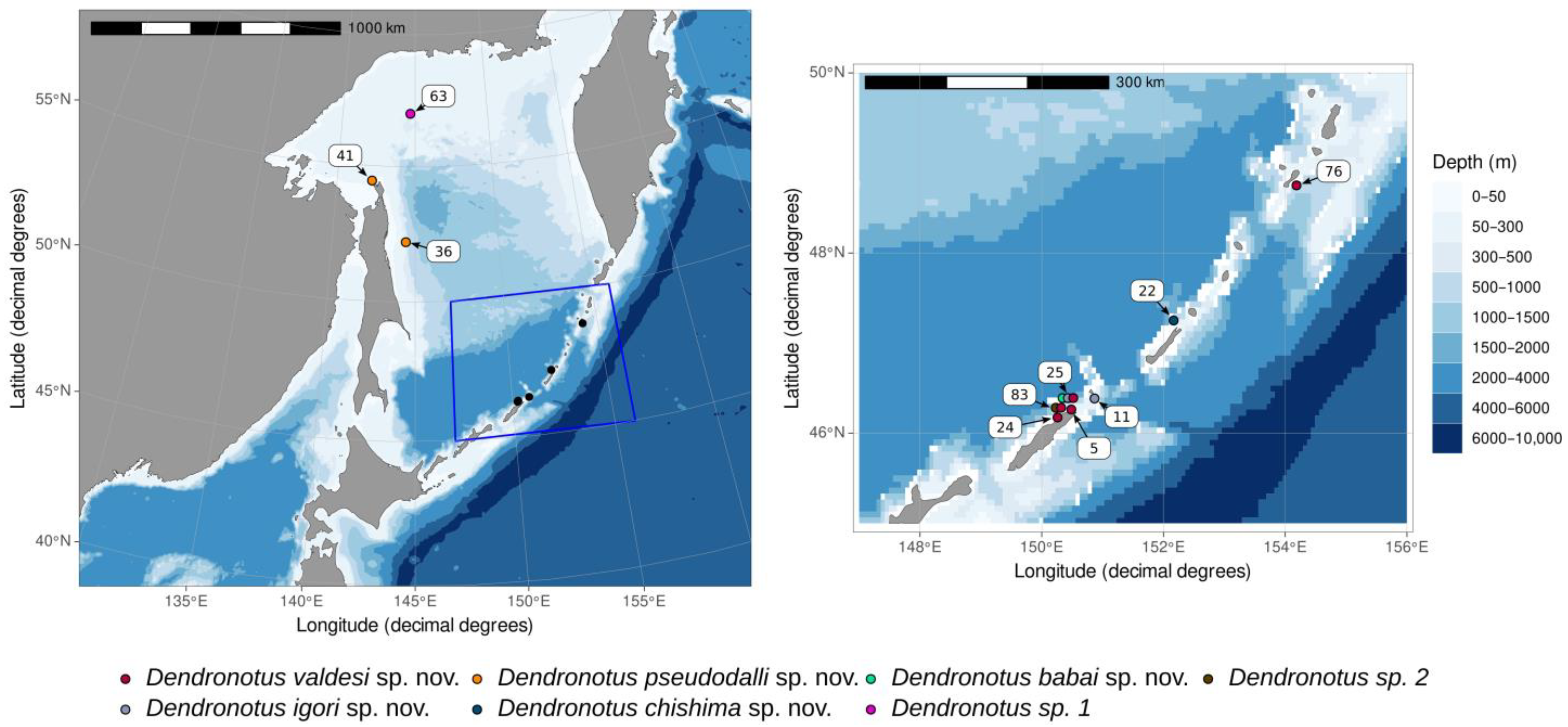
2.1. Collection Data and Community Descriptions
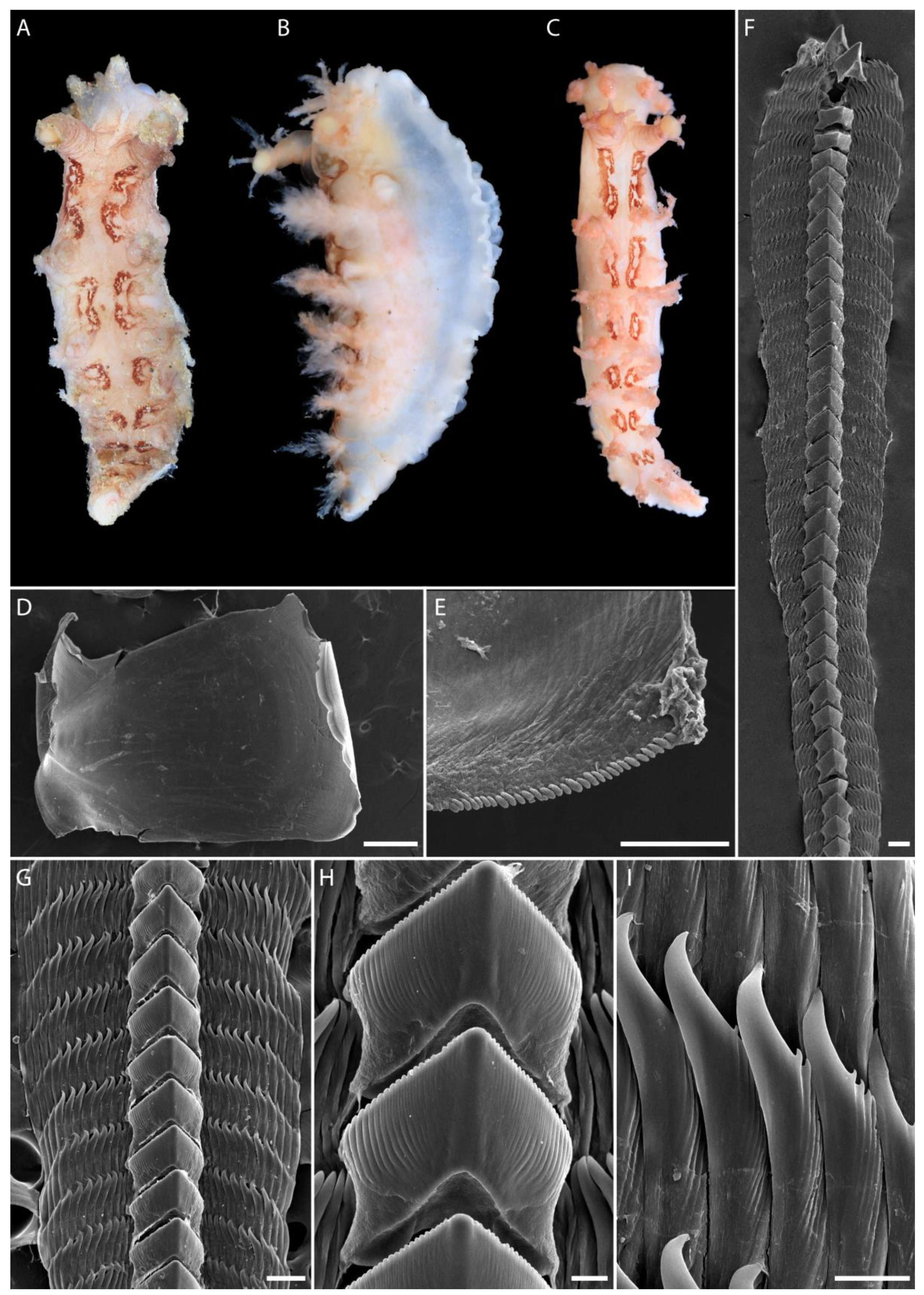
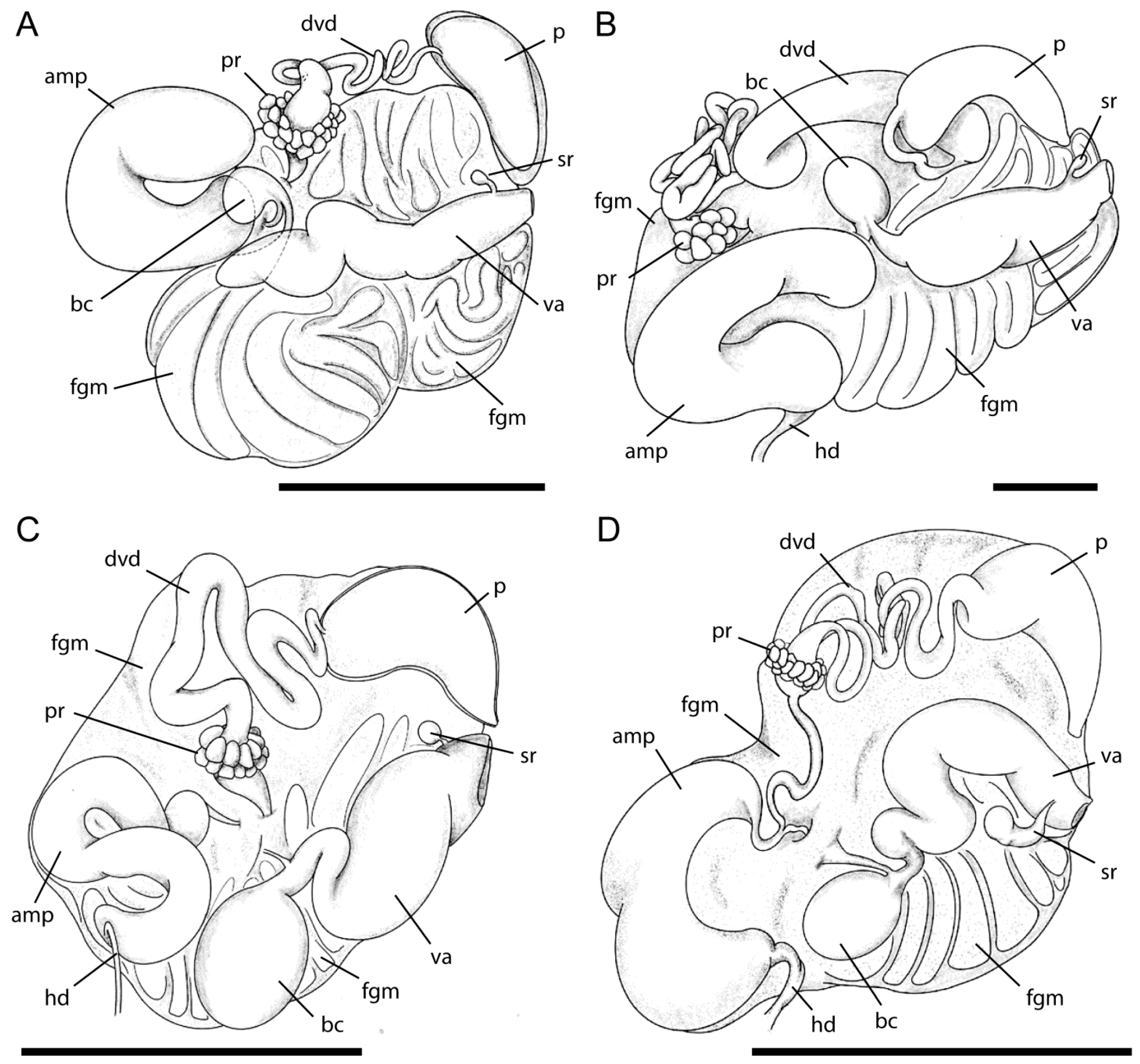
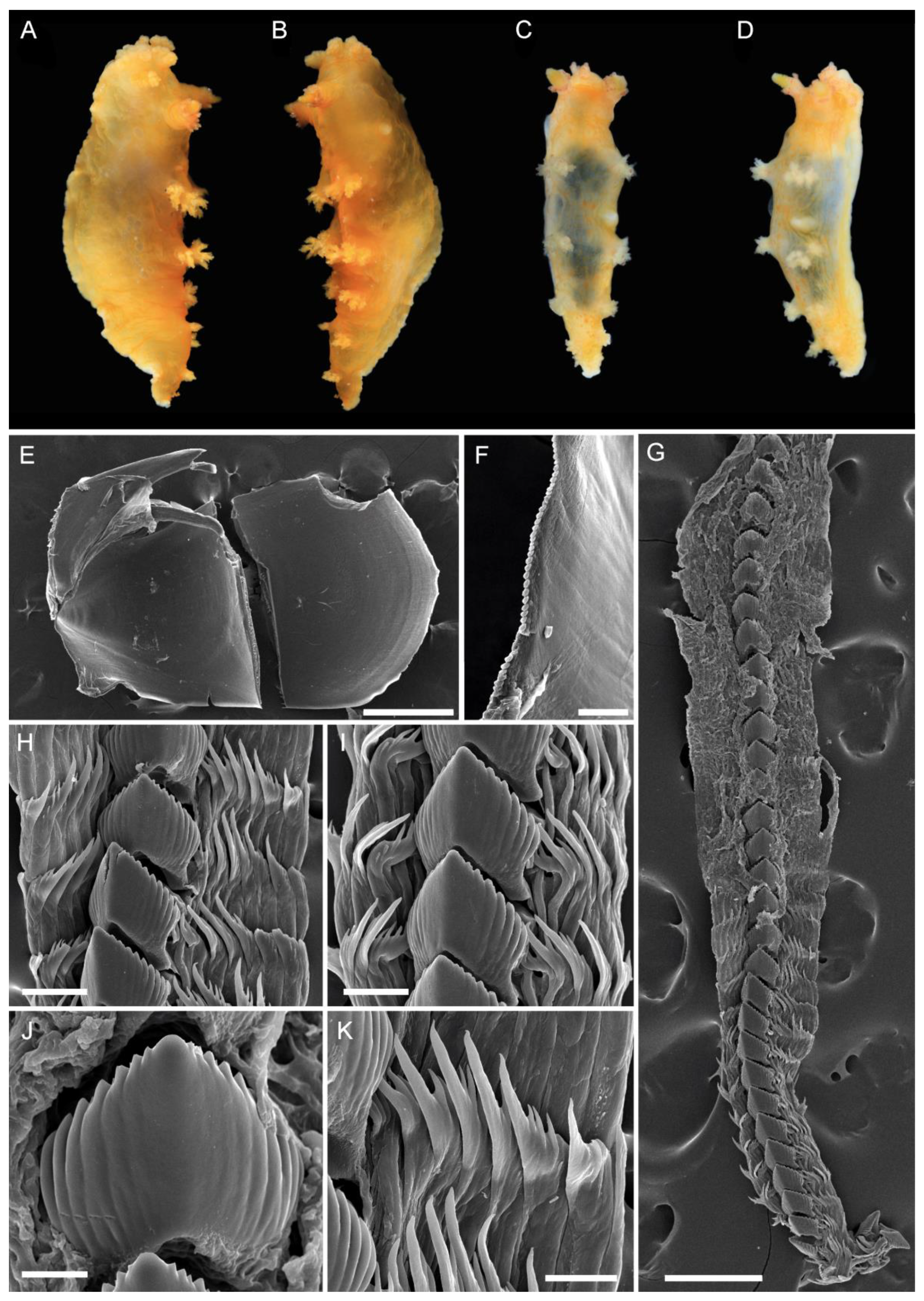
2.2. Morphological Studies
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Data Processing and Phylogenetic Reconstruction
2.5. Species Delimitation
2.6. Ancestral Area Reconstruction
2.7. Nomenclatural Acts
3. Results
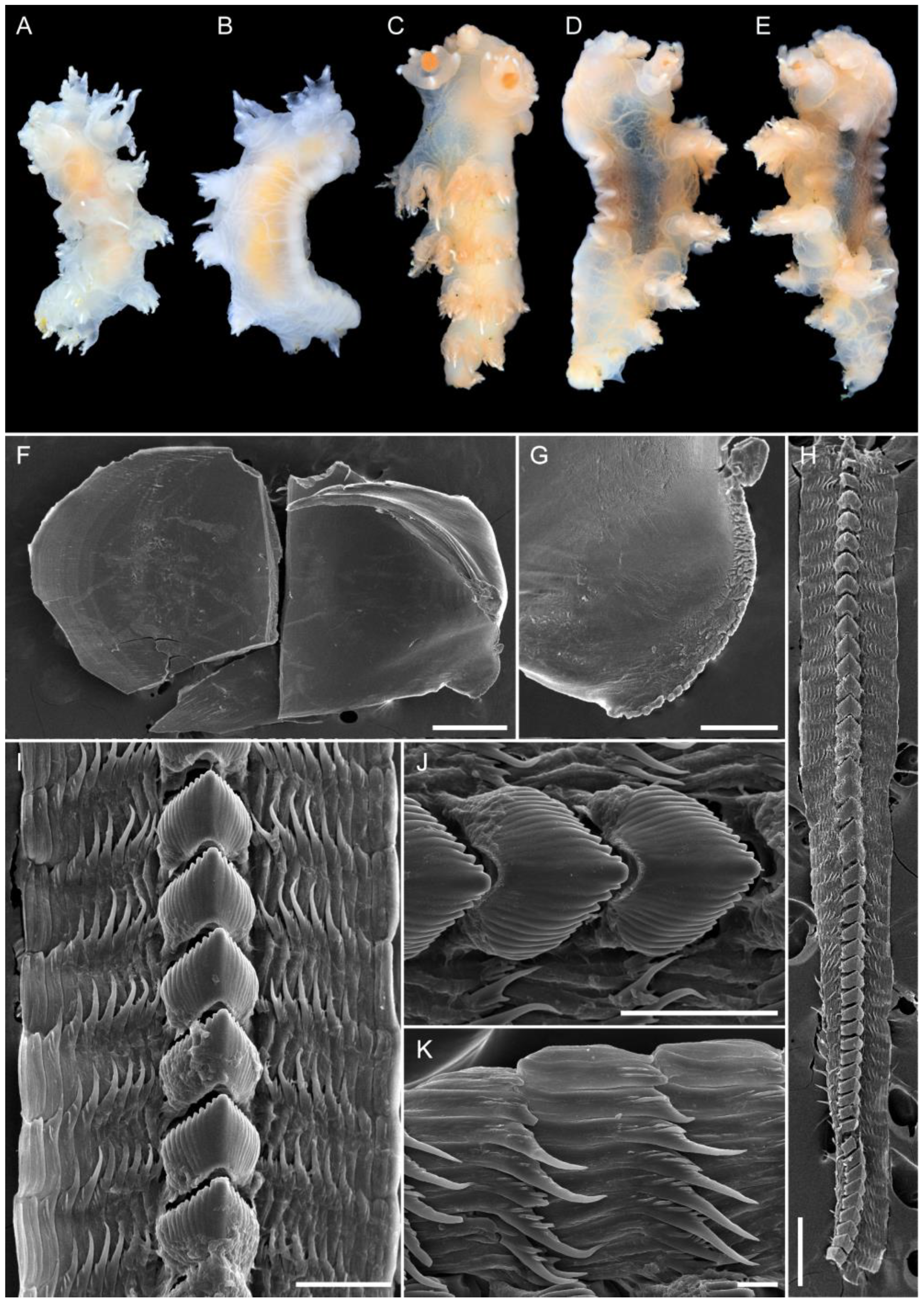
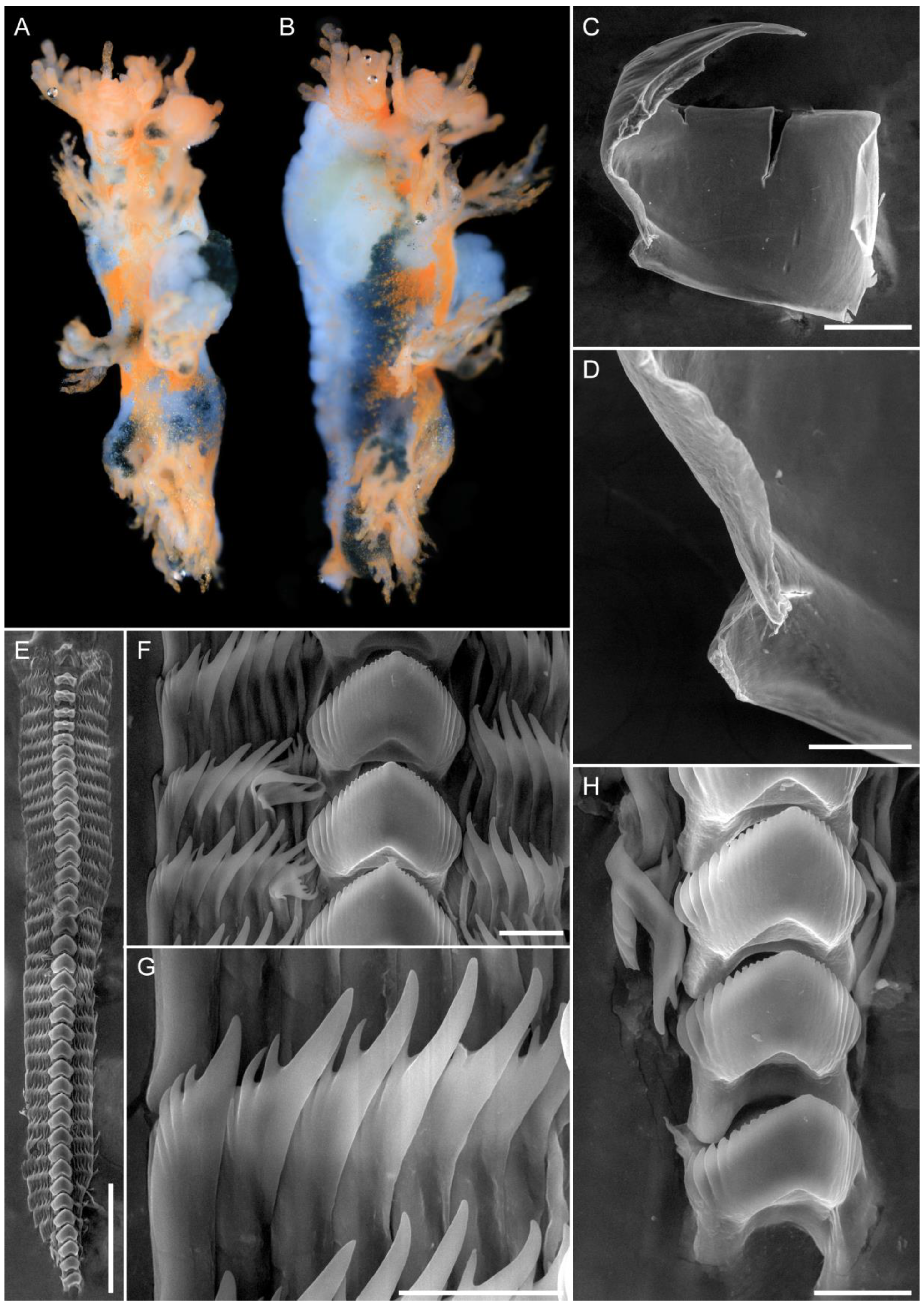
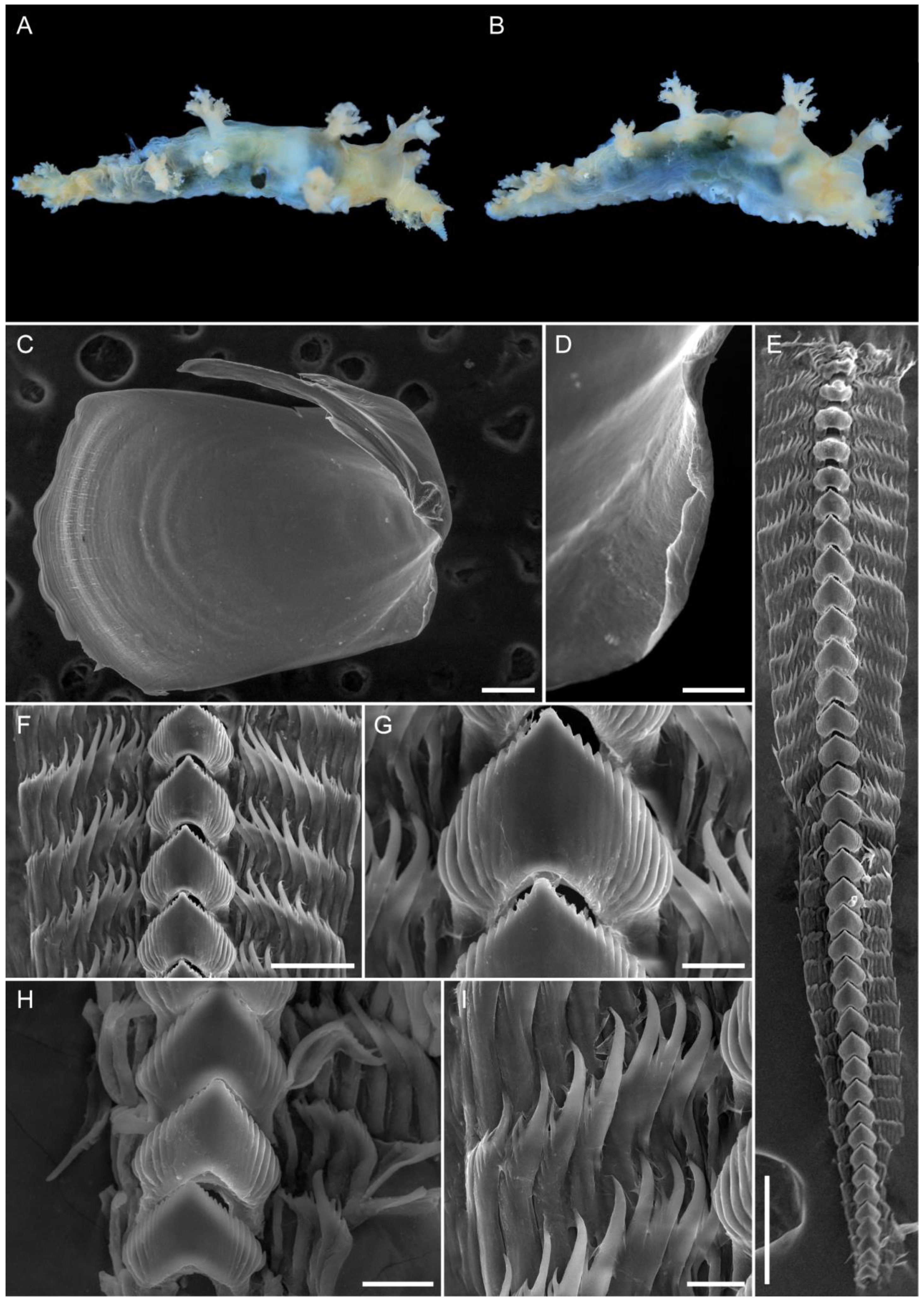
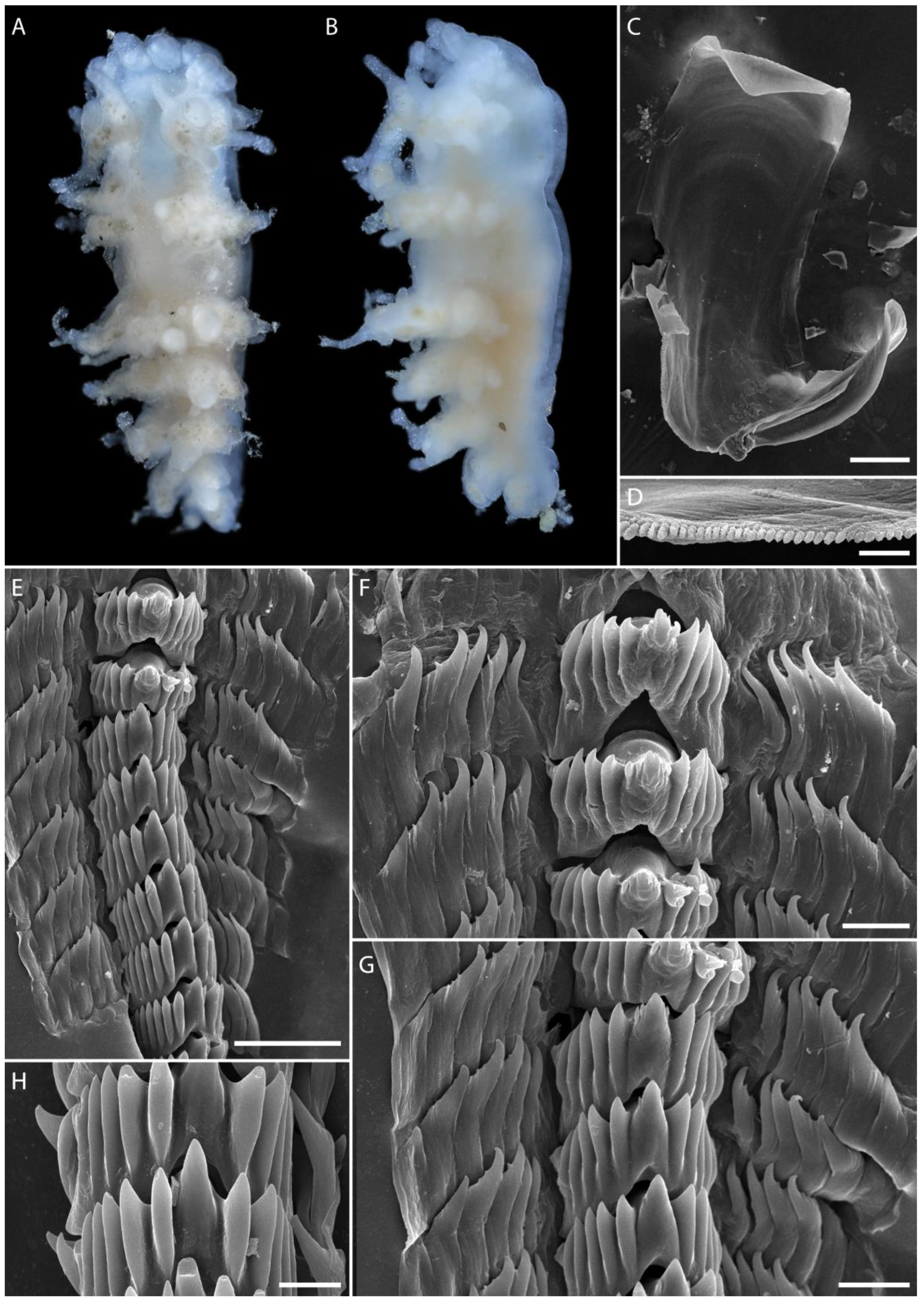
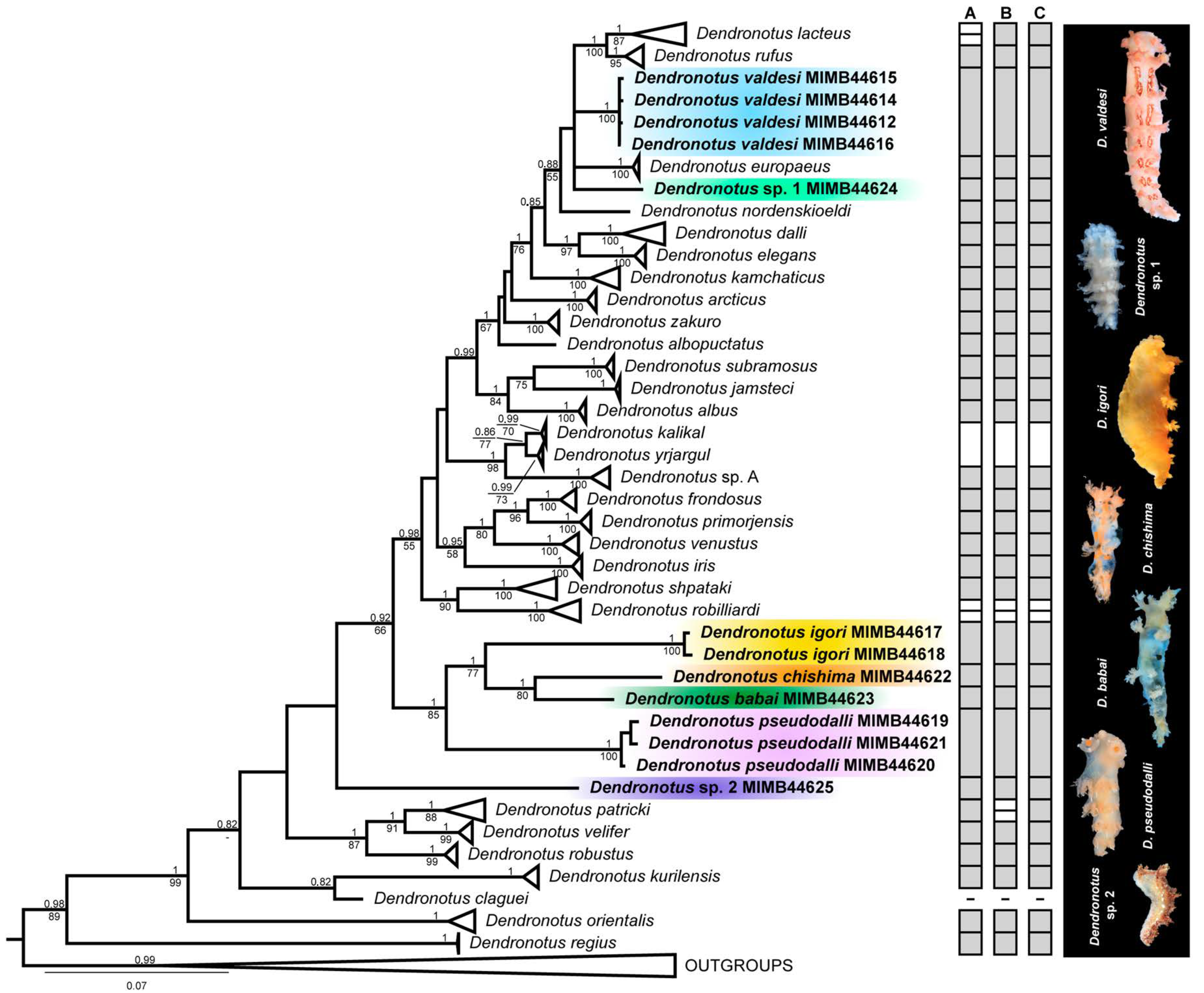
3.1. Taxonomy
3.2. Molecular Phylogenetic and Species Delimitation Analyses
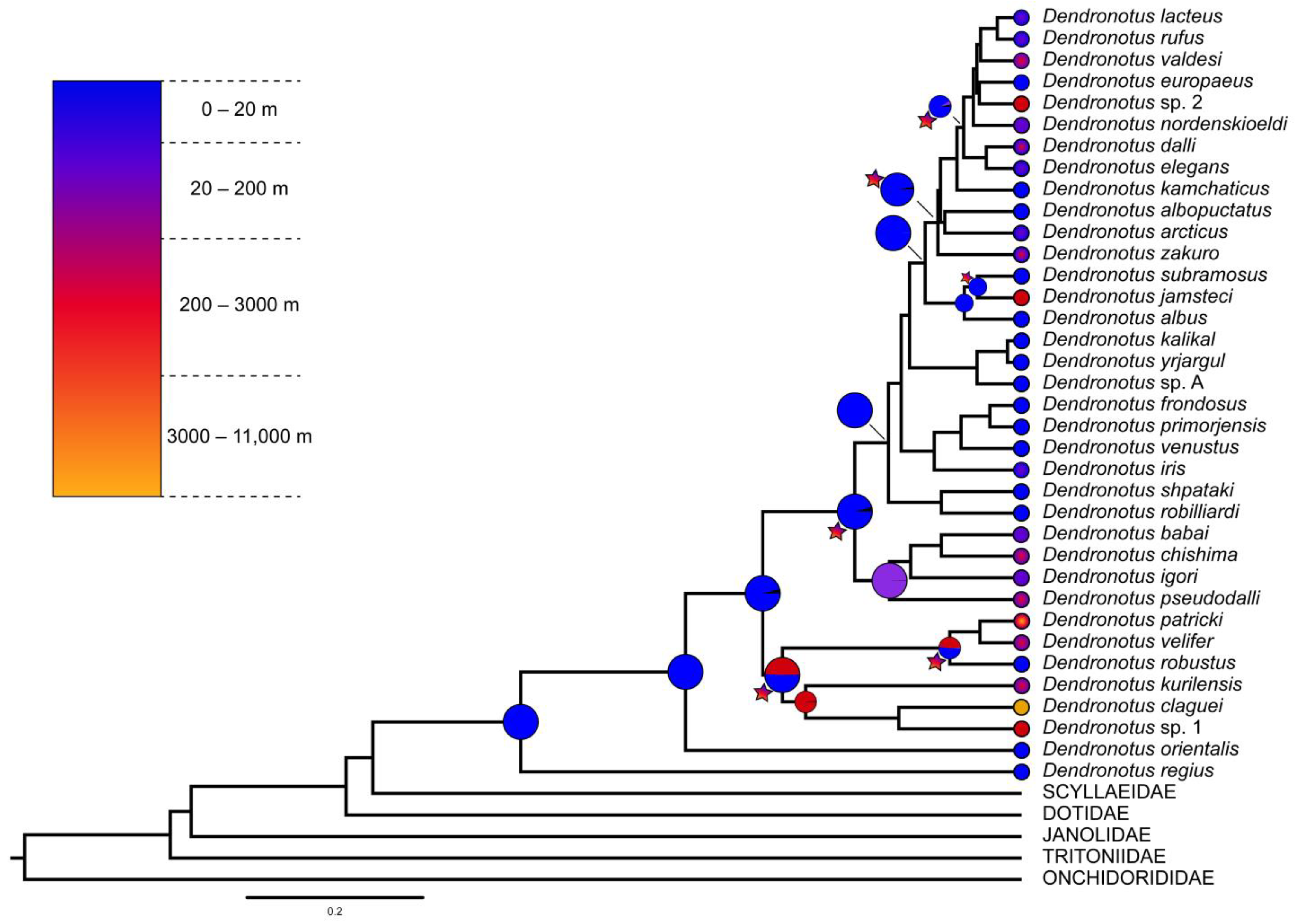
3.3. Ancestral Area Reconstruction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mentges, A.; Blowes, S.A.; Hodapp, D.; Hillebrand, H.; Chase, J.M. Effects of site-selection bias on estimates of biodiversity change. Conserv. Biol. 2021, 35, 688–698. [Google Scholar] [CrossRef]
- Gray, J.S. Marine diversity: The paradigms in patterns of species richness examined. Sci. Mar. 2001, 65, 41–56. [Google Scholar] [CrossRef]
- Mcclain, C.R.; Rex, M.A.; Etter, R.J. Patterns In Deep-Sea Macroecology. In Marine Macroecology; Witman, J.D., Kaustuv, R., Eds.; University of Chicago Press: Chicago, IL, USA, 2009; pp. 65–100. [Google Scholar]
- Ayma, A.; Aguzzi, J.; Canals, M.; Lastras, G.; Bahamon, N.; Mechó, A.; Company, J. Comparison between ROV video and Agassiz trawl methods for sampling deep water fauna of submarine canyons in the Northwestern Mediterranean Sea with observations on behavioural reactions of target species. Deep-Sea Res. Part I 2016, 114, 149–159. [Google Scholar] [CrossRef]
- Ekimova, I.; Valdés, Á.; Stanovova, M.; Mikhlina, A.; Antokhina, T.; Neretina, T.; Chichvarkhina, O.; Schepetov, D. Connected across the ocean: Taxonomy and biogeography of deep-water Nudibranchia from the Northwest Pacific reveal trans-Pacific links and two undescribed species. Org. Divers. Evol. 2021, 21, 753–782. [Google Scholar] [CrossRef]
- Valdés, Á.; Lundsten, L.; Wilson, N.G. Five new deep-sea species of nudibranchs (Gastropoda: Heterobranchia: Cladobranchia) from the Northeast Pacific. Zootaxa 2018, 4526, 401–433. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, T.D.; Rowden, A.A.; Bax, N.J. A southern hemisphere bathyal fauna is distributed in latitudinal bands. Curr. Biol. 2011, 21, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Baco, A.; Smith, C.; Peek, A.; Roderick, G.; Vrijenhoek, R. The phylogenetic relationships of whale-fall vesicomyid clams based on mitochondrial COI DNA sequences. Mar. Ecol. Prog. Ser. 1999, 182, 137–147. [Google Scholar] [CrossRef]
- Lindner, A.; Cairns, S.D.; Cunningham, C.W. From offshore to onshore: Multiple origins of shallow-water corals from deep-sea ancestors. PLoS ONE 2008, 3, e2429. [Google Scholar] [CrossRef]
- Raupach, M.J.; Mayer, C.; Malyutina, M.; Wägele, J.-W. Multiple origins of deep-sea Asellota (Crustacea: Isopoda) from shallow waters revealed by molecular data. Proc. R. Soc. B Boil. Sci. 2009, 276, 799–808. [Google Scholar] [CrossRef]
- Pola, M.; Stout, C.C. Descriptions of the first two tropical Indo-Pacific species of Dendronotus (Gastropoda: Nudibranchia) with new data for the poorly known species Dendronotus gracilis Baba, 1949. Zootaxa 2008, 1960, 45–66. [Google Scholar] [CrossRef]
- Ekimova, I.; Valdés, Á.; Chichvarkhin, A.; Antokhina, T.; Lindsay, T.; Schepetov, D. Diet-driven ecological radiation and allopatric speciation result in high species diversity in a temperate-cold water marine genus Dendronotus (Gastropoda: Nudibranchia). Mol. Phylogenet. Evol. 2019, 141, 106609. [Google Scholar] [CrossRef]
- Ekimova, I.; Korshunova, T.; Schepetov, D.; Neretina, T.; Sanamyan, N.; Martynov, A. Integrative systematics of northern and Arctic nudibranchs of the genus Dendronotus (Mollusca, Gastropoda), with descriptions of three new species. Zool. J. Linn. Soc.-Lond. 2015, 173, 841–886. [Google Scholar] [CrossRef]
- Ekimova, I.; Mikhlina, A.; Antokhina, T.; Schepetov, D. A description of Dendronotus shpataki sp. nov.(Gastropoda: Nudibranchia) from the Sea of Japan: A contribution of citizen science to marine zoology. Zoosyst. Ross. 2022, 31, 3–19. [Google Scholar] [CrossRef]
- Ekimova, I.; Schepetov, D.; Chichvarkhina, O.; Chichvarkhin, A.Y. Nudibranch molluscs of the genus Dendronotus Alder et Hancock, 1845 (Heterobranchia: Dendronotina) from Northwestern Sea of Japan with description of a new species. Invertebr. Zool. 2016, 13, 15–42. [Google Scholar] [CrossRef]
- Ekimova, I.; Valdés, Á.; Schepetov, D.; Chichvarkhin, A. Was Gordon Robilliard right? Integrative systematics suggest that Dendronotus diversicolor (multicolor frond-aeolis) is a valid species. Can. J. Zool. 2016, 94, 793–799. [Google Scholar] [CrossRef]
- Korshunova, T.; Bakken, T.; Grøtan, V.V.; Johnson, K.B.; Lundin, K.; Martynov, A. A synoptic review of the family Dendronotidae (Mollusca: Nudibranchia): A multilevel organismal diversity approach. Contrib. Zool. 2020, 90, 93–153. [Google Scholar] [CrossRef]
- Korshunova, T.; Martynov, A.; Bakken, T.; Picton, B. External diversity is restrained by internal conservatism: New nudibranch mollusc contributes to the cryptic species problem. Zool. Scr. 2017, 46, 683–692. [Google Scholar] [CrossRef]
- Stout, C.C.; Pola, M.; Valdés, Á. Phylogenetic analysis of Dendronotus nudibranchs with emphasis on northeastern Pacific species. J. Molluscan Stud. 2010, 76, 367–375. [Google Scholar] [CrossRef]
- Lundin, K.; Korshunova, T.; Malmberg, K.; Martynov, A. Intersection of historical museum collections and modern systematics: A relict population of the Arctic nudibranch Dendronotus velifer G.O. Sars, 1878 in a Swedish fjord. Contrib. Zool. 2017, 86, 303–318. [Google Scholar] [CrossRef]
- Martynov, A.; Fujiwara, Y.; Tsuchida, S.; Sanamyan, N.; Sanamyan, K.; Fletcher, K.; Korshunova, T. Three new species of the genus Dendronotus from Japan and Russia (Mollusca, Nudibranchia). Zootaxa 2020, 4747, 495–513. [Google Scholar] [CrossRef] [PubMed]
- Stout, C.C.; Wilson, N.G.; Valdés, Á. A new species of deep-sea Dendronotus Alder & Hancock (Mollusca: Nudibranchia) from California, with an expanded phylogeny of the genus. Invertebr. Syst. 2011, 25, 60–69. [Google Scholar]
- Valdés, Á.; Bouchet, P. Naked in toxic fluids: A nudibranch mollusc from hydrothermal vents. Deep-Sea Res. Part II 1998, 45, 319–327. [Google Scholar] [CrossRef]
- MolluscaBase. Available online: https://www.molluscabase.org/ (accessed on 29 November 2022).
- Ivanova, N.V.; Dewaard, J.R.; Hebert, P.D. An inexpensive, automation-friendly protocol for recovering high-quality DNA. Mol. Ecol. Notes 2006, 6, 998–1002. [Google Scholar] [CrossRef]
- Ekimova, I.; Deart, Y.; Antokhina, T.; Mikhlina, A.; Schepetov, D. Stripes matter: Integrative systematics of Coryphellina rubrolineata species complex (Gastropoda: Nudibranchia) from Vietnam. Diversity 2022, 14, 294. [Google Scholar] [CrossRef]
- Ekimova, I.; Valdés, Á.; Malaquias, M.A.E.; Rauch, C.; Chichvarkhin, A.; Mikhlina, A.; Antokhina, T.; Chichvarkhina, O.; Schepetov, D. High-level taxonomic splitting in allopatric taxa causes confusion downstream: A revision of the nudibranch family Coryphellidae. Zool. J. Linn. Soc.-Lond. 2022, 196, 215–249. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef]
- Chaban, E.M.; Ekimova, I.A.; Schepetov, D.M.; Chernyshev, A.V. Meloscaphander grandis (Heterobranchia: Cephalaspidea), a deep-water species from the North Pacific: Redescription and taxonomic remarks. Zootaxa 2019, 4646, 385–400. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting species using single-locus data and the Generalized Mixed Yule Coalescent approach: A revised method and evaluation on simulated data sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLOS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Harris, A.; He, X. S-DIVA (Statistical Dispersal-Vicariance Analysis): A tool for inferring biogeographic histories. Mol. Phylogenet. Evol. 2010, 56, 848–850. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Matzke, N.J. Founder-event speciation in BioGeoBEARS package dramatically improves likelihoods and alters parameter inference in Dispersal-Extinction-Cladogenesis (DEC) analyses. Front. Biogeogr. 2012, 4, 210. [Google Scholar]
- Matzke, N.J. Probabilistic Historical Biogeography: New Models for Founder-Event Speciation, Imperfect Detection, and Fossils Allow Improved Accuracy and Model-Testing. Ph.D. Thesis, University of California, Berkeley, CA, USA, 2013. [Google Scholar]
- Komisarenko, A.; Mordukhovich, V.; Ekimova, I.; Imbs, A. A comparison of food sources of nudibranch mollusks at different depths off the Kuril Islands using fatty acid trophic markers. PeerJ 2021, 9, e12336. [Google Scholar] [CrossRef]
- Robilliard, G. The systematics and some aspects of the ecology of the genus Dendronotus. Veliger 1970, 12, 433–479. [Google Scholar]
- Ekimova, I.; Malakhov, V. Progenesis in the evolution of the nudibranch mollusks genus Dendronotus (Gastropoda: Nudibranchia). Dokl. Biol. Sci. 2016, 467, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L.; Waddell, P.J.; Huelsenbeck, J.P.; Foster, P.G.; Lewis, P.O.; Rogers, J.S. Bias in phylogenetic estimation and its relevance to the choice between parsimony and likelihood methods. Syst. Biol. 2001, 50, 525–539. [Google Scholar] [CrossRef]
- Laurin, M. The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I. Systematics, middle ear evolution and jaw suspension. Ann. Sci. Nat. Zool. 1998, 19, 1–42. [Google Scholar] [CrossRef]
- Spiridonov, V.A.; Neretina, T.V.; Schepetov, D. Morphological characterization and molecular phylogeny of Portunoidea Rafinesque, 1815 (Crustacea Brachyura): Implications for understanding evolution of swimming capacity and revision of the family-level classification. Zool. Anz. 2014, 253, 404–429. [Google Scholar] [CrossRef]
- Geiger, D.L.; Thacker, C.E. Molecular phylogeny of Vetigastropoda reveals non-monophyletic Scissurellidae, Trochoidea, and Fissurelloidea. Molluscan Res. 2005, 25, 47–55. [Google Scholar]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ekimova, I.A.; Mikhlina, A.L.; Stanovova, M.V.; Krupitskaya, N.R.; Chichvarkhina, O.V.; Schepetov, D.M. Still Many to Be Named: An Integrative Analysis of the Genus Dendronotus (Gastropoda: Nudibranchia) in the North Pacific Revealed Seven New Species. Diversity 2023, 15, 162. https://doi.org/10.3390/d15020162
Ekimova IA, Mikhlina AL, Stanovova MV, Krupitskaya NR, Chichvarkhina OV, Schepetov DM. Still Many to Be Named: An Integrative Analysis of the Genus Dendronotus (Gastropoda: Nudibranchia) in the North Pacific Revealed Seven New Species. Diversity. 2023; 15(2):162. https://doi.org/10.3390/d15020162
Chicago/Turabian StyleEkimova, Irina A., Anna L. Mikhlina, Maria V. Stanovova, Nellya R. Krupitskaya, Olga V. Chichvarkhina, and Dimitry M. Schepetov. 2023. "Still Many to Be Named: An Integrative Analysis of the Genus Dendronotus (Gastropoda: Nudibranchia) in the North Pacific Revealed Seven New Species" Diversity 15, no. 2: 162. https://doi.org/10.3390/d15020162
APA StyleEkimova, I. A., Mikhlina, A. L., Stanovova, M. V., Krupitskaya, N. R., Chichvarkhina, O. V., & Schepetov, D. M. (2023). Still Many to Be Named: An Integrative Analysis of the Genus Dendronotus (Gastropoda: Nudibranchia) in the North Pacific Revealed Seven New Species. Diversity, 15(2), 162. https://doi.org/10.3390/d15020162