Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue




{kind=link}
{kind=link}
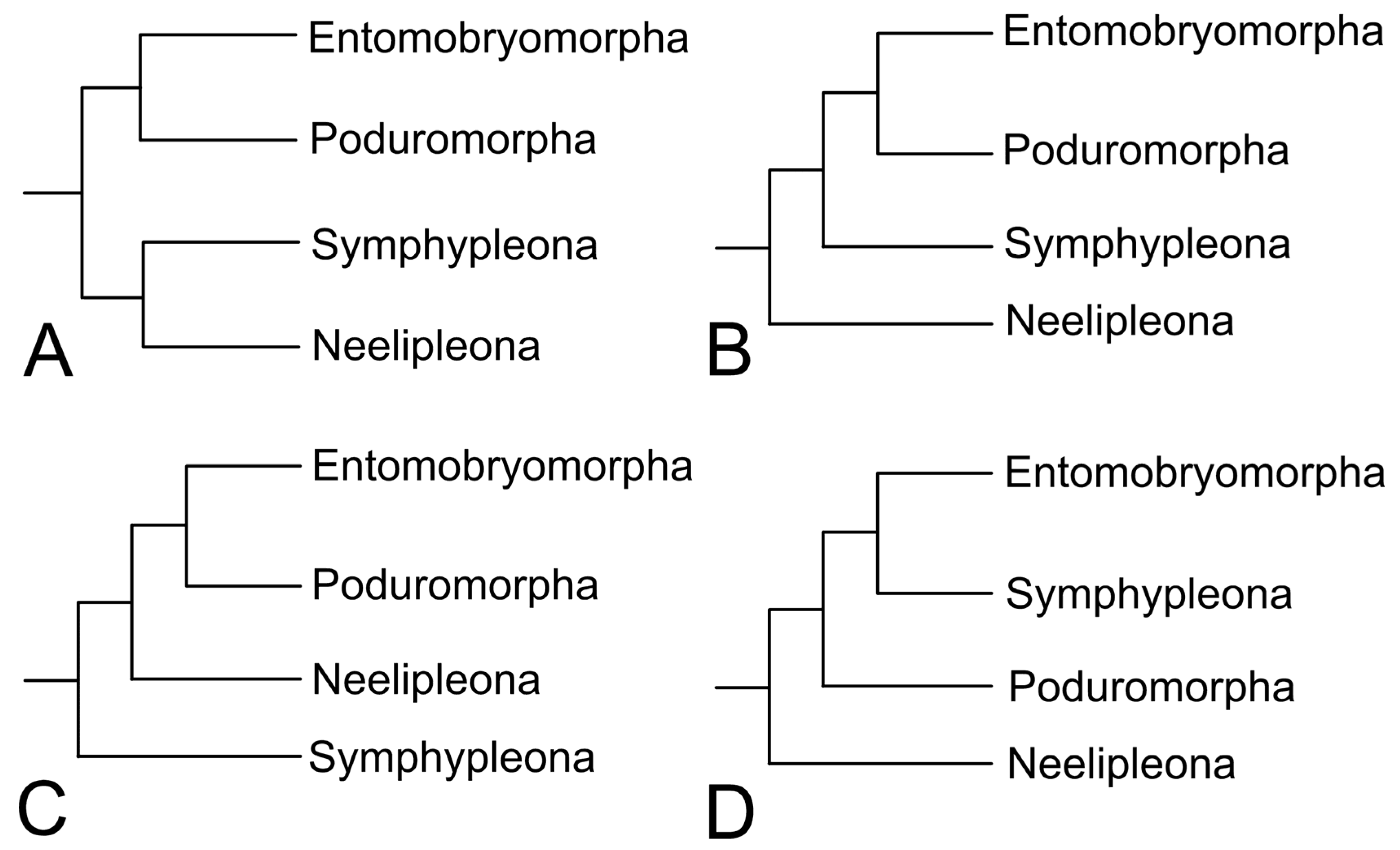{kind=link}
- Diversity’s Special Issue: Systematics, Ecology and Taxonomy of Collembola
- Knowing More about Springtails
Acknowledgments
Conflicts of Interest
References
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 24 January 2023).
- Bretfeld, G.; Gauer, U. Diagnostic description of the males of new Sphaeridia species (Insecta, Collembola) from South America. Andrias 1994, 13, 113–136. [Google Scholar]
- Bretfeld, G. Synopses on Palaeartic Collembola, Volume 2. Symphypleona. Abh. Ber. Naturkundemus. Gorlitz 1999, 71, 1–318. [Google Scholar]
- Grimaldi, D.; Engel, M.S. Evolution of the Insects, 1st ed.; Cambridge University Press: New York, NY, USA, 2005; pp. 1–755. [Google Scholar]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Beutel, R.G.; Yavorskaya, M.I.; Mashimo, Y.; Fukui, M.; Meusemann, K. The Phylogeny of Hexapoda (Arthropoda) and the Evolution of Megadiversity. Proc. Arthropod. Embryol. Soc. Jpn. 2017, 51, 1–15. [Google Scholar]
- Hopkin, S.P. Biology of the Springtails (Insecta: Collembola), 1st ed.; Oxford University Press: New York, NY, USA, 1997; pp. 1–330. [Google Scholar]
- Bellini, B.C. Colêmbolos: Uma riqueza microscópica no Semiárido. In Conhecendo os Artrópodes do Semiárido, 1st ed.; Bravo, F., Calor, A.R., Eds.; Métis Produção Editorial: São Paulo, Brazil, 2016; Volume 1, pp. 43–55. [Google Scholar]
- Bellini, B.C.; Zhang, F.; Souza, P.G.C.; Santos-Costa, R.C.; Medeiros, G.S.; Godeiro, N.N. The Evolution of Collembola Higher Taxa (Arthropoda, Hexapoda) Based on Mitogenome Data. Diversity 2023, 15, 7. [Google Scholar] [CrossRef]
- Davis, R.B.; Baldauf, S.L.; Mayhew, P.J. Many hexapod groups originated earlier and withstood extinction events better than previously realized: Inferences from supertrees. Proc. R. Soc. B Biol. Sci. 2010, 277, 1597–1606. [Google Scholar] [CrossRef]
- Greenslade, P.; Whalley, P.E.S. The systematic position of Rhyniella praecursor Hirst & Maulik (Collembola). The earliest known hexapod. In Proceedings of the Second International Seminar on Apterygota, Siena, Italy, 4–6 September 1986; Volume 1, pp. 319–323. [Google Scholar]
- Soto-Adames, F.N.; Barra, J.A.; Christiansen, K.; Jordana, R. Suprageneric classification of Collembola Entomobryomorpha. Ann. Entomol. Soc. Am. 2008, 101, 501–513. [Google Scholar] [CrossRef]
- Zhang, F.; Deharveng, L. Systematic revision of Entomobryidae (Collembola) by integrating molecular and new morphological evidence. Zool. Scr. 2015, 44, 298–311. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, D.; Yu, D.; Wang, B. Molecular phylogeny supports S-chaetae as a key character better than jumping organs and body scales in classification of Entomobryoidea (Collembola). Sci. Rep. 2015, 5, 12471. [Google Scholar] [CrossRef]
- Zhang, F.; Bellini, B.C.; Soto-Adames, F.N. New insights into the systematics of Entomobryoidea (Collembola: Entomobryomorpha): First instar chaetotaxy, homology and classification. Zool. Syst. 2019, 44, 249–278. [Google Scholar] [CrossRef]
- Zhang, F.; Cipola, N.G.; Pan, Z.-X.; Ding, Y. New insight into the systematics of Heteromurini (Collembola: Entomobryidae: Heteromurinae) with special reference to Alloscopus and Sinodicranocentrus gen. n. Arth. Syst. Phylogeny 2020, 78, 1–16. [Google Scholar] [CrossRef]
- Godeiro, N.N.; Pacheco, G.; Liu, S.; Cipola, N.G.; Berbel-Filho, W.M.; Zhang, F.; Gilbert, M.T.; Bellini, B.C. Phylogeny of Neotropical Seirinae (Collembola, Entomobryidae) based on mitochondrial genomes. Zool. Scr. 2020, 49, 329–339. [Google Scholar] [CrossRef]
- Godeiro, N.N.; Ding, Y.; Cipola, N.G.; Jantarit, S.; Bellini, B.C.; Zhang, F. Phylogenomics and systematics of Entomobryoidea (Collembola): Marker design, phylogeny and classification. Cladistics 2022, 1–15. [Google Scholar] [CrossRef]
- Yu, D.; Ding, Y.; Tihelka, E.; Cai, C.; Hu, F.; Liu, M.; Zhang, F. Phylogenomics of Elongate-Bodied Springtails Reveals Independent Transitions from Aboveground to Belowground Habitats in Deep Time. Syst. Biol. 2022, 71, 1023–1031. [Google Scholar] [CrossRef]
- Gao, Y.; Bu, Y.; Luan, Y.-X. Phylogenetic Relationships of Basal Hexapods Reconstructed from Nearly Complete 18S and 28S rRNA Gene Sequences. Zool. Sci. 2008, 25, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Cruaud, C.; D’Haese, C.A. Unexpected diversity in Neelipleona revealed by molecular phylogeny approach (Hexapoda, Collembola). Soil Org. 2011, 83, 383–398. [Google Scholar]
- Schneider, C.; D’Haese, C.A. Morphological and molecular insights on Megalothorax: The largest Neelipleona genus revisited (Collembola). Invertebr. Syst. 2013, 27, 317–364. [Google Scholar] [CrossRef]
- Leo, C.; Carapelli, A.; Cicconardi, F.; Frati, F.; Nardi, F. Mitochondrial genome diversity in Collembola: Phylogeny, dating and gene order. Diversity 2019, 11, 169. [Google Scholar] [CrossRef]
- Lima, E.C.A.; Lopes, B.C.H.; Oliveira-Neto, M.A.; Mendonça, M.C.; Zeppelini, D. Synthesis of Brazilian Entomobryomorpha (Collembola: Hexapoda) with Special Emphasis on the Equatorial Oceanic Islands and Redescription of the First Species of Collembola Recorded in Brazil. Diversity 2022, 14, 553. [Google Scholar] [CrossRef]
- Medeiros, G.S.; Nunes, R.C.; Zhang, F.; Godeiro, N.N.; Bellini, B.C. A New Genus of Sminthurididae (Collembola, Symphypleona) from Brazil, with Notes on the Systematics of the Family. Diversity 2022, 14, 960. [Google Scholar] [CrossRef]
- Greenslade, P.; Weiner, W.M. Taxonomic papers on Collembola in Zootaxa, 2001 to 2020. Zootaxa 2021, 4979, 215–217. [Google Scholar] [CrossRef]
- Cicconardi, F.; Fanciulli, P.P.; Emerson, B.C. Collembola, the biological species concept and the underestimation of global species richness. Mol. Ecol. 2013, 22, 5382–5396. [Google Scholar] [CrossRef] [PubMed]
- Dukes, C.D.; Janssens, F.; Recuero, E.; Caterino, M.S. Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA). Diversity 2022, 14, 847. [Google Scholar] [CrossRef]
- Christiansen, K. Bionomics of Collembola. Annu. Rev. Entomol. 1964, 9, 147–178. [Google Scholar] [CrossRef]
- Petersen, H.; Luxton, M. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos 1982, 39, 288–388. [Google Scholar] [CrossRef]
- Pollierer, M.M.; Scheu, S. Driving factors and temporal fluctuation of Collembola communities and reproductive mode across forest types and regions. Ecol. Evol. 2017, 7, 4390–4403. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, K. Convergence and parallelism in cave Entomobryinae. Evolution 1961, 15, 288–301. [Google Scholar] [CrossRef]
- Verhoef, H.A.; Brussaard, L. Decomposition and nitrogen mineralization in natural and agroecosystems: The contribution of soil animals. Biogeochemistry 1990, 11, 175–211. [Google Scholar] [CrossRef]
- Rusek, J. Biodiversity of Collembola and their functional role in the ecosystem. Biodivers. Conserv. 1998, 7, 1207–1219. [Google Scholar] [CrossRef]
- Potapov, A.; Bellini, B.C.; Chown, S.L.; Deharveng, L.; Janssens, F.; Kovác, L.; Kuznetsova, N.; Ponge, J.-F.; Potapov, M.; Querner, P.; et al. Towards a global synthesis of Collembola knowledge—Challenges and potential solutions. Soil Org. 2020, 92, 161–188. [Google Scholar] [CrossRef]
- Forey, E.; Coulibaly, S.F.M.; Chauvat, M. Flowering phenology of a herbaceous species (Poa annua) is regulated by soil Collembola. Soil Biol. Biochem. 2015, 90, 30–33. [Google Scholar] [CrossRef]
- Winck, B.R.; Chauvat, M.; Coulibaly, S.F.M.; Santonja, M.; Saccol de Sá, E.L.; Forey, E. Functional collembolan assemblages induce different plant responses in Lolium perenne. Plant Soil 2020, 452, 347–358. [Google Scholar] [CrossRef]
- Castaño-Meneses, G.; Palacios-Vargas, J.G.; Cutz-Pool, L.Q. Feeding habits of Collembola and their ecological niche. An. Inst. Biol. Ser. Zool. 2004, 75, 135–142. [Google Scholar]
- Souza, P.G.C.; Medeiros, G.S.; Ferreira, R.L.; Souza-Silva, M.; Bellini, B.C. A Highly Troglomorphic New Genus of Sminthuridae (Collembola, Symphypleona) from the Brazilian Semiarid Region. Insects 2022, 13, 650. [Google Scholar] [CrossRef]
- Maaß, S.; Caruso, T.; Rillig, M. Functional role of microarthropods in soil aggregation. Pedobiologia 2015, 58, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.D.D.; Bellini, B.C.; Rigotti, V.M.; Nunes, R.C.; Menezes, L.S.; Winck, B.R. Diversity Loss of Epigeic Collembola after Grassland Conversion into Eucalyptus Forestry in Brazilian Pampa Domain. Diversity 2022, 14, 490. [Google Scholar] [CrossRef]
- Culik, M.P.; Zeppelini Filho, D. Diversity and distribution of Collembola (Arthropoda: Hexapoda) of Brazil. Biodivers. Conserv. 2003, 12, 1119–1143. [Google Scholar] [CrossRef]
- Zeppelini, D.; Queiroz, G.C.; Bellini, B.C. Collembola in Catálogo Taxonômico da Fauna do Brasil. PNUD. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/470 (accessed on 24 January 2023).
- Betsch, J.M. Éléments pour une monographie des Collemboles Symplyplêones (Hexapodes, Aptérygotes). Mém. Mus. Natl. Hist. Nat. Sér. A Zool. 1980, 116, 1–227. [Google Scholar]
- Christiansen, K.; Bellinger, P. The Collembola of North America. North of Rio Grande, a Taxonomy Analysis, 2nd ed.; Grinnell College: Grinnell, IA, USA, 1998; pp. 1–1520. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Families: Neogastruridae and Brachystomellidae, 1st ed.; Polska Akademia Umiejętności, Acta monographica Musei Historiae Naturalis: Kraków, Poland, 1949; pp. 1–341. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Families: Anuridae and Pseudachorutidae, 1st ed.; Polska Akademia Umiejętności, Acta monographica Musei Historiae Naturalis: Kraków, Poland, 1949; pp. 1–122. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Bilobidae, 1st ed.; Polska Akademia Umiejętności, Acta monographica Musei Historiae Naturalis: Kraków, Poland, 1951; pp. 1–97. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Onychiuridae, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1954; pp. 1–219. [Google Scholar]
- Massoud, Z. Monographie des Neanuridae, Collemboles Poduromorphes apiéces buccales modifiées. In Biologie de l’Amerique Australe, 1st ed.; Delamare Deboutteville, C., Rapoport, E.H., Eds.; Éditions du CNRS: Paris, France, 1967; Volume 3, pp. 7–399. [Google Scholar]
- Dunger, W. Synopses on Palaeartic Collembola, Volume 1. Tullbergiinae. Abh. Ber. Naturkundemus. Gorlitz 1994, 68, 1–71. [Google Scholar]
- Cassagnau, P. Collemboles Palaeonurini primitifs d’Afrique et Madagascar. Ann. Soc. Entomol. Fr. 1996, 32, 7–33. [Google Scholar]
- Weiner, W.M. Generic revision of Onychiurinae (Onychiuridae, Collembola) with a cladistic analysis. Ann. Soc. Entomol. Fr. 1996, 32, 163–200. [Google Scholar]
- Najt, J.; Weiner, W.M. Geographic distribution of Brachystomellinae (Collembola, Neanuridae). Pan-Pacific Entomol. 1996, 72, 61–69. [Google Scholar]
- Jordana, R.; Arbea, J.I.; Simón, C.; Luciáñez, M.J. Fauna Iberica Volume 8: Collembola, Poduromorpha, 1st ed.; Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1997; pp. 1–807. [Google Scholar]
- Pomorski, R.J. Onychiurinae of Poland (Collembola: Onychiuridae), 1st ed.; Polish Taxonomical Society: Wrocław, Poland, 1998; pp. 1–210. [Google Scholar]
- Fjellberg, A. Fauna Entomologica Scandinavica—The Collembola of Fennoscandia and Denmark, Part I: Poduromorpha, 1st ed.; Brill: Leiden, The Netherlands, 1998; pp. 1–184. [Google Scholar]
- Thibaud, J.M.; Schulz, H.J.; Assalino, M.M.G. Synopses on Palaeartic Collembola, Volume 4: Hypogastruridae. Abh. Ber. Naturkundemus. Gorlitz 2004, 75, 1–287. [Google Scholar]
- Weiner, W.M.; Najt, J. Species of Brachystomella (Collembola: Brachystomellidae) from the Neotropical region. Eur. J. Entomol. 2001, 98, 387–413. [Google Scholar] [CrossRef]
- Najt, J.; Weiner, W.M.; Grandcolas, P. Phylogeny of the Brachystomellidae (Collembola)—Were the mandibles ancestrally absent and did they reappear in this family? Zool. Scr. 2005, 34, 305–312. [Google Scholar] [CrossRef]
- Paśnik, G.; Weiner, W.M. First phylogenetic analysis of the tribe Oligaphorurini (Collembola: Onychiuridae) inferred from morphological data, with implications for generic classification. Org. Divers. Evol. 2017, 17, 619–631. [Google Scholar] [CrossRef]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Isotomidae, 1st ed.; Polska Akademia Umiejętności, Acta monographica Musei Historiae Naturalis: Kraków, Poland, 1947; pp. 1–488. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Tribe: Orchesellini, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1957; pp. 1–151. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Tribe: Entomobryini, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1957; pp. 1–126. [Google Scholar]
- Potapov, M. Synopses on Palearctic Collembola, Volume 3: Isotomidae. Abh. Ber. Naturkundemus. Gorlitz 2001, 73, 1–603. [Google Scholar]
- Fjellberg, A. Fauna Entomologica Scandinavica—The Collembola of Fennoscandia and Denmark, Part II: Entomobryomorpha and Symphypleona, 1st ed.; Brill: Leiden, The Netherlands, 2007; pp. 1–264. [Google Scholar]
- Jordana, R. Synopses on Palaeartic Collembola, Volume 7/1: Capbryinae & Entomobryini. Soil Org. 2012, 84, 1–390. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Sminthuridae, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1956; pp. 1–287. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Families: Neelidae and Dicyrtomidae, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1957; pp. 1–113. [Google Scholar]
- Richards, W.R. Generic classification, evolution, and biogeography of the Sminthuridae of the world (Collembola). Mem. Ent. Soc. Can. 1968, 53, 3–54. [Google Scholar] [CrossRef]
- Bonet, F. Monografia de la familia Neelidae (Collembola). Rev. Soc. Mex. Hist. Nat. 1947, 8, 131–192. [Google Scholar]
- Schneider, C. Morphological review of the order Neelipleona (Collembola) through the redescription of the type species of Acanthoneelidus, Neelides and Neelus. Zootaxa 2017, 4308, 1–94. [Google Scholar] [CrossRef]
- Christiansen, K.; Bellinger, P. Insects of Hawaii—A Manual of the Insects of the Hawaiian Islands, including an Enumeration of the Species and Notes on Their Origin, Distribution, Hosts, Parasites, etc. Volume 15—Collembola, 1st ed.; University of Hawaii Press: Honolulu, HI, USA, 1992; pp. 1–445. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellini, B.C.; Weiner, W.M.; Winck, B.R. Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue. Diversity 2023, 15, 221. https://doi.org/10.3390/d15020221
Bellini BC, Weiner WM, Winck BR. Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue. Diversity. 2023; 15(2):221. https://doi.org/10.3390/d15020221
Chicago/Turabian StyleBellini, Bruno Cavalcante, Wanda Maria Weiner, and Bruna Raquel Winck. 2023. "Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue" Diversity 15, no. 2: 221. https://doi.org/10.3390/d15020221
APA StyleBellini, B. C., Weiner, W. M., & Winck, B. R. (2023). Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue. Diversity, 15(2), 221. https://doi.org/10.3390/d15020221