


Bladderwort Relationship to Zooplankton in Two Northern Michigan Lakes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sampling Protocol
2.3. Data Analysis
2.3.1. Analysis 1: Relations between Bladderworts and Zooplankton Prey Items
2.3.2. Analysis 2: Trap and Water Column Relations of Zooplankton
3. Results
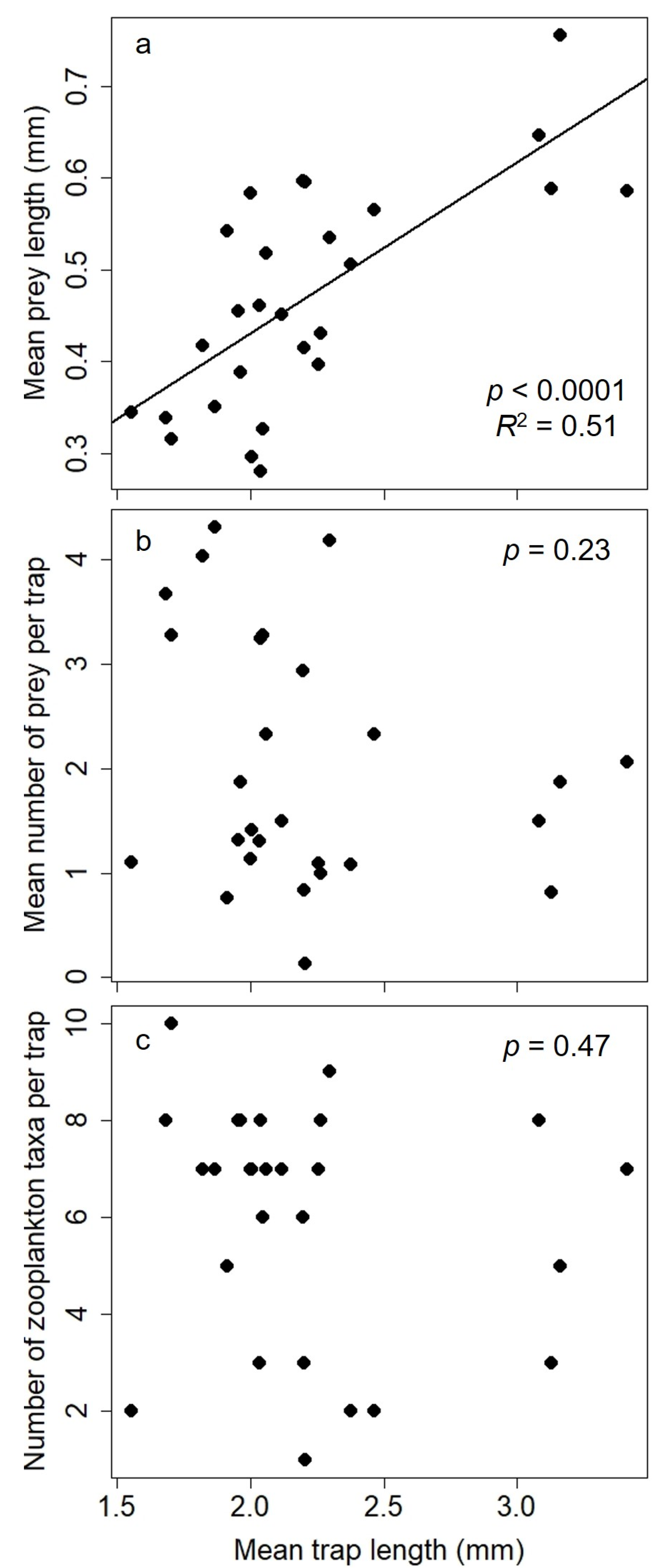
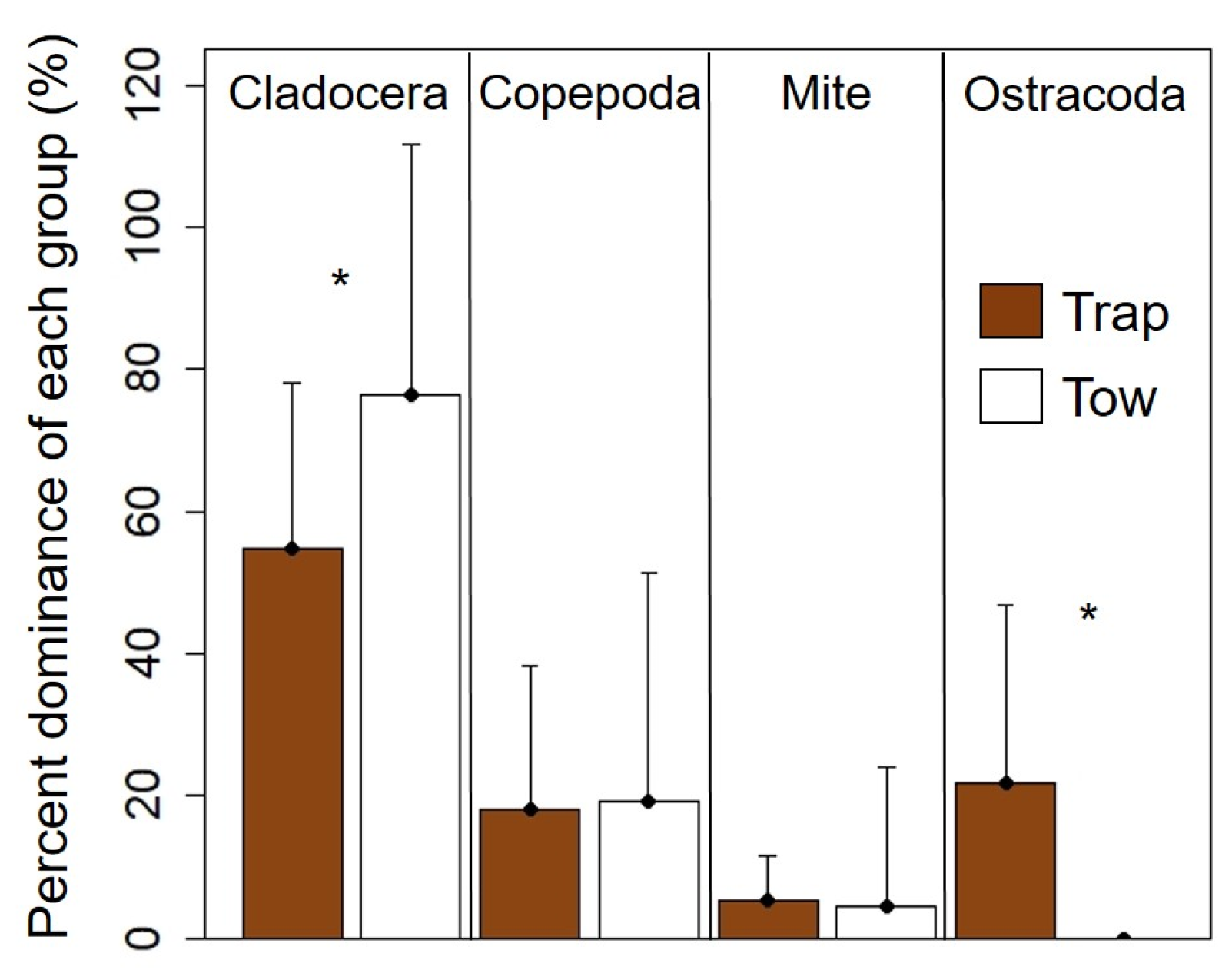
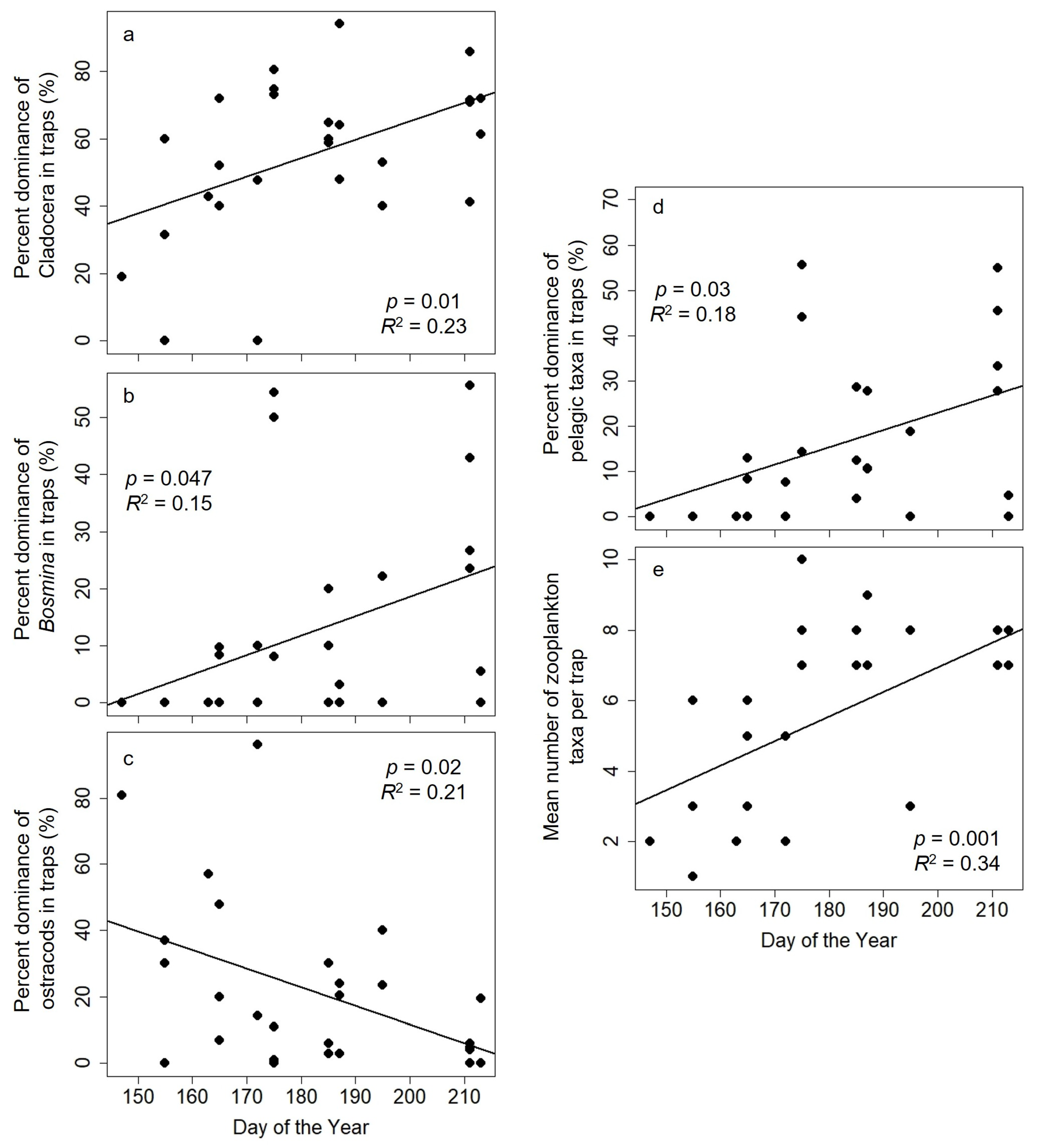
3.1. Analysis 1: Relations between Bladderworts and Zooplankton Prey Items
3.2. Analysis 2: Trap and Water Column Relations of Zooplankton
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenny, J.-P.; Francus, P.; Normandeau, A.; Lapointe, F.; Perga, M.-E.; Ojala, A.; Schimmelmann, A.; Zolitschka, B.; Jenny, J.; Perga, M.; et al. Global spread of hypoxia in freshwater ecosystems during the last three centuries is caused by rising local human pressure. Glob. Chang. Biol. 2016, 22, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W. Recent advances in the understanding and management of eutrophication. Limnol. Oceanogr. 2006, 51, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Woolway, R.I.; Kraemer, B.M.; Lenters, J.D.; Merchant, C.J.; O’Reilly, C.M.; Sharma, S. Global lake responses to climate change. Nat. Rev. Earth Environ. 2020, 1, 388–403. [Google Scholar] [CrossRef]
- Iglesias, C.; Mazzeo, N.; Meerhoff, M.; Lacerot, G.; Clemente, J.M.; Scasso, F.; Kruk, C.; Goyenola, G.; García-Alonso, J.; Amsinck, S.L.; et al. High predation is of key importance for dominance of small-bodied zooplankton in warm shallow lakes: Evidence from lakes, fish exclosures and surface sediments. Hydrobiologia 2011, 667, 133–147. [Google Scholar] [CrossRef]
- Jeppesen, E.; Nõges, P.; Davidson, T.A.; Haberman, J.; Nõges, T.; Blank, K.; Lauridsen, T.L.; Søndergaard, M.; Sayer, C.; Laugaste, R.; et al. Zooplankton as indicators in lakes: A scientific-based plea for including zooplankton in the ecological quality assessment of lakes according to the European Water Framework Directive (WFD). Hydrobiologia 2011, 676, 279–297. [Google Scholar] [CrossRef]
- Yuan, L.L.; Pollard, A.I. Changes in the relationship between zooplankton and phytoplankton biomasses across a eutrophication gradient. Limnol. Oceanogr. 2018, 63, 2493–2507. [Google Scholar] [CrossRef]
- Burks, R.L.; Lodge, D.M.; Jeppesen, E.; Lauridsen, T.L. Diel horizontal migration of zooplankton: Costs and benefits of inhabiting the littoral. Freshw. Biol. 2002, 47, 343–365. [Google Scholar] [CrossRef]
- Doubek, J.P.; Campbell, K.L.; Doubek, K.M.; Hamre, K.D.; Lofton, M.E.; McClure, R.P.; Ward, N.K.; Carey, C.C. The effects of hypolimnetic anoxia on the diel vertical migration of freshwater crustacean zooplankton. Ecosphere 2018, 9, e02332. [Google Scholar] [CrossRef] [Green Version]
- Doubek, J.P.; Goldfarb, S.K.; Stockwell, J.D. Should we be sampling zooplankton at night? Limnol. Oceanogr. Lett. 2020, 5, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Paterson, M. The distribution of microcrustacea in the littoral zone of a freshwater lake. Hydrobiologia 1993, 263, 173–183. [Google Scholar] [CrossRef]
- Walseng, B.; Hessen, D.O.; Halvorsen, G.; Schartau, A.K. Major contribution from littoral crustaceans to zooplankton species richness in lakes. Limnol. Oceanogr. 2006, 51, 2600–2606. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R.; Cochran, P.A.; Elser, J.J.; Elser, M.M.; Lodge, D.M.; Kretchmer, D.; He, X. Regulation of lake primary productivity by food web structure. Ecology 1987, 68, 1863–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinke, K.; Huber, A.M.R.; Kempke, S.; Eder, M.; Wolf, T.; Probst, W.N.; Rothhaupta, K.-O. Lake-wide distributions of temperature, phytoplankton, zooplankton, and fish in the pelagic zone of a large lake. Limnol. Oceanogr. 2009, 54, 1306–1322. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Ludsin, S.A.; Ruberg, S.A.; Höök, T.O.; Pothoven, S.A.; Brandt, S.B.; Lang, G.A.; Liebig, J.R.; Cavaletto, J.F. Hypoxia affects spatial distributions and overlap of pelagic fish, zooplankton, and phytoplankton in Lake Erie. J. Exp. Mar. Biol. Ecol. 2009, 381, S92–S107. [Google Scholar] [CrossRef]
- Hanazato, T.; Yasuno, M. Zooplankton community structure driven by vertebrate and invertebrate predators. Oecologia 1989, 81, 450–458. [Google Scholar] [CrossRef]
- Lane, P.A. Vertebrate and invertebrate predation intensity on freshwater zooplankton communities. Nature 1979, 280, 391–393. [Google Scholar] [CrossRef]
- Winder, M.; Bürgi, H.R.; Spaak, P. Mechanisms regulating zooplankton populations in a high-mountain lake. Freshw. Biol. 2003, 48, 795–809. [Google Scholar] [CrossRef]
- Harms, S. The effect of bladderwort (Utricularia) predation on microcrustacean prey. Freshw. Biol. 2002, 47, 1608–1617. [Google Scholar] [CrossRef]
- Havens, K.E. Summer zooplankton dynamics in the limnetic and littoral zones of a humic acid lake. Hydrobiologia 1991, 215, 21–29. [Google Scholar] [CrossRef]
- Guiral, D.; Rougier, C. Trap size and prey selection of two coexisting bladderwort (Utricularia) species in a pristine tropical pond (French Guiana) at different trophic levels. Ann. Limnol. Int. J. Lim. 2007, 43, 147–159. [Google Scholar] [CrossRef]
- Meyers, D.G.; Strickler, J.R. Capture enhancement in a carnivorous aquatic plant: Function of antennae and bristles in Utricularia vulgaris. Science 1979, 203, 1022–1025. [Google Scholar] [CrossRef] [PubMed]
- Davenport, J.M.; Riley, A.W. Complex inter-Kingdom interactions: Carnivorous plants affect growth of an aquatic vertebrate. J. Anim. Ecol. 2017, 86, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Kurbatova, S.A.; Yershov, I.Y. Crustaceans and rotifers in the predatory feeding of Utricularia. Inland Water Biol. 2009, 2, 271–275. [Google Scholar] [CrossRef]
- Miranda, V.F.O.; Silva, S.R.; Reut, M.S.; Dolsan, H.; Stolarczyk, P.; Rutishauser, R.; Płachno, B.J. A historical perspective of bladderworts (Utricularia): Traps, carnivory and body architecture. Plants 2021, 10, 2656. [Google Scholar] [CrossRef] [PubMed]
- Müller, U.K.; Berg, O.; Schwaner, J.M.; Brown, M.D.; Li, G.; Voesenek, C.J.; van Leeuwen, J.L. Bladderworts, the smallest known suction feeders, generate inertia-dominated flows to capture prey. New Phytol. 2020, 228, 586–595. [Google Scholar] [CrossRef]
- Westermeier, A.S.; Fleischmann, A.; Müller, K.; Schäferhoff, B.; Rubach, C.; Speck, T.; Poppinga, S. Trap diversity and character evolution in carnivorous bladderworts (Utricularia, Lentibulariaceae). Sci. Rep. 2017, 7, 12052. [Google Scholar] [CrossRef] [Green Version]
- Happach, J. Survey of trap contents of four species of bladderworts (Utricularia) from Harlow Lake in Marquette, MI. Conspec. Boreal. 2017, 3, 2. [Google Scholar]
- Englund, G.; Harms, S. Effects of light and microcrustacean prey on growth and investment in carnivory in Utricularia vulgaris. Freshw. Biol. 2003, 48, 786–794. [Google Scholar] [CrossRef] [Green Version]
- Harms, S.; Johansson, F. The influence of prey behaviour on prey selection of the carnivorous plant Utricularia vulgaris. Hydrobiologia 2000, 427, 113–120. [Google Scholar] [CrossRef]
- Kissman, C.E.H.; Knoll, L.B.; Sarnelle, O. Dreissenid mussels (Dreissena polymorpha and Dreissena bugensis) reduce microzooplankton and macrozooplankton biomass in thermally stratified lakes. Limnol. Oceanogr. 2010, 55, 1851–1859. [Google Scholar] [CrossRef]
- Lehman, J.T.; Platte, R.A.; Ferris, J.A. Role of hydrology in development of a vernal clear water phase in an urban impoundment. Freshw. Biol. 2007, 52, 1773–1781. [Google Scholar] [CrossRef]
- Ellwood, N.T.W.; Congestri, R.; Ceschin, S. The role of phytoplankton in the diet of the bladderwort Utricularia australis R.Br. (Lentibulariaceae). Freshw. Biol. 2019, 64, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Balcer, M.D.; Korda, N.L.; Dodson, S.I. Zooplankton of the Great Lakes: A Guide to the Identification and Ecology of the Common Crustacean Species; The University of Wisconsin Press: Madison, WI, USA, 1984. [Google Scholar]
- Ward, H.B.; Whipple, G.C. Fresh-Water Biology; John Wiley & Sons Inc.: New York, NY, USA, 1945. [Google Scholar]
- Downing, J.A.; Rigler, F.H. Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters; Blackwell Scientific Publications: Oxford, UK, 1984. [Google Scholar]
- Bottrell, H.H.; Duncan, A.; Gliwicz, M.Z.; Grygierek, E.; Herzig, A.; Hillbricht-Ilkowska, A.; Kurasawa, H.; Larsson, P.; Weglenska, T. A review of some problems in zooplankton production studies. Norw. J. Zool. 1976, 24, 419–456. [Google Scholar]
- Dumont, H.J.; Van De Velde, I.; Dumont, S. The dry weight estimate of biomass in a selection of Cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental waters. Oecologia 1975, 19, 75–97. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.M.; Rudstam, L.G.; Holeck, K.T. Length Weight Regressions for Zooplankton Biomass Calculations—A Review and a Suggestion for Standard Equations. eCommons Cornell. 2011. Available online: http://hdl.handle.net/1813/24566 (accessed on 1 December 2022).
- Chick, J.H.; Levchuk, A.P.; Medley, K.A.; Havel, J.H. Underestimation of rotifer abundance a much greater problem than previously appreciated. Limnol. Oceanogr. Meth. 2010, 8, 79–87. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Barnett, A.J.; Finlay, K.; Beisner, B.E. Functional diversity of crustacean zooplankton communities: Towards a trait-based classification. Freshw. Biol. 2007, 52, 796–813. [Google Scholar] [CrossRef]
- Chesson, J. The estimation and analysis of preference and its relationship to foraging models. Ecology 1983, 64, 1297–1304. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Lauridsen, T.; Pedersen, L.J.; Jeppesen, E.; Sønergaard, M. The importance of macrophyte bed size for cladoceran composition and horizontal migration in a shallow lake. J. Plankton Res. 1996, 18, 2283–2294. [Google Scholar] [CrossRef] [Green Version]
- Deban, S.M.; Holzman, R.; Müller, U.K. Suction feeding by small organisms: Performance limits in larval vertebrates and carnivorous plants. Integr. Comp. Biol. 2020, 60, 852–863. [Google Scholar] [CrossRef]
- Harms, S. Prey selection in three species of the carnivorous aquatic plant Utricularia (bladderwort). Arch. Hydrobiol. 1999, 146, 449–470. [Google Scholar] [CrossRef]
- Horstmann, M.; Fleischmann, A.; Tollrian, R.; Poppinga, S. Snapshot prey spectrum analysis of the phylogenetically early-diverging carnivorous Utricularia multifida from U. section Polypompholyx (Lentibulariaceae). PLoS ONE 2021, 16, e0249976. [Google Scholar] [CrossRef]
- Whiteside, M.C.; Williams, J.B.; White, C.P. Seasonal abundance and pattern of chydorid, Cladocera in mud and vegetative habitats. Ecology 1978, 59, 1177–1188. [Google Scholar] [CrossRef]
- Berger, S.A.; Diehl, S.; Stibor, H.; Trommer, G.; Ruhenstroth, M.; Wild, A.; Weigert, A.; Jäger, C.G.; Striebel, M. Water temperature and mixing depth affect timing and magnitude of events during spring succession of the plankton. Oecologia 2007, 150, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Sommer, U.; Gliwicz, Z.M.; Lampert, W.; Duncan, A. The PEG-model of seasonal succession of planktonic events in fresh waters. Arch. Hydrobiol. 1986, 106, 433–471. [Google Scholar] [CrossRef]
- Liberto, R.; Mesquita-Joanes, F.; César, I. Dynamics of pleustonic ostracod populations in small ponds on the Island of Martín García (Río de la Plata, Argentina). Hydrobiologia 2011, 688, 47–61. [Google Scholar] [CrossRef]
- van de Bund, W.J.; Davids, C.; Spaas, S.J.H. Seasonal dynamics and spatial distribution of chydorid cladocerans in relation to chironomid larvae in the sandy littoral zone of an oligo-mesotrophic lake. Hydrobiologia 1995, 299, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Ceschin, S.; Bellini, A.; Salituro, A.; Traversetti, L.; Ellwood, N.T.W. Is the capture of invertebrate prey by the aquatic carnivorous plant Utricularia australis selective? Plant Biosyst. 2022, 156, 572–580. [Google Scholar] [CrossRef]
- Knight, S.E.; Frost, T.M. Bladder control in Utricularia macrorhiza: Lake-specific variation in plant investment in carnivory. Ecology 1991, 72, 728–734. [Google Scholar] [CrossRef]
- Bachmann, R.W.; Jones, B.L.; Fox, D.D.; Hoyer, M.; Bull, L.A.; Canfield Jr., D.E. Relations between trophic state indicators and fish in Florida (U.S.A.) lakes. Can. J. Fish. Aquat. Sci. 1996, 53, 842–855. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J.; Wiecko, A.; Górniak, A.; Cudowski, A. A place in space—The horizontal vs vertical factors that influence zooplankton (Rotifera, Crustacea) communities in a mesotrophic lake. J. Limnol. 2019, 78, 243–258. [Google Scholar] [CrossRef]
- Adrian, R.; O’Reilly, C.M.; Zagarese, H.; Baines, S.B.; Hessen, D.O.; Keller, W.; Livingstone, D.M.; Sommaruga, R.; Straile, D.; Van Donk, E.; et al. Lakes as sentinels of climate change. Limnol. Oceanogr. 2009, 54, 2283–2297. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Intercept Value ± 1 S.E. | Slope Value ± 1 S.E. | p-Value | R2 |
|---|---|---|---|---|
| Broad groups | ||||
| Cladocera | −44.7 ± 36.8 | 0.5 ± 0.2 | 0.01 | 0.23 |
| Copepoda | 36.5 ± 36.3 | −0.1 ± 0.2 | 0.62 | 0.01 |
| Mite | −15.4 ± 10.3 | 0.1 ± 0.1 | 0.053 | 0.14 |
| Ostracoda | 124 ± 40.1 | −0.6 ± 0.2 | 0.02 | 0.21 |
| Specific groups | ||||
| Bosmina | −49.4 ± 30.1 | 0.3 ± 0.2 | 0.047 | 0.15 |
| Chydorids | 101 ± 56.2 | −0.2 ± 0.3 | 0.54 | 0.02 |
| Daphnia | −22.4 ± 18.6 | 0.2 ± 0.1 | 0.13 | 0.09 |
| Eurycercus | 7.6 ± 20.9 | −0.02 ± 0.1 | 0.83 | 0.00 |
| Ophryoxus | −7.5 ± 14.5 | 0.1 ± 0.1 | 0.47 | 0.02 |
| Other | ||||
| Pelagic taxa | −53.0 ± 30.6 | 0.4 ± 0.2 | 0.03 | 0.18 |
| Taxa richness | −7.1 ± 3.6 | 0.07 ± 0.02 | 0.001 | 0.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wallace Hoffelt, R.K.; Doubek, J.P. Bladderwort Relationship to Zooplankton in Two Northern Michigan Lakes. Diversity 2023, 15, 171. https://doi.org/10.3390/d15020171
Wallace Hoffelt RK, Doubek JP. Bladderwort Relationship to Zooplankton in Two Northern Michigan Lakes. Diversity. 2023; 15(2):171. https://doi.org/10.3390/d15020171
Chicago/Turabian StyleWallace Hoffelt, Reo K., and Jonathan P. Doubek. 2023. "Bladderwort Relationship to Zooplankton in Two Northern Michigan Lakes" Diversity 15, no. 2: 171. https://doi.org/10.3390/d15020171
APA StyleWallace Hoffelt, R. K., & Doubek, J. P. (2023). Bladderwort Relationship to Zooplankton in Two Northern Michigan Lakes. Diversity, 15(2), 171. https://doi.org/10.3390/d15020171