

Consistent Monthly Reproduction and Completion of a Brooding Coral Life Cycle through Ex Situ Culture

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Species and Colony Collection
2.2. Ex Situ Aquaculture Systems
2.3. Artemia Preparation and Coral Feeding
2.4. Reproductive Output and Timing
2.5. Diel Timing of Reproduction
2.6. Preliminary Assessment of Growth and Reproduction in an F1 Generation
2.7. Statistical Analyses
3. Results
3.1. Aquaculture System Temperature and Water Chemistry
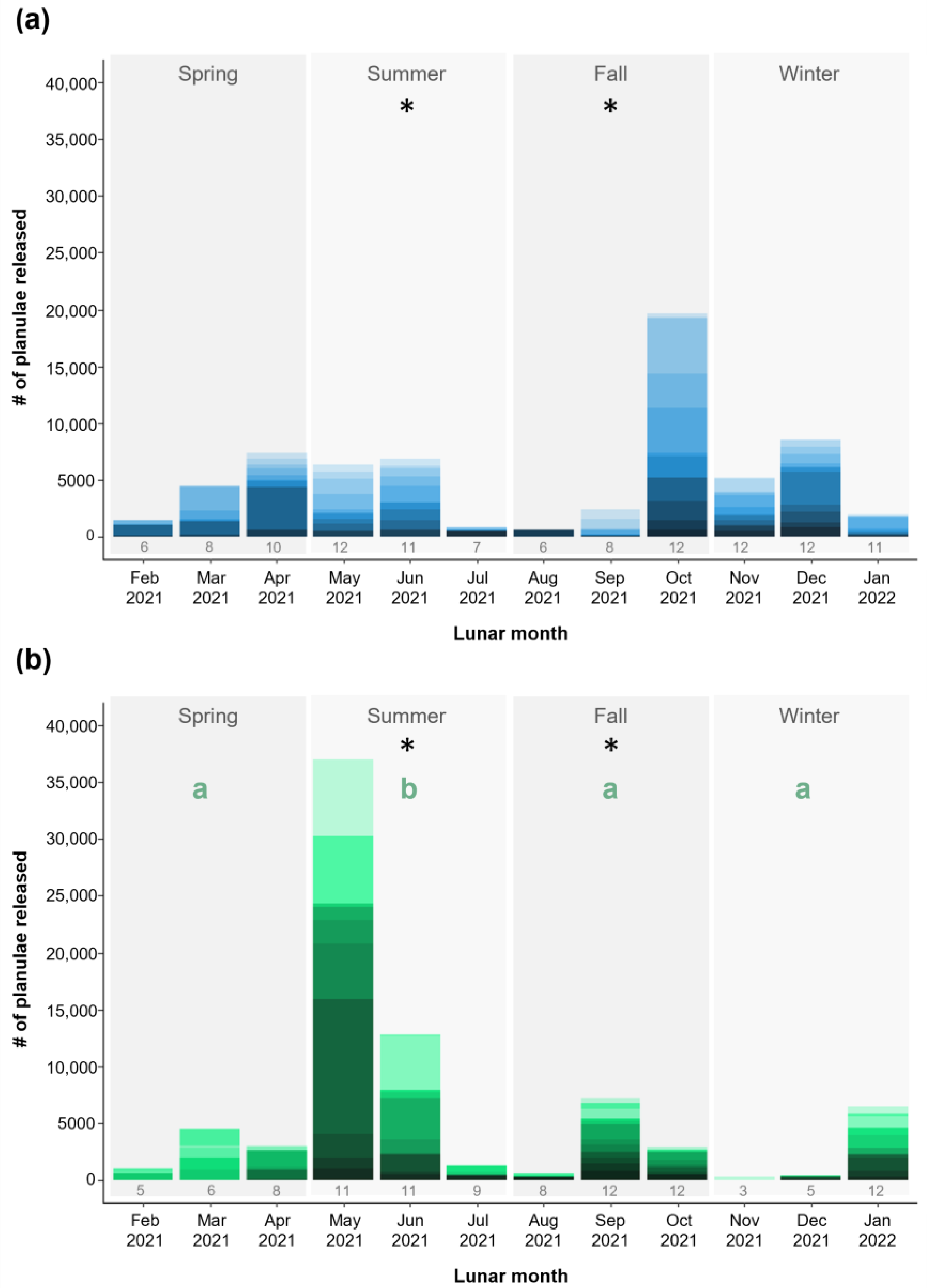
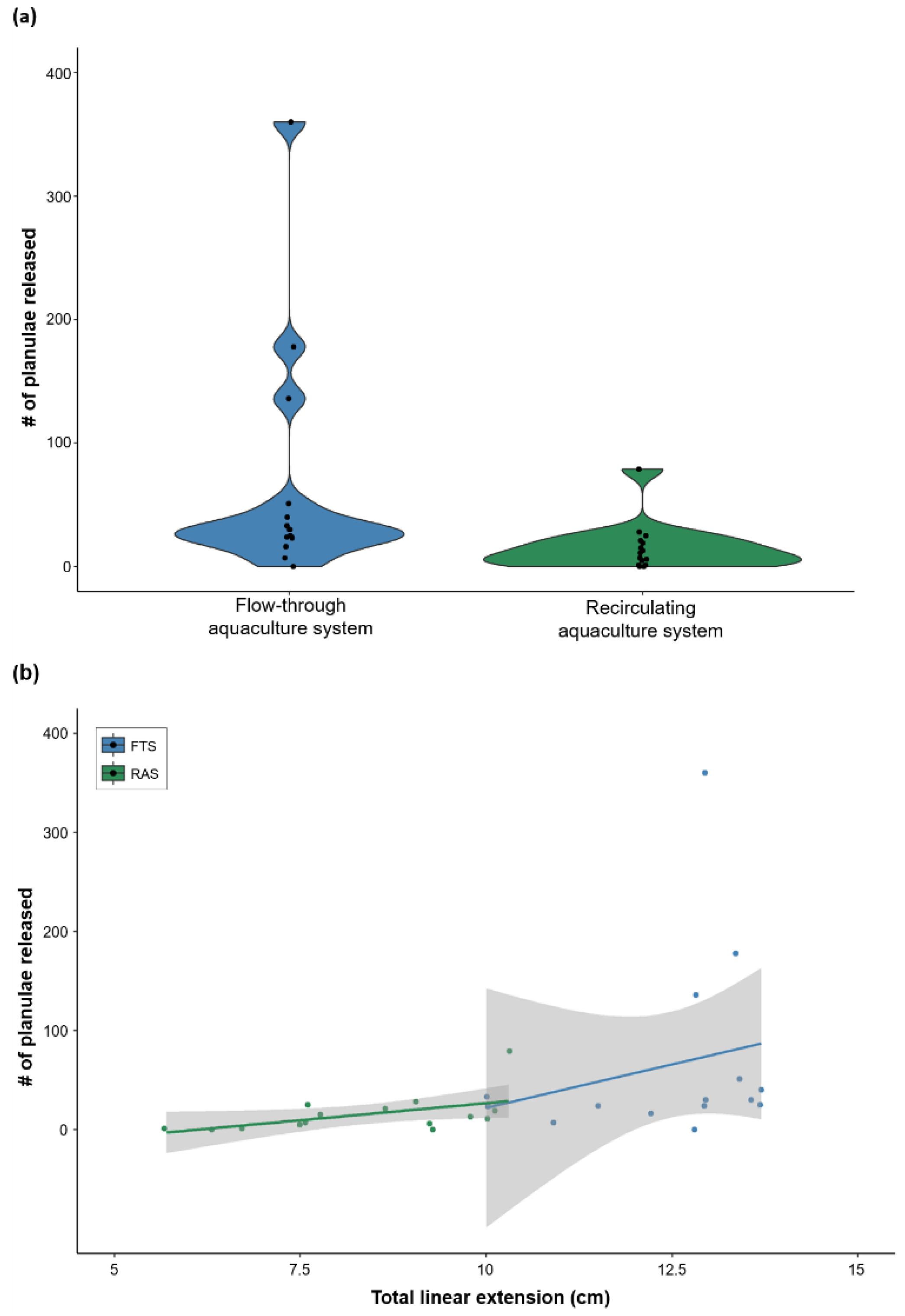
3.2. Coral Colony Reproductive Output
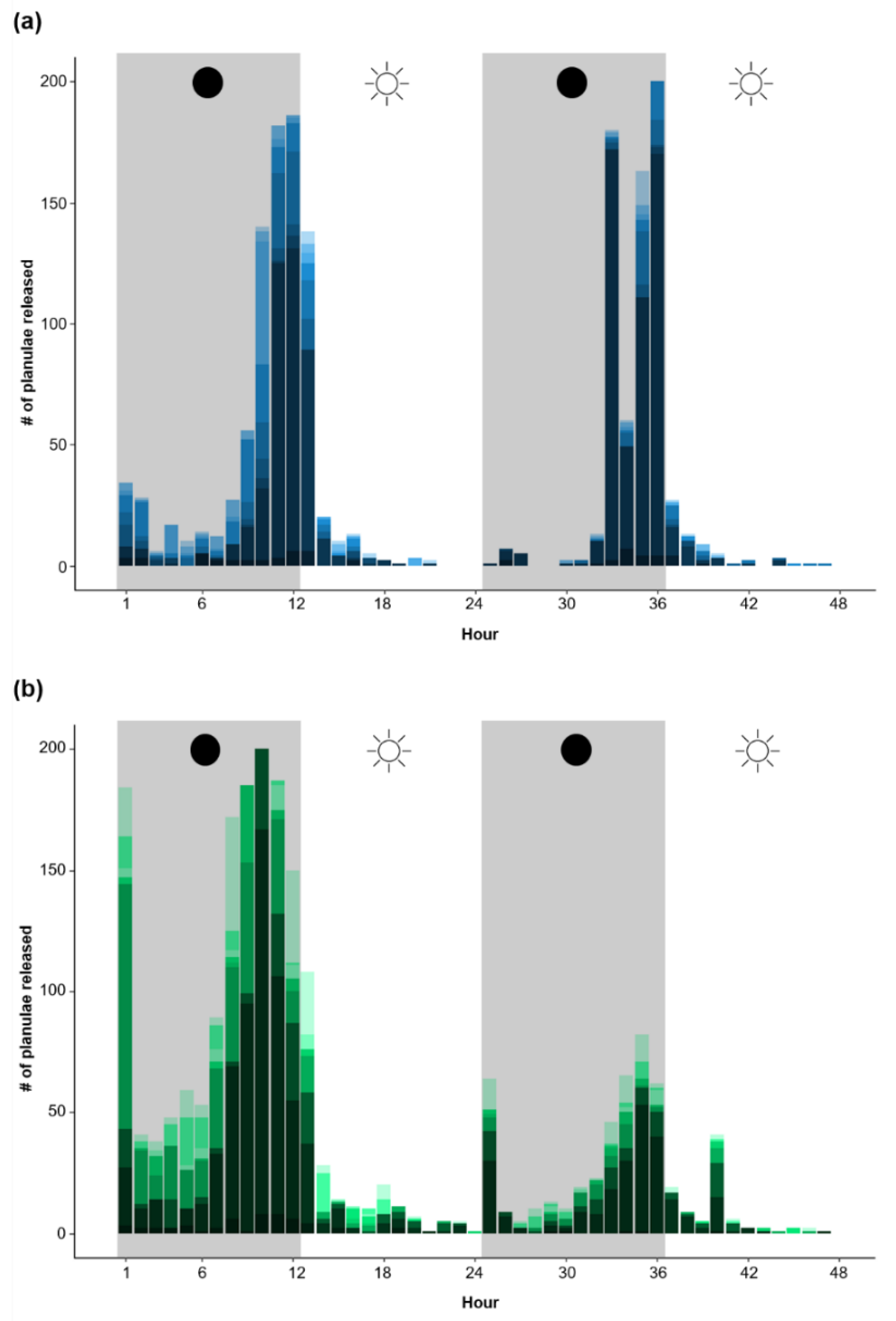
3.3. Coral Colony Reproductive Timing
3.4. Growth and Reproduction of F1 Generation Corals
4. Discussion
4.1. Potential Reasons for Successful Long-Term Ex Situ Reproduction
4.2. Comparing Aquaculture Systems: Reproductive Output
4.3. Comparing Aquaculture Systems: Reproductive Timing
4.4. Multi-Generational Coral Culture
4.5. Potential Application of Ex Situ Culture to Support Climate Change Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hoegh-Guldberg, O.; Kennedy, E.V.; Beyer, H.L.; McClennen, C.; Possingham, H.P. Securing a long-term future for coral reefs. Trends Ecol. Evol. 2018, 33, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.C.; Kleypas, J.; van de Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef]
- Donovan, M.K.; Burkepile, D.E.; Kratochwill, C.; Shlesinger, T.; Sully, S.; Oliver, T.A.; Hodgson, G.; Freiwald, J.; van Woesik, R. Local conditions magnify coral loss after marine heatwaves. Science 2021, 372, 977–980. [Google Scholar] [CrossRef] [PubMed]
- Abelson, A.; Nelson, P.A.; Edgar, G.J.; Shashar, N.; Reed, D.C.; Belmaker, J.; Krause, G.; Beck, M.W.; Brokovich, E.; France, R.; et al. Expanding marine protected areas to include degraded coral reefs. Conserv. Biol. 2016, 30, 1182–1191. [Google Scholar] [CrossRef]
- Camp, E.F. Contingency planning for coral reefs in the Anthropocene; The Potential of Reef Safe Havens. Emerg. Top. Life Sci. 2022, 6, 107–124. [Google Scholar] [CrossRef]
- Voolstra, C.R.; Suggett, D.J.; Peixoto, R.S.; Parkinson, J.E.; Quigley, K.M.; Silveira, C.B.; Sweet, M.; Muller, E.M.; Barshis, D.J.; Bourne, D.G.; et al. Extending the natural adaptive capacity of coral holobionts. Nat. Rev. Earth Environ. 2021, 2, 747–762. [Google Scholar] [CrossRef]
- Putnam, H.M. Avenues of reef-building coral acclimatization in response to rapid environmental change. J. Exp. Biol 2021, 224, jeb239319. [Google Scholar] [CrossRef] [PubMed]
- Hagedorn, M.; van Oppen, M.J.H.; Carter, V.; Henley, M.; Abrego, D.; Puill-Stephan, E.; Negri, A.; Heyward, A.; MacFarlane, D.; Spindler, R. First frozen repository for the Great Barrier Reef coral created. Cryobiology 2012, 65, 157–158. [Google Scholar] [CrossRef] [PubMed]
- Toh, E.-C.; Liu, K.-L.; Tsai, S.; Lin, C. Cryopreservation and cryobanking of cells from 100 coral species. Cells 2022, 11, 2668. [Google Scholar] [CrossRef] [PubMed]
- Shaver, E.C.; McLeod, E.; Hein, M.Y.; Palumbi, S.R.; Quigley, K.; Vardi, T.; Mumby, P.J.; Smith, D.; Montoya-Maya, P.; Muller, E.M.; et al. A roadmap to integrating resilience into the practice of coral reef restoration. Glob. Change Biol. 2022, 28, 4751–4764. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, N.; Corcoran, E.; Felis, T.; Ferse, S.; de Goeij, J.; Grottoli, A.; Harding, S.; Kleypas, J.; Mayfield, A.; Miller, M.; et al. Rebuilding Coral Reefs: A Decadal Grand Challenge; International Coral Reef Society and Future Earth Coasts: Bremen, Germany, 2021. [Google Scholar] [CrossRef]
- Froehlich, H.E.; Gentry, R.R.; Halpern, B.S. Conservation aquaculture: Shifting the narrative and paradigm of aquaculture’s role in resource management. Biol. Conserv. 2017, 215, 162–168. [Google Scholar] [CrossRef]
- Zoccola, D.; Ounais, N.; Barthelemy, D.; Calcagno, R.; Gaill, F.; Henard, S.; Hoegh-Guldberg, O.; Janse, M.; Jaubert, J.; Putnam, H.; et al. The World Coral Conservatory (WCC): A Noah’s ark for corals to support survival of reef ecosystems. PLoS Biol. 2020, 18, e3000823. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, C.; Høj, L.; Bourne, D.G. Probiotics for coral aquaculture: Challenges and considerations. Curr. Opin. Biotech. 2022, 73, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Henley, E.M.; Bouwmeester, J.; Jury, C.P.; Toonen, R.J.; Quinn, M.; Lager, C.V.A.; Hagedorn, M. Growth and survival among Hawaiian corals outplanted from tanks to an ocean nursery are driven by individual genotype and species differences rather than preconditioning to thermal stress. PeerJ 2022, 10, e13112. [Google Scholar] [CrossRef]
- Gibbs, M.T. Technology requirements, and social impacts of technology for at-scale coral reef restoration. Technol. Soc. 2021, 66, 101622. [Google Scholar] [CrossRef]
- Osinga, R.; Schutter, M.; Wijgerde, T.; Rinkevich, B.; Shafir, S.; Shpigel, M.; Luna, G.M.; Danovaro, R.; Bongiorni, L.; Deutsch, A.; et al. The CORALZOO Project: A synopsis of four years of public aquarium science. J. Mar. Biol. Assoc. 2012, 92, 753–768. [Google Scholar] [CrossRef]
- Barton, J.A.; Willis, B.L.; Hutson, K.S. Coral Propagation: A review of techniques for ornamental trade and reef restoration. Rev. Aquac. 2017, 9, 238–256. [Google Scholar] [CrossRef]
- Merck, D.E.; Petrik, C.G.; Manfroy, A.A.; Muller, E.M. Optimizing seawater temperature conditions to increase the productivity of ex situ coral nurseries. PeerJ 2022, 10, e13017. [Google Scholar] [CrossRef]
- Craggs, J.; Guest, J.R.; Davis, M.; Simmons, J.; Dashti, E.; Sweet, M. Inducing broadcast coral spawning ex situ: Closed system mesocosm design and husbandry protocol. Ecol. Evol. 2017, 7, 11066–11078. [Google Scholar] [CrossRef]
- Rocha, R.J.M.; Bontas, B.; Cartaxana, P.; Leal, M.C.; Ferreira, J.M.; Rosa, R.; Serôdio, J.; Calado, R. Development of a standardized modular system for experimental coral culture. J. World Aquac. Soc. 2015, 46, 235–251. [Google Scholar] [CrossRef]
- Craggs, J.; Guest, J.; Bulling, M.; Sweet, M. Ex situ co culturing of the sea urchin, Mespilia globulus and the coral Acropora millepora enhances early post-settlement survivorship. Sci. Rep. 2019, 9, 12984. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-L.; Mayfield, A.B.; Fan, T.-Y. Effects of feeding on the physiological performance of the stony coral Pocillopora acuta. Sci. Rep. 2020, 10, 19988. [Google Scholar] [CrossRef] [PubMed]
- Yuen, Y.S.; Yamazaki, S.S.; Nakamura, T.; Tokuda, G.; Yamasaki, H. Effects of live rock on the reef-building coral Acropora digitifera cultured with high levels of nitrogenous compounds. Aquac. Eng. 2009, 41, 35–43. [Google Scholar] [CrossRef]
- Leal, M.C.; Ferrier-Pagès, C.; Petersen, D.; Osinga, R. Coral aquaculture: Applying scientific knowledge to ex situ production. Rev. Aquac. 2016, 8, 136–153. [Google Scholar] [CrossRef]
- Osinga, R.; Schutter, M.; Griffioen, B.; Wijffels, R.H.; Verreth, J.A.J.; Shafir, S.; Henard, S.; Taruffi, M.; Gili, C.; Lavorano, S. The biology and economics of coral growth. Mar. Biotechnol. 2011, 13, 658–671. [Google Scholar] [CrossRef]
- Crowder, C.M.; Meyer, E.; Fan, T.-Y.; Weis, V.M. Impacts of temperature and lunar day on gene expression profiles during a monthly reproductive cycle in the brooding coral Pocillopora damicornis. Mol. Ecol. 2017, 26, 3913–3925. [Google Scholar] [CrossRef]
- Lin, C.-H.; Takahashi, S.; Mulla, A.J.; Nozawa, Y. Moonrise timing is key for synchronized spawning in soral Dipsastraea speciosa. Proc. Natl. Acad. Sci. USA 2021, 118, e2101985118. [Google Scholar] [CrossRef]
- van Woesik, R. Calm before the spawn: Global coral spawning patterns are explained by regional wind fields. Proc. R. Soc. B 2010, 277, 715–722. [Google Scholar] [CrossRef]
- O’Neil, K.L.; Serafin, R.M.; Patterson, J.T.; Craggs, J.R.K. Repeated ex situ spawning in two highly disease susceptible corals in the family Meandrinidae. Front. Mar. Sci. 2021, 8, 669976. [Google Scholar] [CrossRef]
- Craggs, J.; Guest, J.; Davis, M.; Sweet, M. Completing the life cycle of a broadcast spawning coral in a closed mesocosm. Invertebr. Reprod. Dev. 2020, 64, 244–247. [Google Scholar] [CrossRef]
- Jokiel, P.L.; Ito, R.Y.; Liu, P.M. Night irradiance and synchronization of lunar release of planula larvae in the reef coral Pocillopora damicornis. Mar. Biol. 1985, 88, 167–174. [Google Scholar] [CrossRef]
- Cantin, N.; Stephenson, S.; Drury, C.; Majerova, E.; Roper, C.; van Oppen, M.J.H. Pre-Conditioning Three Generations of Pocillopora acuta to Explore Mechanisms Underpinning Coral Acclimation to Climate Change Stress; International Coral Reef Society: Bremen, Germany, 2022; p. 1018. [Google Scholar]
- Fan, T.; Hsieh, Y.; Lin, K.; Kuo, F.; Soong, K.; McRae, C.; Edmunds, P.; Fang, L. Plasticity in lunar timing of larval release of two brooding Pocilloporid corals in an internal tide-induced upwelling reef. Mar. Ecol. Prog. Ser. 2017, 569, 117–127. [Google Scholar] [CrossRef]
- Crowder, C.M.; Liang, W.-L.; Weis, V.M.; Fan, T.-Y. Elevated temperature alters the lunar timing of planulation in the brooding coral Pocillopora damicornis. PLoS ONE 2014, 9, e107906. [Google Scholar] [CrossRef] [PubMed]
- Nietzer, S.; Moeller, M.; Kitamura, M.; Schupp, P.J. Coral larvae every day: Leptastrea purpurea, a brooding species that could accelerate coral research. Front. Mar. Sci. 2018, 5, 466. [Google Scholar] [CrossRef]
- McRae, C.J.; Huang, W.-B.; Fan, T.-Y.; Côté, I.M. Effects of thermal conditioning on the performance of Pocillopora acuta adult coral colonies and their offspring. Coral Reefs 2021, 40, 1491–1503. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.F.; Ravago-Gotanco, R. Rarity of the “common” coral Pocillopora damicornis in the western Philippine Archipelago. Coral Reefs 2018, 37, 1209–1216. [Google Scholar] [CrossRef]
- Johnston, E.C.; Forsman, Z.H.; Toonen, R.J. A Simple molecular technique for distinguishing species reveals frequent misidentification of Hawaiian corals in the Genus Pocillopora. PeerJ 2018, 6, e4355. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.A.; Moya, A.; Cantin, N.E.; van Oppen, M.J.H.; Torda, G. Observations of simultaneous sperm release and larval planulation suggest reproductive assurance in the coral Pocillopora acuta. Front. Mar. Sci. 2019, 6, 362. [Google Scholar] [CrossRef]
- Putnam, H.M.; Ritson-Williams, R.; Cruz, J.A.; Davidson, J.M.; Gates, R. Environmentally-induced parental or developmental conditioning influences coral offspring ecological performance. Sci. Rep. 2020, 10, 13664. [Google Scholar] [CrossRef]
- Hsu, P.-C.; Lee, H.-J.; Zheng, Q.; Lai, J.-W.; Su, F.-C.; Ho, C.-R. Tide-induced periodic sea surface temperature drops in the coral reef area of Nanwan Bay, southern Taiwan. J. Geophys. Res. Ocean 2020, 125, e2019JC015226. [Google Scholar] [CrossRef]
- Keshavmurthy, S.; Kuo, C.-Y.; Huang, Y.-Y.; Carballo-Bolaños, R.; Meng, P.-J.; Wang, J.-T.; Chen, C.A. Coral reef resilience in Taiwan: Lessons from long-term ecological research on the coral reefs of Kenting National Park (Taiwan). JMSE 2019, 7, 388. [Google Scholar] [CrossRef]
- Aquilino, F.; Paradiso, A.; Trani, R.; Longo, C.; Pierri, C.; Corriero, G.; de Pinto, M.C. Chaetomorpha linum in the bioremediation of aquaculture wastewater: Optimization of nutrient removal efficiency at the laboratory scale. Aquaculture 2020, 523, 735133. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022.
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear mixed-effects models using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.; Christensen, R. LmerTest Package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Length, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means, R Package Version 1.7.4-1. 2022.
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Kleiber, C.; Zeileis, A. Applied Econometrics with R; Springer-Verlag: New York, NY, USA, 2008; ISBN 978-0-387-77316-2. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. GlmmTMB Balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models, R package Version 0.4.5. 2022.
- Huffmyer, A.S.; Johnson, C.J.; Epps, A.M.; Lemus, J.D.; Gates, R.D. Feeding and thermal conditioning enhance coral temperature tolerance in juvenile Pocillopora acuta. R. Soc. Open Sci. 2021, 8, 210644. [Google Scholar] [CrossRef]
- Conti-Jerpe, I.E.; Thompson, P.D.; Wong, C.W.M.; Oliveira, N.L.; Duprey, N.N.; Moynihan, M.A.; Baker, D.M. Trophic strategy and bleaching resistance in reef-building corals. Sci. Adv. 2020, 6, eaaz5443. [Google Scholar] [CrossRef] [PubMed]
- Bellworthy, J.; Spangenberg, J.E.; Fine, M. Feeding increases the number of offspring but decreases parental investment of Red Sea coral Stylophora pistillata. Ecol. Evol. 2019, 9, 12245–12258. [Google Scholar] [CrossRef] [PubMed]
- Séré, M.G.; Massé, L.M.; Perissinotto, R.; Schleyer, M.H. Influence of heterotrophic feeding on the sexual reproduction of Pocillopora verrucosa in Aquaria. J. Exp. Mar. Biol. Ecol. 2010, 395, 63–71. [Google Scholar] [CrossRef]
- Conlan, J.A.; Bay, L.K.; Severati, A.; Humphrey, C.; Francis, D.S. Comparing the capacity of five different dietary treatments to optimise growth and nutritional composition in two scleractinian corals. PLoS ONE 2018, 13, e0207956. [Google Scholar] [CrossRef]
- Ward, S. Two patterns of energy allocation for growth, reproduction and lipid storage in the scleractinian coral Pocillopora damicornis. Coral Reefs 1995, 14, 87–90. [Google Scholar] [CrossRef]
- Leuzinger, S.; Willis, B.L.; Anthony, K.R.N. Energy allocation in a reef coral under varying resource availability. Mar. Biol. 2012, 159, 177–186. [Google Scholar] [CrossRef]
- Combosch, D.J.; Vollmer, S.V. Mixed asexual and sexual reproduction in the Indo-Pacific reef coral Pocillopora damicornis. Ecol. Evol. 2013, 3, 3379–3387. [Google Scholar] [CrossRef]
- Yeoh, S.-R.; Dai, C.-F. The production of sexual and asexual larvae within single broods of the scleractinian coral, Pocillopora damicornis. Mar. Biol. 2010, 157, 351–359. [Google Scholar] [CrossRef]
- Schmidt-Roach, S.; Miller, K.J.; Woolsey, E.; Gerlach, G.; Baird, A.H. Broadcast spawning by Pocillopora species on the Great Barrier Reef. PLoS ONE 2012, 7, e50847. [Google Scholar] [CrossRef]
- Chang, T.-C.; Mayfield, A.B.; Fan, T.-Y. Culture systems influence the physiological performance of the soft coral Sarcophyton glaucum. Sci. Rep. 2020, 10, 20200. [Google Scholar] [CrossRef]
- Canizales-Flores, H.M.; Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Cupul-Magaña, A.L. A long-term symbiotic relationship: Recruitment and fidelity of the crab Trapezia on its coral host Pocillopora. Diversity 2021, 13, 450. [Google Scholar] [CrossRef]
- Holling, C.S.; Walters, C. Adaptive Environmental Assessment and Management; John Wiley and Sons: Toronto, Canada, 1978. [Google Scholar]
- Gerber, N.; Kokko, H. Abandoning the ship using sex, dispersal or dormancy: Multiple escape routes from challenging conditions. Philos. Trans. R. Soc. B 2018, 373, 20170424. [Google Scholar] [CrossRef]
- Griffiths, J.G.; Bonser, S.P. Is sex advantageous in adverse environments? A test of the abandon-ship hypothesis. Am. Nat. 2013, 182, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Permata, W.; Kinzie, R.; Hidaka, M. Histological studies on the origin of planulae of the coral Pocillopora damicornis. Mar. Ecol. Prog. Ser. 2000, 200, 191–200. [Google Scholar] [CrossRef]
- Nalley, E.M.; Tuttle, L.J.; Conklin, E.E.; Barkman, A.L.; Wulstein, D.M.; Schmidbauer, M.C.; Donahue, M.J. A systematic review and meta-analysis of the direct effects of nutrients on corals. Sci. Total Environ. 2023, 856, 159093. [Google Scholar] [CrossRef]
- Loya, Y.; Lubinevsky, H.; Rosenfeld, M.; Kramarsky-Winter, E. Nutrient enrichment caused by in situ fish farms at Eilat, Red Sea is detrimental to coral reproduction. Mar. Pollut. Bull. 2004, 49, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Isomura, N.; Nishihira, M. Size variation of planulae and its effect on the lifetime of planulae in three Pocilloporid corals. Coral Reefs 2001, 20, 309–315. [Google Scholar] [CrossRef]
- de Putron, S.J.; Lawson, J.M.; White, K.Q.L.; Costa, M.T.; Geronimus, M.V.B.; MacCarthy, A. Variation in larval properties of the Atlantic brooding coral Porites astreoides between different reef sites in Bermuda. Coral Reefs 2017, 36, 383–393. [Google Scholar] [CrossRef]
- Cumbo, V.R.; Fan, T.-Y.; Edmunds, P.J. Physiological development of brooded larvae from two Pocilloporid corals in Taiwan. Mar. Biol. 2012, 159, 2853–2866. [Google Scholar] [CrossRef]
- Linden, B.; Huisman, J.; Rinkevich, B. Circatrigintan instead of lunar periodicity of larval release in a brooding coral species. Sci. Rep. 2018, 8, 5668. [Google Scholar] [CrossRef]
- Fan, T.; Lin, K.; Kuo, F.; Soong, K.; Liu, L.; Fang, L. Diel patterns of larval release by five brooding scleractinian corals. Mar. Ecol. Prog. Ser. 2006, 321, 133–142. [Google Scholar] [CrossRef]
- Goodbody-Gringley, G. Diel Planulation by the brooding coral Favia fragum (Esper, 1797). J. Exp. Mar. Biol. Ecol. 2010, 389, 70–74. [Google Scholar] [CrossRef]
- Torda, G.; Donelson, J.M.; Aranda, M.; Barshis, D.J.; Bay, L.; Berumen, M.L.; Bourne, D.G.; Cantin, N.; Foret, S.; Matz, M.; et al. Rapid adaptive responses to climate change in corals. Nat. Clim. Change 2017, 7, 627–636. [Google Scholar] [CrossRef]
- Byrne, M.; Foo, S.A.; Ross, P.M.; Putnam, H.M. Limitations of cross- and multigenerational plasticity for marine invertebrates faced with global climate change. Glob. Change Biol. 2020, 26, 80–102. [Google Scholar] [CrossRef]
- Humanes, A.; Beauchamp, E.A.; Bythell, J.C.; Carl, M.K.; Craggs, J.R.; Edwards, A.J.; Golbuu, Y.; Lachs, L.; Martinez, H.M.; Palmowski, P.; et al. An experimental framework for selectively breeding corals for assisted evolution. Front. Mar. Sci. 2021, 8, 669995. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Sweet, M.; Villela, H.D.M.; Cardoso, P.; Thomas, T.; Voolstra, C.R.; Høj, L.; Bourne, D.G. Coral probiotics: Premise, promise, prospects. Annu. Rev. Anim. Biosci. 2021, 9, 265–288. [Google Scholar] [CrossRef] [PubMed]
- Kleypas, J.; Allemand, D.; Anthony, K.; Baker, A.C.; Beck, M.W.; Hale, L.Z.; Hilmi, N.; Hoegh-Guldberg, O.; Hughes, T.; Kaufman, L.; et al. Designing a blueprint for coral reef survival. Biol. Conserv. 2021, 257, 109107. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, K.-W.; McRae, C.J.; Zhang, X.-C.; Ye, Z.-M.; Qiu, Y.-T.; Jiang, M.-Q.; Cheng, T.-H.; Chen, G.K.; Fan, T.-Y. Consistent Monthly Reproduction and Completion of a Brooding Coral Life Cycle through Ex Situ Culture. Diversity 2023, 15, 218. https://doi.org/10.3390/d15020218
Lam K-W, McRae CJ, Zhang X-C, Ye Z-M, Qiu Y-T, Jiang M-Q, Cheng T-H, Chen GK, Fan T-Y. Consistent Monthly Reproduction and Completion of a Brooding Coral Life Cycle through Ex Situ Culture. Diversity. 2023; 15(2):218. https://doi.org/10.3390/d15020218
Chicago/Turabian StyleLam, Kwok-Wai, Crystal J. McRae, Xuan-Ci Zhang, Zong-Min Ye, Yu-Ting Qiu, Ming-Qi Jiang, Ting-Hui Cheng, Guanyan Keelung Chen, and Tung-Yung Fan. 2023. "Consistent Monthly Reproduction and Completion of a Brooding Coral Life Cycle through Ex Situ Culture" Diversity 15, no. 2: 218. https://doi.org/10.3390/d15020218
APA StyleLam, K.-W., McRae, C. J., Zhang, X.-C., Ye, Z.-M., Qiu, Y.-T., Jiang, M.-Q., Cheng, T.-H., Chen, G. K., & Fan, T.-Y. (2023). Consistent Monthly Reproduction and Completion of a Brooding Coral Life Cycle through Ex Situ Culture. Diversity, 15(2), 218. https://doi.org/10.3390/d15020218